
东方蜜蜂卵黄原蛋白基因cDNA克隆及其基本生物信息学特征
2015-05-25 00:35王秀秀杨明华匡海鸥和绍禹李亚辉
中国蜂业 2015年3期
王秀秀 杨明华 李 昌 匡海鸥 和绍禹 李亚辉
(1云南农业大学动物科学技术学院,昆明650201;2云南农业大学云南省动物营养与饲料重点实验室,昆明650201;3云南农业大学食品科学技术学院,昆明650201)
东方蜜蜂卵黄原蛋白基因cDNA克隆及其基本生物信息学特征
王秀秀1*杨明华1,2*李 昌3匡海鸥3和绍禹3李亚辉3
(1云南农业大学动物科学技术学院,昆明650201;2云南农业大学云南省动物营养与饲料重点实验室,昆明650201;3云南农业大学食品科学技术学院,昆明650201)
卵黄原蛋白(vitellogenin,Vg)基因在蜜蜂的卵巢发育、胚胎营养、劳动分工、行为调控、气候适应、寿命延长等方面起着重要作用,但东方蜜蜂中该基因的基础数据却鲜有报道。本研究对东方蜜蜂Vg基因的cDNA全长序列进行了克隆、测序和基本的生物信息学分析,结果表明:东方蜜蜂Vg基因cDNA全长序列为5313bp,提交到GenBank的登录号为KP119837,该基因共编码1770个氨基酸,其cDNA和所编码的氨基酸序列与意大利蜜蜂的同源性分别为93%和89%,其保守区序列占全序列的63%。基于Vg基因对蜜蜂属内序列已知的4种蜜蜂构建的系统发育树显示:东方蜜蜂与意大利蜜蜂的亲缘关系最近,与大蜜蜂的亲缘关系稍远,与小蜜蜂的亲缘关系最远。本论文为进一步研究东方蜜蜂Vg基因的功能奠定了基础。
东方蜜蜂,卵黄原蛋白,Vg,cDNA,克隆,生物信息学
卵黄原蛋白(vitellogenin,Vg)普遍存在于卵生雌性动物的卵黄前体中,是一个专门结合和转运母体资源(如脂质、碳水化合物、金属离子和磷)供应卵母细胞的磷酸糖脂蛋白[1]。卵黄原蛋白前体在卵巢外脂肪体组织,经雌雄特异性和阶段特异性地合成,后被分泌到血淋巴,最后被卵母细胞通过受体介导的内吞作用摄取[2~4]。在脂肪体中,Vg分子被蛋白酶水解后,再经翻译后修饰以协助将碳水化合物、脂质和其他营养物质转运到卵巢。被卵母细胞吸收后,Vg以结晶形式的卵黄蛋白((vitellin,Vn或yolk protein,YP))贮存起来,作为未来胚胎的营养来源[2,5~7]。
在昆虫中,卵黄原蛋白最早在惜古比天蚕体内发现,当时被称作雌性特异性蛋白,Tefler证明其参与卵黄的形成[8]。1969年,Pan等人确定了脂肪体是该蛋白的主要合成部位,并将其命名为Vg,作为卵黄蛋白的前体[9]。之后的研究表明,在其他昆虫中,雌性个体的脂肪体是Vg生物合成的主要部位[10,11]。但这并不意味着Vg只能在脂肪体中合成,例如,环裂亚目的卵巢滤泡上皮细胞也产生Vg[12-14]。值得一提的是:一些物种的雄性动物体内也发现有Vg存在(尽管量很少),因此,Vg不再被认为具有雌性特异性[15~18]。
蜜蜂只能在脂肪体中合成Vg,其分子量为180 kDa[19]。大多数物种只在具有生殖能力的雌性中合成Vg,而在蜜蜂中,Vg在繁殖力很强的蜂王体内,雄蜂体内以及不具有生殖能力的工蜂体内均能表达[20]。Vg是由工蜂的咽下腺产生的,作为蜂王浆的一部分(是蜂王浆的氨基酸来源)被饲喂给蜂王,为蜂王和发育中的幼虫提供营养[21]。国外的研究发现,Vg在蜜蜂中还有多种调节功能:(1)Vg抑制工蜂从巢内工作向采集工作转变[22];(2)Vg影响工蜂的采集偏好,血淋巴中Vg水平低的年轻工蜂偏爱采集花蜜,而Vg水平较高的工蜂则偏爱采集花粉[23];(3)Vg可以通过清除自由基,减少氧化应激,从而延长工蜂和蜂王的寿命[24,25]。此外,Vg还与蜜蜂的气候适应,生殖竞争相关[26,27]。
但有关东方蜜蜂Vg基因研究的报道极少,本论文以分子生物学理论为基础,利用分子生物学和生物信息学技术手段,对东方蜜蜂的Vg基因进行了克隆、测序和基本的生物信息学分析,本研究为解读蜜蜂功能基因Vg,深入研究东方蜜蜂Vg基因的作用与功能奠定了良好的基础。
1 材料与实验方法
1.1 实验材料
本实验所使用的东方蜜蜂由云南农业大学东方蜜蜂研究所提供。
1.2 卵黄原蛋白基因的cDNA扩增
取东方蜜蜂的腹部,根据TRNzol-A+总RNA提取试剂盒(TIANGEN)的操作说明提取总RNA,根据NCBI中已发表的意大利蜂的mRNA序列分三段设计引物(FF:5'CATGTTGCTACTTCTAACGCTT 3';FR:5' GTTTGGGTAATGGCATCTCT 3';SF:5'ACCGAACGATAAATCTCAGG 3' ;SR: 5' CATCTTCGGCTTCTGGTC 3';TF:5' GGAAACTCTTCTGTCCTACG 3';TR:5'CTTTCGAGTTAAGCCTTGC 3'),按照TIANScript cDNA第一链合成试剂盒 (TIANGEN)合成cDNA第一链。
使用Taq Polymerase(TIANGEN)和Long Taq Polymerase(TIANGEN)分别对目的片段进行扩增。
1.3 cDNA的克隆与测序
使用普通琼脂糖DNA回收试剂盒(TIANGEN)纯化目的基因PCR产物,然后用零背景快速连接试剂盒(TIANGEN)进行连接转化,挑取重组克隆菌落进行菌落PCR,提取质粒并进行酶切鉴定,最后将阳性克隆送到英潍捷基(上海)贸易有限公司测序。
1.4 卵黄原蛋白基因的生物信息学分析
使用SeqMan对得到的测序结果进行校正和拼接得到Vg基因cDNA全长序列。然后将处理后的序列分别应用Blastn检索程序(http://www.ncbi.nlm.nih.gov)和开放阅读框(ORF)查找程序(http://www.ncbi.nlm.nih. gov/gorf)在GenBank数据库中进行核苷酸序列同源性检索分析与开放阅读框识别。同时,将所得序列用EditSeq翻译成氨基酸。最后将所得到的Vg基因cDNA全长序列及其编码的氨基酸序列通过Banklt提交程序提交到GenBank。
将东方蜜蜂Vg基因cDNA全长序列与在Gen-Bank检索到的已提交的同源性序列在MEGA中进行Alignment计算后再用Gblock 0.91软件(在线服务器:http://www.phylogeny.lirmm.fr/phylo_cgi/one_task.cgi
task_type=gblocks)进行保守区序列选择后,最后用MEGA和MrBayes件用ML(最大似然法),MP(最大简约法)和BI(贝叶斯法)进行系统发育分析并构建系统发育树。
2 结果与分析
2.1 东方蜜蜂卵黄原蛋白基因cDNA全长序列克隆
东方蜜蜂Vg基因cDNA全长序列为5313bp,提交到GenBank的登录号为KP119837,A、G、T、C的碱基数分别为1549、1337、1124、1303个,各占总碱基数的29.5%、25.16%、21.16%和24.52%。
2.2 基因序列的开放阅读框,编码氨基酸及保守区分析
开放阅读框(ORF)查找程序(http://www.ncbi.nlm. nih.gov/gorf)分析东方蜜蜂Vg基因cDNA全长序列有一个完整的开放阅读框。EditSeq分析其共编码1770个氨基酸,其理论等电点和相对分子量分别为6.612和201.23KD。使用Gblock 0.91软件分析其保守区序列占总序列的63%。
2.3 Vg基因系统发育分析
应用Blastn检索程序(http://www.ncbi.nlm.nih.gov)在GenBank数据库中将东方蜜蜂Vg基因cDNA全长序列进行核苷酸同源性分析,发现其与意大利蜜蜂Vg基因cDNA全长序列的相似性达到93%,其各自编码的氨基酸序列的相似性达到89%。东方蜜蜂Vg基因cDNA全长序列与大蜜蜂、小蜜蜂类似卵黄原蛋白基因(Vg-like)cDNA全长序列的相似性分别为 93%与90%,其编码氨基酸序列相似性分别达到90%与87%。
将克隆出的东方蜜蜂的Vg基因与GenBank上已发表的相关物种的Vg基因进行核苷酸序列比对,用Gblock 0.91软件选择保守区序列选择后,用ML法、MP法和BI法构建系统发育树,结果如下图:
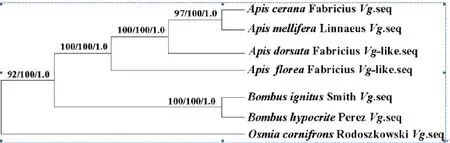
ML、MP和BI三种方法构建系统发育树
用三种方法(树上各分支的数据分别代表三种方法,ML、MP、BI)构建的系统发育树,可以看出,三种方法得到的结果一致,且置信度都很高,说明得到的结果非常可靠。系统发育树结果表明,在蜜蜂属内,东方蜜蜂与意大利蜂的亲缘关系最近,与大蜜蜂亲缘关系次之,与小蜜蜂的亲缘关系相对较远。东方蜜蜂作为蜜蜂属内的物种,与熊蜂属内的红光熊蜂、晓峰熊蜂关系较近,与壁蜂属内的角额壁蜂亲缘关系较远。
3 讨论
对Vg基因的研究始于1954年[8],由于其重要作用,被西方学者所重视。他们对昆虫中非蜜蜂科的天蚕、柞蚕、美国小蠊、浅色按蚊、米蛾,蜜蜂科内的意大利蜜蜂、普通熊蜂、苜蓿切叶蜂、角额壁蜂、丽蚜小蜂的Vg基因cDNA不仅进行了克隆,其表达特性的研究亦很深入,它的功能也得到了验证。在我国,从报道资料来看,目前对Vg基因的研究只限于棉铃虫、大草蛉、七星瓢虫及几种常见的寄生蜂[28~31]。Li et al.和李继莲等人分别于2010年和2012年成功克隆出晓峰熊蜂、红光熊蜂Vg基因cDNA全长序列[32,33],填补了国内对蜜蜂科内物种Vg基因克隆工作的空白。有关东方蜜蜂Vg基因的研究,目前仅见于2012年李志勇等人对东方蜜蜂不同日龄成蜂个体中卵黄原蛋白基因的转录模式的研究[34]。本研究以东方蜜蜂总RNA为模板,利用RT-PCR技术首次克隆了东方蜜蜂Vg基因cDNA全长序列,为今后进一步研究东方蜜蜂Vg的结构,Vg基因的功能和表达调控等奠定了基础。
东方蜜蜂Vg基因cDNA全长序列为5313bp,编码1770个氨基酸,序列分析发现其与GenBank中已提交的意大利蜜蜂Vg基因cDNA全长序列,发现其编码的开放阅读框数量以及编码的氨基酸数量相同[35]。其E值与意大利蜜蜂最小,表明其序列与意大利蜜蜂的同源性最高。而根据ML、MP和ML三种方法构建的系统发育树发现,东方蜜蜂与意大利蜜蜂位于同一分支,亲缘关系最近,结果与在Genbank中Blast的结果一致。
根据NCBI在线BLAST的结果,东方蜜蜂Vg基因cDNA全长序列与大蜜蜂、小蜜蜂类似Vg基因cDNA全长序列的相似性分别为93%与90%,结合三种方法进行的系统发育分析,我们推测:由美国贝勒医学院小蜜蜂全基因测序小组于2012年3月提交的小蜜蜂类似Vg基因的序列就是小蜜蜂的Vg基因的序列;2014年1月由美国冷泉港大蜜蜂全基因组测序实验室向GenBank提交的大蜜蜂类似Vg基因的序列就是大蜜蜂的Vg基因序列。
由系统发育分析本研究得出,蜜蜂科内东方蜜蜂、意大利蜂、大蜜蜂、小蜜蜂作为蜜蜂属内的物种,与熊蜂属的关系较近,与壁蜂属中的角额壁蜂的亲缘关系次之。Vg基因参与胚胎营养,巢内工作向采集工作的转变以及对花粉与花蜜的采集偏好,上述蜂种在以上方面存在的差异是否与Vg基因相关,需要进一步验证。
总之,本研究首次克隆了东方蜜蜂Vg基因(据我们所知,这是迄今为止第一次报道的东方蜜蜂Vg基因cDNA全长序列,该基因的cDNA全长序列也已提交到GenBank),并对该基因进行了基本的生物信息学分析。但我们的研究还不够深入,许多问题有待解决,如Vg基因在东方蜜蜂中的表达情况,对行为有何调控作用等还有待于进一步研究。弄清Vg基因在东方蜜蜂体内所起的作用,将对保护我国这一宝贵蜂种产生积极的作用。
[1]Byrne B M,Gruber M,Ab G.The evolution of egg yolk proteins.Prog.Biophys.Mol.Biol,1989,53:33-69.
[2]Raikhel AS,Dhadialla TS,Accumulation of yolk proteins in insect oocytes.Annual Review of Entomology,1992,37:217-251.
[3]Sappington TW,Raikhel AS,Molecular characteristics of insect vitellogenins and vitellogenin receptors.Insect Biochem Mol Biol, 1998,28:277-300.
[4]Snigirevskaya E S,Raikhel AS.Receptor mediated endocytosis of yolk proteins in insect oocytes.Reproductive Biology of Invertebrates,vol.XII.Part B.Science Publishers,Inc,Enfield,USA-Plymouth UK,2005,pp.199-228.
[5]Hagedorn H H,Maddison D R,Tu Z The evolution of vitellogenins,cyclorrhaphan yolk proteins and related molecules.Advances in Insect Physiology,1998,27:335-384.
[6]Sappington T W,Oishi K,Raikhel A S.Reproductive Biology of Invertebrates,vol.XII.Part A.Science Publishers,Inc.,Enfield, USA-Plymouth UK,2002,pp.69-101.
[7]Tufail M,Takeda M.Molecular cloning,characterization and regulation of the cockroach vitellogenin receptor during oogenesis. Insect Molecular Biology,2005,14:389-401.
[8]Tefler W H.Immunological studies of insect metamorphosis.II. The role of a sex-limited blood protein in egg formation by the Cecropia silkworm.The Journal of General Physiology,1954,37:538-558.
[9]Pan ML,Bell WJ,Telfer WH.Vitellogenin blood protein synthesis by insect fat body.Science,1969,165:393-394.
[10]Engelmann,F.Endocrinology of Insects.Alan R.Liss,New York,1983,pp.259-270.
[11]Wyatt G R,Davey K G.Cellular and molecular actions of juvenile hormone.II.Roles of juvenile hormones in adult insects.Advances in Insect Physiology,1996,26:1-155.
[12]Giorgi F,Snigirevskaya E S,Raikhel A S.Progress in Vitellogenesis.Reproductive Biology of Invertebrates,vol.XII.Part B. Science Publishers,Inc.,Enfield,USA-Plymouth UK,2005,pp.33-68.
[13]Belles X.Endocrine effectors in insect vitellogenesis.Recent Advances in Arthropod Endocrinology.Cambridge University Press, Cambridge,1998,pp.71-90.
[14]Belles X.Vitellogenesis directed by juvenile hormone.Reproductive Biology of Invertebrates,vol.XII.Part B.Science Publishers,Inc.,Enfield,USA-Plymouth UK,2005,pp.157-197.
[15]Engelmann F.Insect vitellogenin:identification,biosynthesis, and the role in vitellogenesis.Advances in Insect Physiology,1979, 14:49-108.
[16]Trenczek TA,EngelsW.Occurring of vitellogenin in drone honeybees.International Journal of Invertebrate Reproduction and Development,1986,10:307-311.
[17]Valle,D.Vitellogenesis in insects and other groups a review. Memorias DoInstituto Oswaldo Cruz,1993,88:1-26.
[18]Piulachs M D,Guidugli K R,Barchuk A R,et al.The vitellogenin of the honey bee Apis mellifera:structural analysis of the cDNA and expression studies.Insect Biochemistry and Molecular Biology,2003,33:49-465.
[19]Guidugli K R,Piulachs M D,Belle s X.Vitellogenin expression in queen ovaries and in larvae of both sexes of Apis mellifera. Archives of Insect Biochemistry and Physiology,2005,59:211-218.
[20]Hartfelder K,Engels W.Social insect polymorphism:Hormonal regulation of plasticity in development and reproduction in the honeybee.Curr Top Dev Biol,1998,40,45-77.
[21]Rutz W,Liischer M.The occurrence of vitellogenin in workers and queens of Apis mellifica and the possibility of its transmission to the queen.J Insect Physiol,1974,20:897.
[22]Engels,W.and Fahrenhorst,H.Alters-und kastenspezifische Veranderungen der Haemolymph-Protein-Spektren bei Apis mellificia.Dev Genes Evol,1974,174:285-296.
[23]Amdam G V,Simoes Z L,Hagen A,et al.Hormonal control of the yolk precursor vitellogenin regulates immune function and longevity in honeybees.Exp Gerontol,2004,39:767-773.
[24]Amdam GV,Aase AL,Seehuus SC,et al.Social reversal of immunosenescence in honey bee workers.Experimental Gerontology, 2005,40:939-947.
[25]Seehuus SC,Norberg K,Gimsa U,et al.Reproductive protein protects functionally sterile honey bee workers from oxidative stress. PNAS,2006.
[26]Fluri P,Wille H,Gerig L,et al.M.Juvenile hormone,vitellogenin,and hemocyte composition in winter worker honeybees (Apis mellifera).Experientia,1977,33:1240-1241.
[27]Engels,W.Occurrence and Significance of Vitellogenins in Female castes of Social Hymenoptera.Integr Comp Biol,1974,14: 1229-1237.
[28]王智健,牛长缨.UV照射对棉铃虫产卵量及HMG—CoA还原酶和卵黄原蛋白基因表达量的影响 [J].华中农业大学学报, 2014,(2):46-50.
[29]刘昌燕,曾凡荣.大草蛉卵黄原蛋白基因功能研究[D].中国农业科学院,2013.
[30]龚和,张建中,翟启慧.七星瓢虫卵黄原蛋白的合成[J].动物学集刊,1982(2):175-180.
[31]董胜张,叶恭银,胡萃.不同类群寄生蜂卵黄原蛋白的检测和免疫相关性分析[J].昆虫学报,2009,52(9):1024-1027.
[32]Li J,Huang J,Cai W,ZhaoZ.The vitellogenin of the bumblebee.Bombushypo crita:studies on structural analysis of the cDNA and expression of the mRNA.J.Comp.Physiol.B,Biochem.Syst. Environ.Physiol,2010,180(2):161-170.
[33]李继莲,吴杰,彭文君.红光熊蜂卵黄原蛋白基因的cDNA全长序列克隆和表达分析应用.昆虫学报,2012,49(5):1132-1139.
[34]李志勇,薛运波,任炳忠.东方蜜蜂卵黄原蛋白基因转录模式 [A].国家蜂产业技术体系蜜蜂育种与授粉功能研究室学术研讨会暨中国养蜂学会蜜蜂育种专业委员会第四届第一次会议暨中国养蜂学会蜜源与蜜蜂授粉专业委员会第五届第一次会议论文汇编[C],2012.
[35]The Honeybee Genome Sequencing Consortium.Insights into social insects from the genome of the honeybee Apis mellifera.Nature,2006,443(7114):931-949.
Cloning and fundamental bioinformatical characteristics of Vitellogenin cDNA in honeybees Apis cerana
Wang Xiuxiu1*,Yang Minghua1,2*,Li Chang3,Kuang Haiou3He Shaoyu3,Li Yahui3
(1.College of Animal Science and Technology,Yunnan Agricultural University,Kunming 650201;2.Yunnan Key Laboratory of Animal Nutrition and Feed,Yunnan Agricultural University,Kunming 650201;3.College of Food Science and Technology,Yunnan Agricultural University,Kunming 650201)
Vitellogenin (Vg)gene plays a pivotal role in ovarian development,embryotrophy,labor division,behavior regulation,climate adaptation and life extending.However,the fundamental data of Vg gene in Eastern honeybees (Apis cerana)has rarely been reported.In the present study,the full-length cDNA of Vg in Apis cerana was cloned,sequenced and analyzed bioinformatically.The results indicate that:the full-length cDNA of Vg in Apis cerana was composed of 5313bp which encode 1770 amino acids.The Vg cDNA sequence of Apis cerana was presented to GenBank with the accession number of KP119837.The sequence identity of the Vg cDNA and the encoding amino acid for Apis cerana and Apis mellifera were 93%and 89%,respectively.The conserved cDNA sequence accounted for 63%in the whole sequence.Phylogenetic tree of Vg was constructed with 4 honeybee species in Apis genus whose Vg sequences are known.The genetic relationship analysis showed that Apis cerana is genetically closest to Apis mellifera,while it is further and furthest to Apis dorsata and Apis florea,respectively.This study will lay a foundation for the further study of Vg function in Apis cerana.
Apis cerana,vitellogenin,Vg,cDNA,cloning,bioinformatics.
现代农业(蜜蜂)产业技术体系建设专项资金资助(CARS-45-kxj14)。
*对本研究贡献等同,并列第一作者。王秀秀(1989-),女,河南人,在读硕士,从事蜜蜂分子生物学研究,E-mail:cera0393@163. com;杨明华(1967-),女,云南人,硕士,实验师,从事动物分子营养与代谢调控研究,E-mail:yangmh85@hotmail.com.
和绍禹(1952-),男,教授,博士生导师,从事蜜蜂资源与授粉研究,E-mail:kmhsy@163.com;李亚辉(1967-),男,博士,教授,从事动物生殖与发育生物学研究,E-mail:kmliyh@163.com.
猜你喜欢
环球时报(2022-09-20)2022-09-20
娃娃乐园·综合智能(2021年4期)2021-07-20
今日农业(2021年6期)2021-06-09
快乐语文(2020年36期)2021-01-14
今日农业(2020年24期)2020-12-15
作文成功之路·小学版(2019年5期)2019-08-23
儿童故事画报·发现号趣味百科(2017年10期)2018-03-13
儿童故事画报·发现号趣味百科(2017年9期)2018-03-13
儿童故事画报·发现号趣味百科(2016年4期)2016-06-24
小资CHIC!ELEGANCE(2015年14期)2015-09-23
