
大肠杆菌K 235能量代谢扰动影响聚唾液酸合成的研究
2015-10-25 07:02闫霞吴剑荣郑志永詹晓北
生物技术通报 2015年9期
闫霞 吴剑荣 郑志永 詹晓北
(江南大学生物工程学院 糖化学与生物技术教育部重点实验室,无锡 214122)
大肠杆菌K 235能量代谢扰动影响聚唾液酸合成的研究
闫霞 吴剑荣 郑志永 詹晓北
(江南大学生物工程学院糖化学与生物技术教育部重点实验室,无锡214122)
通过不同装液量、不同氧化还原状态的碳源和添加NAD+的前体烟酸来调控大肠杆菌胞内的能荷、还原力水平,研究胞内核苷酸和聚唾液酸合成之间的关系。结果表明,聚唾液酸合成量越高,胞内UTP含量越低、UMP含量越高;同时,合成UTP、CTP消耗大量ATP使胞内能荷水平较低。与其他碳源的发酵相比,以山梨醇为碳源时,由于胞内NADH水平的提高使菌体合成的聚唾液酸量最高。添加烟酸可以提高胞内的NAD+水平,导致胞内的氧化水平提高,从而使胞内还原产物如聚唾液酸、琥珀酸、乳酸的产量降低甚至消失。
大肠杆菌;聚唾液酸;能荷;核苷酸;氧化还原势
聚唾液酸(Polysialic acid,PSA)是以N-乙酰神经氨酸为单体,以 α-2,8 和/或 α-2,9 糖苷键连接的聚合度在 8-400 的聚阴离子型线性多糖[1-4],通常存在于脊椎动物细胞的神经黏附分子(NCAM)末端和一些致病菌(如大肠杆菌K1株和奈瑟斯脑膜炎球菌)细胞表面。在脊椎动物体内,PSA参与细胞迁移、突触形成等生物功能且与神经系统的可塑性有关。由于PSA具有良好的生物相容性和生物可降解性,因此PSA可用作神经再生修复的组织工程支架材料[5]。另外,PSA在结构上具有较多羟基,从而具有高度亲水性,再加上其非免疫原性,使得聚唾液酸化的药物可以延长在血液循环中的半衰期,是目前发现可以替代聚乙二醇(PEG)的比较理想的蛋白类药物缓释材料[6]。
目前,生产PSA的唯一方法是通过微生物发酵,之后从发酵液中提取、纯化来制备[7,8],用于生产PSA的大肠杆菌(Escherichia coli)主要有K235和K1两株[9,10]。从1999年起,本实验室对利用E. coli K235生产PSA进行研究,通过诱变、培养基的优化、培养条件的优化,使PSA的产量较出发菌株提高了5倍[9,11]。但总体而言,PSA的产量仍较低,且菌株的PSA产量不稳定,难以满足产业化的需求。
我们前期研究发现E. coli K235发酵生产PSA受溶氧的严重影响[12],而氧作为电子传递链(Electron transport chain,ETC)的最终电子受体直接影响微生物细胞内的能荷(Energy charge,EC)和氧化还原状态。调节微生物细胞内能荷、还原力的方法有细胞改造和发酵工艺调节两种方法。其中,细胞改造方法包括:(1)通过基因工程操作使与NAD+/H代谢有关的酶过量表达[13,14];(2)将与NAD+/H竞争的代谢途径删除[15,16];(3)引入其他生物的NAD+/H再生系统[17,18]。发酵工艺调节方法包括:补加(1)外源电子受体例如富马酸[19]、乙醛[20];(2)不同氧化还原状态的碳源[21],如山梨醇、葡萄糖、葡萄糖酸钠;(3)NAD+的前体。本研究尝试利用不同氧化还原状态的碳源和NAD+合成的前体来调控菌体细胞内能荷、还原力水平,探索其与PSA合成之间的联系,用于指导PSA的生产合成。
1 材料与方法
1.1 材料
大肠杆菌(Escherichia coli CCTCC M208088),由江南大学糖化学与生物技术教育部重点实验室保藏。ATP、ADP、AMP、UMP、UTP、NAD+等购于Sigma-Aldrich公司,其他药品由国药试剂提供。
斜面培养基(g/L):氯化钠5,胰蛋白胨10,牛肉膏3,琼脂粉20,pH7.2-7.4。种子培养基(g/L):氯化钠5,胰蛋白胨10,牛肉膏3,pH7.2-7.4。发酵培养基(g/L):山梨醇40,硫酸铵4.94,磷酸氢二钾(三水)26.2,胰蛋白胨1.5,硫酸镁0.9,pH7.8(灭菌前)。
1.2 方法
1.2.1 斜面培养 将保存于4℃ 冰箱里的菌种,挑取一环到准备好的固体斜面培养基上,在37℃ 恒温箱内恒温培养12 h。
1.2.2 种子培养 将斜面培养活化后的种子接种两环于250 mL 三角瓶中(内装50 mL种子培养基)后,放置于摇床中培养,摇床条件为转速250 r/min、温度37℃,培养8-10 h。
1.2.3 发酵培养 将准备好的种子培养基按照4%的接种量接种于250 mL三角摇瓶(内装35 mL发酵培养基)后,250 r/min、37℃培养60 h。
1.2.4 烟酸对PSA发酵的影响 准确称取1 g烟酸溶于1 L蒸馏水中,配制成1 g/L的烟酸溶液,灭菌待用。向分装灭菌的1.2的发酵培养基中(35 mL/250 mL)分别加入上述1 g/L的烟酸溶液175 μL、350 μL、525 μL、700 μL,配制成含烟酸为5 mg/L、10 mg/L、15 mg/L、20 mg/L的发酵培养基,研究烟酸对PSA发酵的影响。
1.2.5 分析方法
1.2.5.1 生物量测定 将样品稀释至一定倍数后,测定600 nm下吸光值,然后对照 OD600吸光度-菌体干重的标准曲线计算生物量,OD600=1.0相当于 0.4 g/L 细菌干重。
1.2.5.2 PSA的测定 对PSA的测定采用间苯二酚法[12]。
1.2.5.3 碳源及有机酸含量的测定 发酵液离心后即得待测样品,利用高效液相色谱(LC-2010A,岛津)测定,色谱柱为Aminex HPX-87H(9 μm,300 mm×7.8 mm,Biorad);流动相为5 mmol/L硫酸;流动相流速0.6 mL/min;示差检测器;柱温35℃;进样量5 μL。
1.2.5.4 胞内核苷酸的测定 样品的处理:取4 mL对数中期发酵液,经4℃,12 000 r/min离心10 min后,弃去上清,得到菌体,将获得的菌体悬浮于2 mL预冷的0.1 mol/L pH7.4的磷酸钾缓冲液中,超声波破碎(超声1 s,间隔1 s,)10 min,之后经4℃,12 000 r/min 离心10 min,得到的上清于HPLC中进行测定。
色谱条件:色谱柱为InertSustain C18柱(5 μm,4.6 mm×250 mm,Shimadzu);流动相为20%乙腈与含20 mmol/L四丁基溴化铵的0.4 mol/L pH7.0磷酸钠缓冲液以1∶1混合;流动相流速0.5 mL/min;紫外检测器;检测波长254 nm;柱温25℃;进样量10 μL。
2 结果
2.1 装液量对PSA发酵的影响
2.1.1 装液量对PSA合成和菌体生长的影响 由图1可以看出,与装液量35 mL的发酵结果相比,其它装液量下菌体的PSA产量和生长状况都较差,尤其在装液量大时更为明显。当装液量为50 mL时PSA产量和生物量分别为1.08 g/L和3.02 g/L,而装液量为35 mL时,PSA产量和生物量分别达1.51 g/L和4.07 g/L。山梨醇消耗与PSA产量、菌体生长趋势相一致。如图1所示,当装液量为35 mL,菌体的山梨醇消耗量最大,为22.40 g/L。
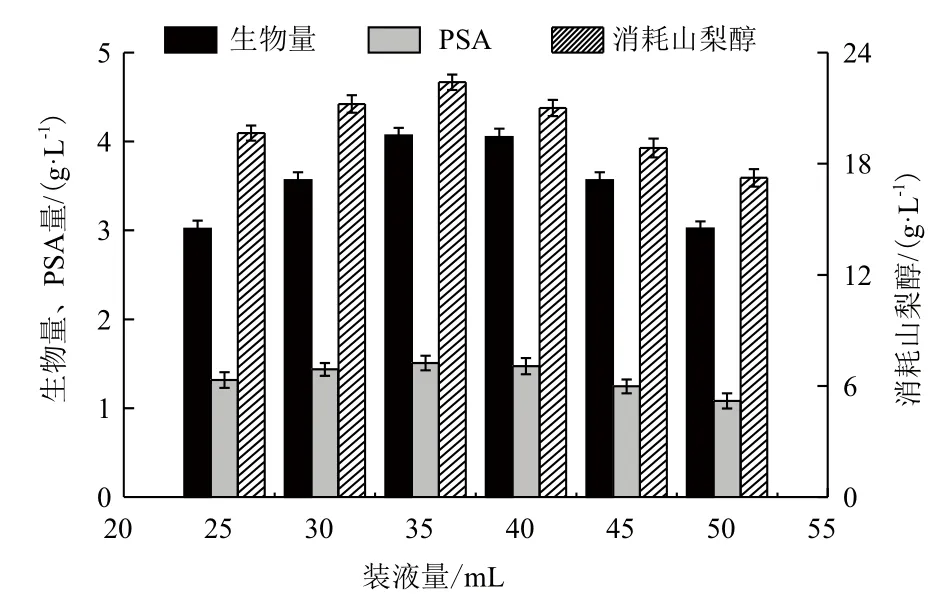
图1 不同装液量下的发酵参数
2.1.2 装液量对有机酸合成的影响 不同装液量下有机酸的合成情况如图2所示。在装液量小的情况下,柠檬酸合成量较高;当装液量为25 mL时,柠檬酸的合成量最高,为62.33 mmol/L。而当装液量分别为35 mL、40 mL、45 mL、50 mL时,柠檬酸的合成量比较接近,为55 mmol/L左右。获得PSA产量最高的装液量(35 mL)下,乙酸的合成量也最低(1.82 mmol/L),远低于其它装液量下的乙酸合成量,表明该条件下供氧充足,消耗的山梨醇用于菌体生长和PSA合成。在各个装液量下乳酸、琥珀酸的合成量较低或无法检测到,而在所有的装液量下没有乙醇的合成(数据没有给出)。

图2 不同装液量下的有机酸合成量
2.1.3 装液量对胞内核苷酸的影响 根据上述结果可知,E. coli合成PSA受溶氧的影响,而氧作为ETC的最终电子受体与氧化磷酸化产生能量密切相关。在不同装液量下胞内能荷与PSA合成的关系如图3所示,随着摇瓶装液量的增大,胞内ATP含量缓慢下降,从25 mL装液量的3.22 μM/g·cell下降到50 mL装液量的2.65 μM/g·cell,而EC水平在装液量为35 mL时最低(0.22),此时PSA的产量最高。
在E. coli合成PSA的过程中,ATP除了用于菌体生长外,还用于合成UTP、CTP、GTP等高能物质。其中UTP用于UDP-葡萄糖胺和CTP的生成,CTP用于活化PSA合成前体——N-乙酰神经氨酸(Neu5Ac)和2-酮基-3-脱氧辛酸(KDO),因此菌体细胞内UTP、CTP的含量也将影响PSA合成。装液量对胞内UTP、UMP的影响如图3所示,随着装液量的增加,胞内UTP的量先下降后上升,在装液量为35 mL时菌体细胞内的UTP含量最低,为3.38 μM/g·cell。而菌体细胞内UMP的含量随装液量的增加先上升后下降,在装液量为35 mL时UMP含量最高,为83.19 μM/g·cell。菌体合成PSA消耗的UTP转化为UDP、UMP,从而使UMP的量明显高于其他装液量。
根据上述分析,菌体PSA合成和溶氧有紧密的联系,而氧气在代谢过程中作为电子受体接受通过ETC传递的质子并最终被氧化为水,伴随氧气的氧化同时释放能量,同时携带电子的载体是NADH,其与胞内氧化还原水平密切相关,因此胞内能荷、还原力的变化存在一定的关联性。故初步确定PSA的合成与能荷、还原力有密不可分的关系。
2.2 不同氧化还原状态的碳源对PSA发酵的影响
2.2.1 碳源对PSA合成和菌体生长的影响 分别以40 g/L山梨醇、葡萄糖、木糖、葡萄糖酸钠为碳源发酵生产PSA,结果如图4所示。发现E. coli K235可以利用不同的碳源进行菌体生长,利用葡萄糖酸钠为唯一碳源时菌体干重(1.66 g/L)仅是对照组(4.07 g/L)的40.8%。与其他碳源相比,山梨醇是生产PSA最有效的碳源。在以葡萄糖、木糖、葡萄糖酸钠为唯一碳源时,菌体的PSA产量极低,甚至无法合成PSA,而以山梨醇为唯一碳源时,PSA产量最高。此结果表明,菌体细胞的还原状态较高时更有利于PSA的生成。
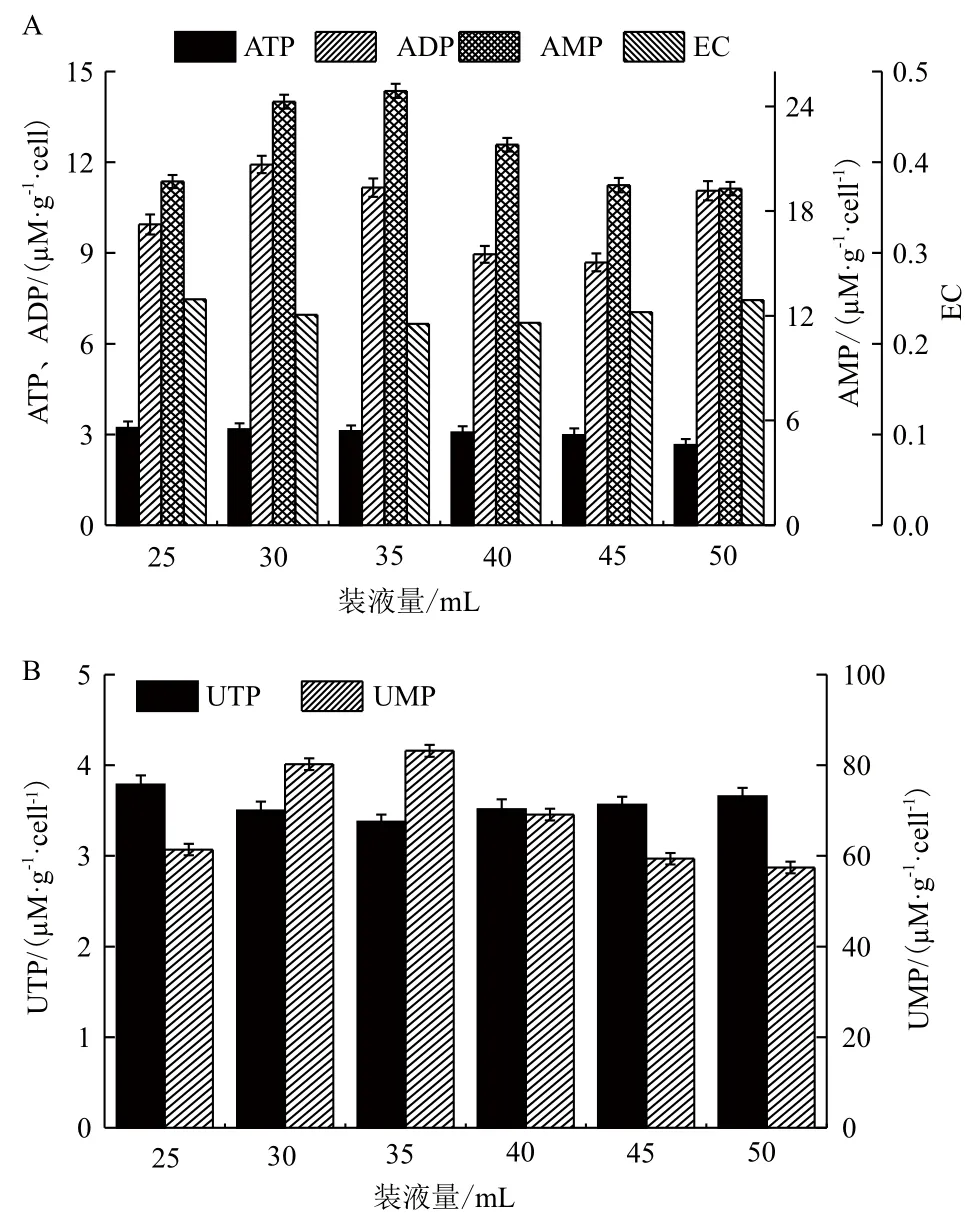
图3 不同装液量下的胞内能荷、UTP、UMP
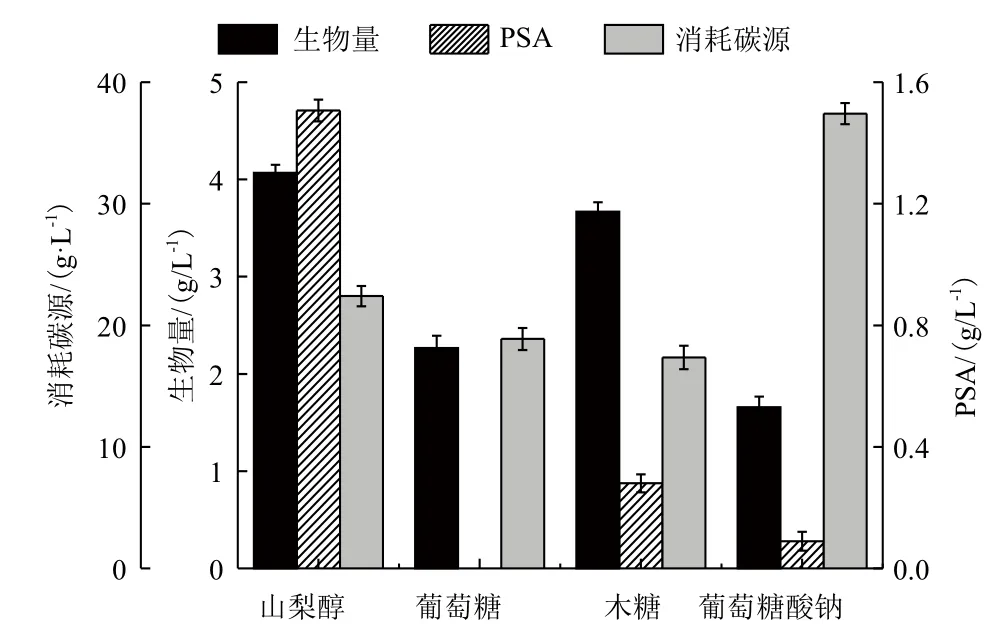
图4 不同碳源下的发酵参数
在消耗碳源方面,当以山梨醇、葡萄糖、木糖为碳源时,发酵结束时碳源的消耗量分别为22.40 g/L、18.88 g/L和17.35 g/L,消耗碳源量比较接近;而以葡萄糖酸钠为碳源时,发酵结束后葡萄糖酸钠的消耗量达到37.41 g/L,碳源基本全部耗尽,如图4所示。利用不同碳源发酵生产PSA时,山梨醇、葡萄糖、木糖的消耗主要在发酵过程的前25 h,而葡萄糖酸钠为碳源时,其碳源消耗速度非常快,在发酵进行仅12 h时,葡萄糖酸钠的消耗量将近30 g/L。在以葡萄糖酸钠作为碳源时,葡萄糖酸钠的消耗量虽然很高,但是菌体浓度和PSA产量都较低,且乙酸大量合成。
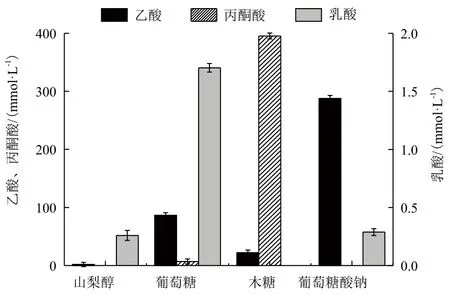
图5 不同碳源下有机酸的合成量
2.2.2 碳源对有机酸合成的影响 在呼吸方式方面,当以山梨醇为碳源时,菌体主要进行有氧呼吸,无氧呼吸产物极少,发酵液中乙酸、乳酸的含量分别仅为1.82 mmol/L和0.26 mmol/L(图5)。而当以葡萄糖、木糖、葡萄糖酸钠为碳源时,无氧呼吸明显加强,发酵液中存在大量被氧化的副产物如乙酸,无氧呼吸代谢产物量明显高于以山梨醇为碳源的发酵液中无氧呼吸代谢产物量。以葡萄糖酸钠和葡萄糖为碳源时,发酵液中乙酸的量分别高达287.93 mmol/L和86.57 mmol/L(图5)。而乙酸的生成伴随1分子ATP的生成,这样胞内ATP水平有所提高,更有利于菌体的生长。另外,一般情况下,乙酸的大量生成是由糖酵解途径和三羧酸循环的不平衡引起的。在以葡萄糖或葡萄糖酸钠为唯一碳源的情况下,除了积累大量的乙酸外,还有较多的乳酸生成(积累的乳酸量分别为1.70 mmol/L和0.29 mmol/L),比以山梨醇或木糖为碳源的乳酸积累量要高。而乳酸的生成意味着从丙酮酸到乙酰-CoA这一步的代谢速率降低,从而使糖酵解途径和三羧酸循环之间的不平衡降低。
当以木糖为碳源时,发酵液中存在大量的丙酮酸,高达395.4 mmol/L,没有乳酸的生成,而其他碳源下的发酵液中并无丙酮酸的积累,如图5所示。木糖为碳源时发酵液中乙酸的积累相对于葡萄糖、葡萄糖酸钠为碳源时低。由于木糖在代谢过程中主要经过糖酵解途径生成丙酮酸,在丙酮酸节点只有小部分经乙酰-CoA转化为乙酸。在E. coli的代谢过程中,由丙酮酸生成乙酰-CoA时伴随甲酸的生成,而在4种不同氧化还原状态的碳源下发酵液中都未检测到有甲酸的积累,这可能是由于无氧呼吸形成的甲酸在甲酸脱氢酶(Formate dehydrogenase,FDHF)的催化下分解为二氧化碳和氢气。同时4种碳源下柠檬酸的合成量较接近、琥珀酸的合成量较低(数据未给出)。
2.2.3 不同碳源对胞内核苷酸的影响 分别以山梨醇、葡萄糖、木糖、葡萄糖酸钠为碳源进行PSA发酵时,最终PSA合成量明显不同,测定对数中期胞内的EC水平,结果如图6所示。与对照(以40 g/L山梨醇为碳源的发酵)相比,以葡萄糖或葡萄糖酸钠为碳源时,胞内的EC水平较高,分别为0.42和0.40,远高于对照的0.22。通过以上分析可知,当以葡萄糖或葡萄糖酸钠为碳源时,菌体在代谢过程中生成浓度较高的乙酸,而乙酸的生成也偶联合成ATP,所以当葡萄糖或葡萄糖酸钠作为碳源时,胞内的ATP、EC水平相对较高。当以木糖为碳源时,由于菌体生成大量的丙酮酸,积累的丙酮酸并未通过无氧或有氧呼吸的方式释放能量,从而导致以木糖为碳源时,胞内的ATP水平较低,且EC水平最低(0.21)。以葡萄糖酸钠为碳源时,虽然生成乙酸量和生成乙酸偶联生成的ATP量要远远高于以葡萄糖为碳源的情况,但由于其生成PSA量要高于后者,且在PSA的合成中需要大量的能量,最终导致胞内ATP、EC水平低于以葡萄糖为碳源的发酵。

图6 不同碳源对胞内核苷酸的影响
PSA的合成需要UTP的参与并生成UDP-葡萄糖胺,足量的UTP是合成PSA的基础。在4种不同的碳源下,以葡萄糖为碳源时,胞内的UTP含量最高(5.85 μM/g.cell)。此时没有PSA的合成,所以UTP可以在胞内积累,相应UMP的量在胞内减少,如图6所示。同理,在以葡萄糖、木糖、葡萄糖酸钠为碳源时,PSA产量越低,胞内UTP含量越高,UMP含量越低。当以山梨醇为碳源时,菌体进行有氧呼吸生成的ATP激活UTP合成代谢中的天冬酸氨基甲酰转移酶,从而导致胞内的UTP含量相对较高,且合成PSA时消耗的UTP使胞内的UMP、UDP含量也有所提高,胞内的UMP量高达83.2 μM/g.cell。
2.3 添加NAD+的前体烟酸对PSA发酵的影响
2.3.1 烟酸对PSA合成和菌体生长的影响 向以山梨醇为碳源的发酵培养基中添加不同浓度的NA,研究NA对PSA发酵的影响。当向发酵培养基中添加NA后,PSA的产量会下降(图7),而且生物量、消耗山梨醇量也会随之下降。向培养基中加入不同浓度的NA后,PSA产量从对照(没有添加NA)的1.51 g/L下降到0.5 g/L左右,生物量同样出现明显的下降趋势。NA的添加使山梨醇的消耗量明显下降,当添加NA的量为10 mg/L时,山梨醇消耗量仅为8.03 g/L,比对照的山梨醇消耗量下降了64.2%。加入NA后PSA产量和生物量的下降可能是由于山梨醇的消耗量大幅度降低、微生物细胞的分解代谢受阻所致。
2.3.2 NA对有机酸合成的影响 微生物细胞中存在大量辅因子,如ATP、ADP、AMP、NADH、NAD+、NADPH、NADP+、乙酰-CoA及其衍生物、维生素和微量元素等,这些辅因子是大量生化反应的基础。作为辅因子NAD+/H参与许多生化反应,胞内NAD+/H的改变会影响细胞生长和代谢产物的合成,甚至导致代谢流发生改变。当向发酵培养基中添加NA后,使还原性代谢产物(聚唾液酸、琥珀酸、乳酸)的产量降低甚至消失,如图8所示。当加入的NA量为5 mg/L时,琥珀酸和乳酸的合成量从对照的0.14 mmol/L、0.26 mmol/L分别下降到0.02 mmol/L、0.03 mmol/L。当NA的添加量高于5 mg/L时,菌体就不再合成琥珀酸和乳酸。与琥珀酸、乳酸不同的是,当加入NA后,即使浓度较高,仍有不需要消耗NADH的乙酸正常合成。乙酸的合成使细胞减少对NADH的利用量,从而导致细胞氧化还原状态的平衡。添加NA后,菌体合成的柠檬酸量稍有降低(数据未给出)。

图7 烟酸对PSA发酵的影响

图8 烟酸对有机酸合成的影响
2.3.3 NA对胞内核苷酸的影响 向山梨醇的发酵培养基中添加不同浓度的NA,其中以不含NA的培养条件为对照,取对数中期的细胞,测定胞内的EC、还原力水平。由于E. coli可以NA作为合成NAD+的前体物质,与对照相比,当向发酵培养基中添加NA后,胞内的NAD+水平提高,如图9所示。还原力(NADH/NAD+)水平相对降低,此时菌体会减少对NADH的利用,从而导致琥珀酸、乳酸的合成量降低(合成琥珀酸、乳酸所需要的酶是依赖NADH的酶),如图10所示。
在向发酵培养基中添加NA后,菌体合成的乙酸量降低,而乙酸的生成偶联ATP的再生,同时,发酵培养基中较高浓度的NA导致胞内还原力水平降低后,抑制NADH通过ETC产生能量,故添加NA后菌体细胞内的ATP水平有所降低。如图9所示,当加入的NA量为15 mg/L时,胞内的ATP水平仅为2.05 μM/g.cell,比对照的低34.2%。故添加NA后,ATP的不足导致PSA的合成量明显降低,由于用于PSA合成的ATP量减少,胞内的EC水平要高于对照的0.22。

图9 烟酸对胞内能荷的影响
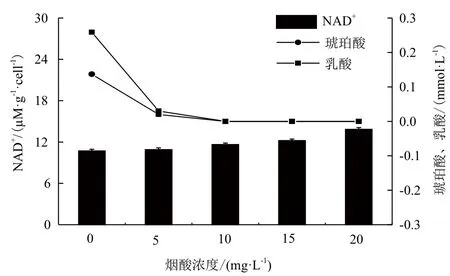
图10 烟酸对胞内NAD+量、菌体有机酸合成量的影响
在向发酵培养基中添加NA后,PSA合成量降低,故合成PSA消耗的UTP量降低,菌体细胞内积累的UTP量增加,如图11所示。当添加10 mg/L NA后,菌体细胞内的UTP量高达3.67 μM/g·cell,高于对照的3.38 μM/g·cell。与UTP的变化相反,当菌体由于添加NA使合成PSA的量降低时,菌体细胞内积累的UMP会降低,从对照的83.2 μM/g·cell下降到36.6 μM/g·cell。

图11 烟酸对胞内UTP、UMP的影响
3 讨论
本研究在利用不同氧化还原状态的碳源发酵生产PSA时,最终PSA产量明显不同,其中以山梨醇为碳源时PSA产量最高,这与山梨醇在代谢过程中产生较多的NADH使细胞处于较高的还原状态有关。Hong等[23]在利用E. coli工程菌生产琥珀酸时,选用山梨醇和葡萄糖分别作为碳源,结果表明山梨醇作为碳源提高琥珀酸产量与其提高胞内还原力(消耗1 mol山梨醇可以形成3 mol NADH,而同样消耗1 mol葡萄糖却只能产生2 mol NADH)有密切的关系,因为从PEP向琥珀酸的转化中需要2分子NADH,而在山梨醇的代谢过程中比葡萄糖可形成更多的NADH[24]。同样在琥珀酸的生产中,Lin等[21]在利用山梨醇为碳源时的产量比以葡萄糖为碳源时的高81%,而琥珀酸产量的提高是在胞内可利用的NADH量增加时产生,同时胞内NADH的增加也导致乙醇(在生成乙醇时需要消耗2分子NADH)的产量提高了62%。最近,Liu等[25]在利用E. coli生产莽草酸时,由于山梨醇较高的还原性使莽草酸的产量是用葡萄糖为碳源时的3.7倍。本研究中以葡萄糖、葡萄糖酸钠为碳源时胞内较高的ATP、EC水平与乙酸的生成有关,因为乙酸的生成偶联ATP的合成;以木糖为碳源时,由于菌体生成大量的丙酮酸,积累的丙酮酸并未通过无氧或有氧呼吸的方式释放能量,从而导致以木糖为碳源时,胞内的ATP、EC水平都较低。
微生物会自动调节胞内的NADH水平,而对NADH/NAD+的比没有严格的要求[26,27]。向培养基中添加NA后,虽然胞内NAD+总量提高,但NADH水平基本维持平衡状态,从而导致胞内还原力水平下降。由于PSA的合成需要在较高的还原环境下进行,所以向培养基中加入NA后,PSA产量会降低。Liu等[20]利用光滑球拟酵母发酵生产丙酮酸时,当向发酵培养基中添加8 mg/L NA后,胞内的NAD+量从未添加NA的13.1 mg/g DCW提高到28.6 mg/g DCW,从而降低了胞内NADH/NAD+水平;由于在丙酮酸合成代谢中需要消耗NAD+,故导致葡萄糖的消耗速率和丙酮酸的产量分别增加了48.4%、29%。
微生物细胞内NADH/NAD+水平的改变会导致代谢产物组成的变化。Berrios-Rivera等[28]向E. coli内引入来自Candida boidinii的NADH再生途径,该途径中的甲酸脱氢酶(Formate dehydrogenase,FDH)可以将甲酸转化为二氧化碳,并伴随NADH的生成,而存在于E. coli内的FDH在催化过程中没有NADH的生成,故该异源代谢途径的引入可以提高菌体细胞内的还原水平。在厌氧和好氧条件下,异源NADH再生途径对代谢物的组成都产生显著的影响,在厌氧条件下主要形成还原代谢产物乙醇,而在好氧条件下由于胞内NADH的增多使菌体在有氧环境中却转向厌氧代谢,主要表现在乙醇、乳酸、琥珀酸等厌氧代谢产物的大量合成中。本研究向E. coli的发酵培养基中添加NA后,胞内的NAD+水平明显提高,此时菌体就会降低对NADH的利用。由于琥珀酸合成需要消耗2分子的NADH,乳酸的合成需要消耗1分子的NADH,菌体为了达到胞内氧化还原的平衡,减少了向琥珀酸、乳酸的代谢流量,当NA的添加量大于5 mg/L,菌体甚至停止合成琥珀酸、乳酸。
4 结论
可以利用装液量、不同氧化还原状态的碳源、烟酸等来调控菌体细胞内的核苷酸变化。当装液量为35 mL时,PSA产量达到最高值1.51 g/L,由于合成PSA消耗胞内大量的ATP,导致EC水平较低。较其他碳源而言,山梨醇高的还原性使其成为发酵生产PSA的最有效碳源。胞内NAD+、NADH量的改变会影响PSA合成。当胞内NAD+增多会使PSA合成量减少,反之胞内NADH增多会使PSA合成量增多。
[1] Rodriguez-Aparicio LB, Reglero A, Ortiz AI, et al. Effect of physical and chemical conditions on the production of colominic acid by Escherchia coli in a defined medium[J]. Applied Microbiology and Biotechnology, 1988, 27(5-6):474-483.
[2] Barker SA, Jones RG, Somers PJ. Improvements in the production and isolation of colominic acid[J]. Carbohydrate Research, 1967,3(3):369-376.
[3] Inoue S, Inoue Y. Development profile of neural cell adhesion molecule glycoforms with a varying degree of polymerization of polysialic acid chains[J]. The Journal of Biological Chemistry,2001, 276(34):31863-31870.
[4] Nakata D, Troy FA. Degree of polymerization(DP)of polysialic acid(polySia)on neural cell adhesion molecules(N-CAMS):development and application of a new strategy to accurately determine the DP of polySia chains on N-CAMS[J]. The Journal of Biological Chemistry, 2005, 280(46):38305-38316.
[5] Haile Y, Berski S, Drager G, et al. The effect of modified polysialic acid based hydrogels on the adhesion and viability of primary neurons and glial cells[J]. Biomaterials, 2008, 29(12):1880-1891.
[6] Chen C, Constantinou A, Chester KA, et al. Glycoengineering approach to half-Life extension of recombinant biotherapeutics[J]. Bioconjugate Chemistry, 2012, 23(8):1524-1533.
[7] Camino CC, Maria LJ. High production of polysialic acid by Escherichia coli K-92 grow in a chemically defined medium regulated by temperature[J]. Biological Chemistry Hoppe-Seyler, 1990, 371(11):1101-1106.
[8] 郭良栋, 钱世钧, 叶军, 等. 产多聚唾液酸的菌种筛选及产酸条件[J]. 微生物学报, 1998, 38(2):103-107.
[9] 贾薇. 聚唾液酸生产菌的诱变育种及发酵研究[D]. 无锡:无锡轻工大学, 1999.
[10]Rode B, Endres C, Ran C, et al. Large-scale production and homogenous purification of long chain polysialic acids from E. coli K1[J]. Journal of Bacteriology, 2008, 135(2):202-209.
[11]于军华. 聚唾液酸生产菌种的选育及其发酵工艺的研究[D].无锡:无锡轻工大学, 2000.
[12]张琦. 聚唾液酸发酵和分离纯化工艺的研究[D]. 无锡:江南大学, 2009.
[13]Berrios-Rivera SJ, San KY, Bennett GN. The effect of NAPRTase overexpression on the total levels of NAD, the NADH/NAD(+)ratio, and the distribution of metabolites in Escherichia coli[J]. Metabolic Engineering, 2002, 4(3):238-247.
[14] Heuser F, Schroer K, Lutz S, et al. Enhancement of the NAD(P)(H)pool in Escherichia coli for biotransformation[J]. Engineering in Life Sciences, 2007, 7(4):343-353.
[15]Geertman JM, van Maris AJ, van Dijken JP, et al. Physiological and genetic engineering of cytosolic redox metabolism in Saccharomyces cerevisiae for improved glycerol production[J]. Metabolic Engineering, 2006, 8(6):532-542.
[16]Zhang YP, Li Y, Du CY, et al. Inactivation of aldehyde dehydrogenase:A key factor for engineering 1, 3-propanediol production by Klebsiella pneumoniae[J]. Metabolic Engineering,2006, 8(6):578-586.
[17]Hou J, Lages NF, Oldiges M, et al. Metabolic impact of redox cofactor perturbations in Saccharomyces cerevisiae[J]. Metabolic Engineering, 2009, 11(4-5):253-261.
[18]Zhang Y, Huang Z, Du C, et al. Introduction of an NADH regeneration system into Klebsiella oxytoca leads to an enhanced oxidative and reductive metabolism of glycerol[J]. Metabolic Engineering, 2009, 11(2):101-106.
[19]de Graef MR, Alexeeva S, Snoep JL, et al. The steady-state internal redox state(NADH/NAD+)reflects the external redox state and is correlated with catabolic adaptation in Escherichia coli[J]. Journal of Bacteriology, 1999, 181(8):2351-2357.
[20]Liu LM, Li Y, Shi ZP, et al. Enhancement of pyruvate productivity in Torulopsis glabrata:Increase of NAD(+)availability[J]. Journal of Bacteriology, 2006, 126(2):173-185.
[21]Lin H, Bennett GN, San KY. Effect of carbon sources differing in oxidation state and transport route on succinate production in metabolically engineered Escherichia coli[J]. Journal of Industrial Microbiology and Biotechnology, 2005, 32(3):87-93.
[22]Svennerholm L. Quantitative estimation of sialic acids[J].Biochimica et Biophysica Acta, 1957, 24(4):604-611.
[23]Hong SH, Lee SY. Importance of redox balance on the production of succinic acid by metabolically engineered Escherichia coli[J]. Applied Microbiology and Biotechnology, 2002, 58:286-290.
[24]San KY, Bennett GN, Berrios-Rivera SJ, et al. Metabolic engineering through cofactor manipulation and its effects on metabolic flux redistribution in Escherichia coli[J]. Metabolic Engineering, 2002, 4:182-192.
[25]Liu XL, Lin J, Hu HF, et al. Metabolic engineering of Escherichia coli to enhance shikimic acid production from sorbitol[J]. World Journal of Microbiology and Biotechnology, 2014, 30(9):2543-2550.
[26]W impenn, JWT, Firth A. Levels of nicotinam ide adenine dinucleotide and reduced nicotinamide adenine dinucleotide in facultative bacteria and the effect of oxygen[J]. Journal of Bacteriology, 1972, 111(1):24-32.
[27]Matin A, Gottschal JC. Influence of dilution rate on NAD(P)and NAD(P)H concentrations and ratios in a Pseudomonas sp. grown in continuous culture[J]. Journal of General Microbiology, 1976,94(2):333-341.
[28]Berrıos-Rivera SJ, Bennett GN, San KY, et al. Metabolic engineering of Escherichia coli:increase of NADH availability by overexpressing an NAD+-dependent formate dehydrogenase[J]. Metabolic Engineering, 2002, 4(3):217-229.
(责任编辑 李楠)
Influence of Perturbation of Energy M etabolism in Escherichia coli K 235 on Biosynthesis of Polysialic Acid
Yan Xia Wu Jianrong Zheng Zhiyong Zhan Xiaobei
(Key Laboratory of Carbohydrate Chemistry & Biotechnology of Ministry of Education,School of Biotechnology,Jiangnan University,Wuxi214122)
Different medium volumes, carbon sources with varied redox states and adding nicotinic acid, and the precursor of NAD+were used to adjust the intracellular energy charge and reduction of Escherichia coli in order to investigate the relationships between biosynthesis of polysialic acid and intracellular nucleotides. The results showed that the higher the production of polysialic acid was, the lower UTP was, but higher UMP was. Meanwhile, energy charge level was lower because the synthesis of UTP and CTP consumed a lot of ATP. Using sorbitol as carbon source, more polysialic acid was produced than those of fermentation with glucose, xylose and sodium gluconate due to the increase of NADH. Addition of nicotinic acid to the fermentation medium elevated intracellular NAD+, which led to the raise of intracellular oxidative stress,sequentially the decrease of the reducing metabolites such as polysialic acid, succinate and lactate, and even the vanish of them.
Escherichia coli;polysialic acid;energy charge;nucleotides;redox potential
10.13560/j.cnki.biotech.bull.1985.2015.09.030
2014-12-19
国家“863”计划项目(2012AA021505),江苏省自然科学基金项目(BK2011158)
闫霞,女,硕士研究生,研究方向:发酵工程;E-mail:13643544548@126.com
吴剑荣,男,副教授,硕士生导师,研究方向:工业微生物;E-mail:kinowu@jiangnan.edu.cn
猜你喜欢
中国食用菌(2021年10期)2021-11-04
石油化工(2021年9期)2021-10-18
传染病信息(2021年6期)2021-02-12
化工设计通讯(2020年5期)2020-01-12
食品工业科技(2019年24期)2019-02-18
中成药(2017年12期)2018-01-19
福建轻纺(2017年12期)2017-04-10
农业工程学报(2016年9期)2016-12-19
中国塑料(2016年5期)2016-04-16
陕西科技大学学报(2014年1期)2014-06-27
