
里氏木霉α-半乳糖苷酶基因agl2在毕氏酵母中高效表达
2015-12-27 00:50钟开新陈丽芝李阳源
微生物学杂志 2015年4期
钟开新, 陈丽芝, 李阳源
(广东溢多利生物科技股份有限公司,广东 珠海 519060)
里氏木霉α-半乳糖苷酶基因agl2在毕氏酵母中高效表达
钟开新, 陈丽芝, 李阳源*
(广东溢多利生物科技股份有限公司,广东 珠海 519060)
通过反转录PCR从里氏木霉RutC-30 基因中克隆到α-半乳糖苷酶基因agl2,将其构建到具有AOX1强启动子的表达载体pPIC9K中,线性化后转化到毕氏酵母GS115中进行表达。结果表明agl2基因在毕氏酵母GS115中得到了高效表达,在50 L发酵罐中高密度发酵168 h,酶活达最高值1 106 U/mL。经PCR产物鉴定,结果表明agl2基因已整合到毕氏酵母GS115基因组中。
α-半乳糖苷酶;里氏木霉;毕氏酵母;高密度发酵
α-半乳糖苷酶的应用涉及饲料工业和制糖工业[1]、血型改造[2-3]、基因表达检测[4]等方面。特别在饲料工业中,α-半乳糖苷酶能催化α-半乳糖苷类寡糖如棉子糖、水苏糖等动物难以消化的抗营养因子的水解而作为添加剂加入到豆粕饲料中[5]。目前我国α-半乳糖苷酶的商品化生产还很少且价格昂贵,开发高酶活、廉价的α-半乳糖苷酶作为饲料添加剂具有巨大的经济价值。毕氏酵母作为一种新型外源基因表达系统,具有一套与真核生物极其相似的分泌途径,是一种非常理想的真核蛋白表达系统。到目前为止,来源于青霉、曲霉、扬奇青霉等的α-半乳糖苷酶实现了在毕氏酵母中高效表达,其发酵酶活相比原菌株发酵高出几十倍之多,且酵母自身产生的分泌蛋白较少,从而有利于后续的分离纯化。本实验室从里氏木霉RutC-30中克隆到α-半乳糖苷酶的编码基因agl2,利用pPIC9K的强启动子AOX1构建pPIC9K-agl2表达载体,并转化毕氏酵母GS115,实现了AGL2的异源表达,并进行了高密度发酵,为α-半乳糖苷酶的基因工程改良研究和工业化生产提供参考。
1 材料与方法
1.1 材料
1.1.1 菌种与质粒 里氏木霉RutC-30来自中科院微生物研究所菌种保藏中心,大肠埃希菌Top10为本实验保存;毕氏酵母GS115和pPIC9K质粒均购自上海Invitrogen公司。
1.1.2 培养基 LB培养基(g/L):胰蛋白胨 10,酵母膏 5,NaCl 10,根据需要添加50 mg/mL氨苄青霉素;察氏培养基(g/L):NaNO33,K2HPO41,MgSO40.5,KCl 0.5,FeSO40.01,蔗糖30。培养基YPD和BMGY均参考Invitrogen公司操作手册配制。
1.1.3 工具酶和主要试剂PfuDNA聚合酶、TaqDNA聚合酶、T4DNA连接酶均购自Fermentas公司;限制性内切酶购自NEB公司;抗生素G418,对硝基酚、对硝基酚-α-D-半乳糖苷为Sigma公司产品。真菌DNA提取试剂盒、PCR产物试剂盒、胶回收试剂盒等均购于上海生工公司。
1.2 方法
1.2.1 α-半乳糖苷酶基因克隆 将里氏木霉RutC-30菌株接种于察氏培养基中,30 ℃、250 r/min培养3 d,菌液离心收集菌丝,用无菌水洗涤3次,再次离心后液氮冷冻研磨,根据真菌RNA提取试剂盒提取总RNA,再经反转录试剂盒转录成cDNA。根据GenBank中已公布的里氏木霉α-半乳糖苷酶基因agl2(登录号:Z69254)序列,设计不含编码信号肽序列的克隆引物,上游引物TRagl2-F:5′-GACGAATTCGCAAGTCCCATCTCC-GTGTCTG-3′,下游引物TRagl2-R:5′-ATATCTAGACATGTCTGCTTCTCCA-3′。上下游引物分别含有EcoRⅠ和XbaⅠ酶切位点(以下划线标示)。以里氏木霉cDNA为模板,用高保真PfuDNA聚合酶进行PCR扩增,得到agl2基因片段。
1.2.2 重组表达质粒构建 分别将载体pPIC9K和agl2基因的PCR扩增产物用EcoRⅠ和XbaⅠ双酶切,回收后经T4DNA 连接酶 4 ℃过夜连接。取10 μL连接产物于大肠埃希菌Top10感受态中,静置30 min后于42 ℃热激90 s,涂布于含氨苄青霉素的抗性平板进行筛选。生长出的转化子进一步进行原位PCR验证和双酶切鉴定。
1.2.3 重组表达质粒的毕氏酵母转化 构建好的pPIC9K-agl2重组质粒经SacⅠ线性化后纯化回收,取15 μL质粒与100 μL毕氏酵母GS115感受态细胞混合,在1.5 kV、25 μF、200 Ω下电击转化,立即加入1 mL预冷的1 mol/L山梨醇,30 ℃静置培养3 h后涂布于含50 μg/mL G418的YPD平板,30 ℃培养48~72 h至菌落出现。
1.2.4 转化子的α-半乳糖苷酶活性初筛 挑取50个转化子分别培养于装有5 mL BMGY的50 mL离心管中,置于30 ℃、250 r/min的摇床中培养18 h后,每12 h加入终浓度0.6%的甲醇进行诱导。2 d后测定各转化子表达的α-半乳糖苷酶活性。
1.2.5 重组毕氏酵母的鉴定 提取初筛的重组毕氏酵母基因组DNA,以TRagl2-F和TRagl2-R为引物进行PCR扩增,反应条件:94 ℃预变性4 min;95 ℃ 30 s,55 ℃ 30 s,72 ℃ 120 s,共30个循环;72 ℃延伸5 min,16 ℃保温。PCR产物经1%琼脂糖凝胶电泳进行鉴定。
1.2.6 高表达毕氏酵母菌株的摇瓶发酵 将初筛结果中产α-半乳糖苷酶活性最高的表达菌株培养于5 mL BMGY培养液的50 mL离心管中,30 ℃摇床培养18 h后转接至装有50 mL BMGY培养液的250 mL三角瓶中,且调整初始细胞浓度OD600均约为1。30 ℃、250 r/min的摇床中培养18 h后,每24 h加入终浓度0.6%的甲醇进行诱导。每24 h测定一次粗蛋白浓度、细胞浓度及α-半乳糖苷酶活性。
1.2.7 50 L发酵罐高密度发酵 接种重组酵母菌株于YPD培养基中,生长至OD600为2~6,火焰法接种至BSM培养基中,接种量为10%,经过甘油单批培养、甘油分批补料培养和甲醇分批补料培养,诱导168 h左右。具体方法参考文献[6]。
1.2.8 粗蛋白浓度及α-半乳糖苷酶活性测定 ①粗蛋白浓度测定方法:取适量培养液离心后获得的上清液进行稀释,按改良型Bradford法蛋白浓度测定试剂盒法进行测定。②α-半乳糖苷酶活性测定方法[7]:酶活单位定义为在pH 5.5且37 ℃条件下,每分钟内从10 mmol/L对硝基酚-α-D-吡喃半乳糖中降解释放1 μmol对硝基酚所需要的酶量为一个酶活单位U。经浓度为0.05 mol/L的醋酸钠缓冲液(pH 5.5)稀释的酶液和底物对硝基酚-α-D-吡喃半乳糖溶液于37 ℃下振荡10 min预热,然后吸取各溶液0.1 mL,充分混合,于37 ℃恒温振荡10 min,加入0.8 mL 浓度为0.2 mol/L的碳酸钠溶液,振荡混匀,终止酶反应,在400 nm处测定吸光度。空白对照用经过预热的酶液各0.1 mL,然后加入0.8 mL碳酸钠溶液,振荡混匀,再加入0.1 mL预热的底物于37 ℃振荡10 min。
2 结果与分析
2.1 克隆α-半乳糖苷酶基因agl2
以里氏木霉cDNA为模板,利用高保真pfu酶以及克隆引物Tragl2-F和Tragl2-R扩增α-半乳糖苷酶基因,扩增产物经1.2%琼脂糖凝胶电泳分析表明与理论产物(2 162 bp)大小相吻合(见图1)。

图1 agl2基因扩增产物Fig.1 The PCR product of agl2 gene
2.2 构建重组表达质粒
经EcoRⅠ和XbaⅠ双酶切的目的片段连接到pPIC9K表达载体中,转化大肠埃希菌感受态Top10。抗性平板筛选后的单菌落进一步经原位菌落PCR鉴定(图2)和双酶切验证(图3)证实重组表达质粒构建成功(图4)。重组表达质粒送华大基因公司测序,证实目的基因与GenBank数据一致。

图2 原位菌落PCR产物鉴定Fig.2 The determination of transformants by PCR

图3 重组表达载体pPIC9K-agl2双酶切鉴定Fig.3 The determination of pPIC9K-agl2 by enzymatic digestion
2.3 重组毕氏酵母表达α-半乳糖苷酶
重组表达质粒经SacⅠ线性化后,用电激法转化毕氏酵母GS115,转化细胞直接涂布于G418抗性平板筛选。挑取50个单菌落转接至5 mL BMGY培养基中培养,经甲醇诱导48 h后测定上清液中α-半乳糖苷酶活性,从中筛选出3株酶活相对较高的转化子:AGL2-4#(17.72 U/mL)、AGL2-30#(20.21 U/mL)、AGL2-45#(18.64 U/mL)。分别提取AGL2-4#、AGL2-30#、AGL2-45#的毕氏酵母基因组DNA,以TRagl2-F和TRagl2-R为引物进行PCR扩增,PCR产物经1%琼脂糖凝胶电泳分析,结果见图5,表明agl2基因已整合毕氏酵母基因组中。
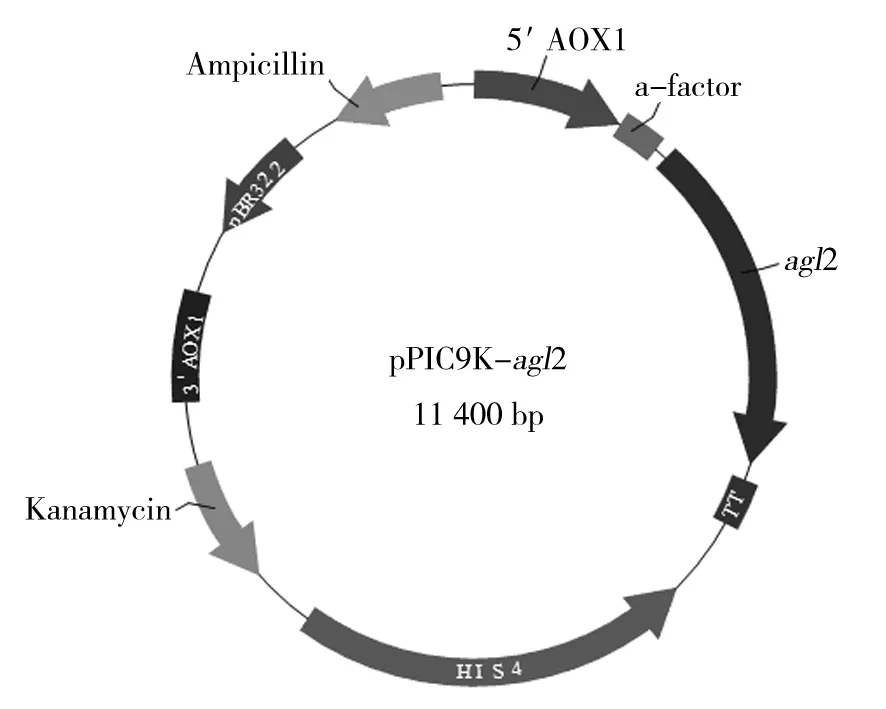
图4 pPIC9K-agl2质粒图谱Fig.4 The map of pPIC9K-agl2

图5 重组毕氏酵母的PCR产物鉴定Fig.5 The PCR products of agl2 in Pichia pastoris
2.4 摇瓶发酵高产α-半乳糖苷酶菌株
250 mL摇瓶发酵重组毕氏酵母AGL2-30#,每24 h测定1次细胞密度、α-半乳糖苷酶活性和上清液中粗蛋白含量(见表1)。AGL2-30#表达菌株诱导96 h后酶活高达103.801 9 U/mL,粗蛋白含量为45.969 1 μg/mL。

表1 外源α-半乳糖苷酶的摇瓶发酵
2.5 50 L高密度发酵
50 L发酵罐发酵经过甘油单批培养、甘油分批补料培养和甲醇分批补料培养3个阶段,发醇168 h后终止。最终发酵液中α-半乳糖苷酶活性达到1 106 U/mL,发酵液中粗蛋白浓度达884 μg/mL。发酵过程的酶活和粗蛋白含量的动态变化见图6。

图6 重组毕氏酵母AGL2-30#在50 L发酵罐高密度发酵Fig.6 Higt density fermentation of recombinant strain AGL2-30#
3 讨 论
α-半乳糖苷酶主要应用于食品[7]、饲料[8]、医药工业[9-10]等领域,应用前景巨大。目前,α-半乳糖苷酶工业化产品主要通过动、植物提取以及筛选微生物菌株发酵获得,传统方法普遍存在酶活低、热稳定性能差、提取工艺复杂、生产效率低等弊端。蔡国林等[11]从黄酒麦曲中筛选到1株黑曲霉wx-07,通过优化发酵条件提高产酶能力,培养96 h时α-半乳糖苷酶活力达到2.4 U/mL。刘彩琴等[12]从发酵豆制品中筛选到1株高产α-半乳糖苷酶的曲霉菌株,96 h培养酶活达22.27 U/mL。王春林等[13]研究青霉产α-半乳糖苷酶的固态发酵生产条件,经过优化培养基使最终发酵酶活达到170 U/g,是目前最适合固体发酵,产酶较高的菌种之一。而通过现代生物技术的基因工程技术和蛋白质技术,已实现α-半乳糖苷酶在真菌(酵母、曲霉)中的异源表达,从而极大提高了α-半乳糖苷酶的生产力,为α-半乳糖苷酶的大规模产业化生产提供坚实基础[14]。高新等[15]通过基因工程技术,克隆到α-半乳糖苷酶基因且实现在毕氏酵母表达,在5 L发酵罐中表达,其α-半乳糖苷酶活性为36 IU/mL。本实验室从青霉克隆到表达α-半乳糖苷酶的新基因,并实现在毕氏酵母中异源表达,在50 L发酵罐高密度发酵,168 h时最高表达酶活力达600 U/mL以上。通过进一步的基因技术改造,酶活力提升1.5倍。本研究从里氏木霉中克隆agl2基因,将agl2基因构建到具有强启动子的pPIC9K表达载体中,并实现在毕氏酵母GS115中异源表达。通过50 L发酵罐高密度发酵,168 h时最高酶活达1 106 U/mL,已具备工业化生产的要求。结合蛋白质工程技术、菌种优化改良和深入摸索发酵条件等生物技术手段,仍具备进一步提升酵母发酵产酶能力,提高酶活性能,以降低生产成本,尽早实现α-半乳糖苷酶产业化,满足市场需求。
影响外源基因在毕氏酵母中表达的因素很多,主要包括:宿主菌的甲醇利用表型、分泌信号、表达基因自身特性、基因剂量、表达框的染色体整合位点和方式、产物稳定性和翻译后修饰等[16]。由于外源蛋白质的复杂性和多样性,外源基因与酵母基因组重组情况及表达调控的途径还不清楚,来源不同的α-半乳糖苷酶基因在同一宿主中的表达受多种因素共同作用的影响,其表达量因此存在很大差别。
[1] 张树政.酶制剂工业[M]. 北京:科学出版社, 1998:780-781.
[2] 章扬培,杨军,高新.新基因重组α-半乳糖苷酶进行B-O血型改造[J]. 科学通报, 2003, 48(1):43-47.
[3] Melissa O, Davis D,Jane H. Cloning, expression and characterization of a blood group B active recombinant α-galactosidase from soybean[J]. Biochemistry Molecular Biology Inter, 1996, 39(3):471-485.
[4] Post DA,Luebke VE. Purification, cloning and properties of alpha-galactosidase fromSaccharopolysporaerythraeaand its use as a reporter system[J]. Applied Microbiology and Biotechnology, 2005, 67(1):91-96.
[5] 丁贤, 戴志明, 殷波. 抑制植物性饲料中抗营养因子的方法[J]. 中国饲料, 2001, (10):9-10.
[6] 李飞, 喻晓蔚, 沙冲, 等. 基因拷贝数和甲醇浓度对重组毕氏酵母产华根霉脂肪酶的影响[J]. 微生物学通报, 2011, 38(3): 301-309.
[7] 潘宝海, 李德发, 陆文清, 等. 对硝基酚-α-D-吡喃半乳糖法测定饲用α-半乳糖苷酶(黑曲霉)活力的方法[J]. 中国农业大学学报, 2002, 7(5): 107-111.
[8] 李孝辉, 竺莉红, 吴吉安, 等. 青霉α-半乳糖苷酶的纯化及酶学特征的研究[J]. 浙江农业学报, 2003,15(2): 99-102.
[9] 肖敏, 刘树峰. 短双歧杆菌α-D-半乳糖苷酶的纯化及性质[J]. 中华微生物学和免疫学杂志, 2001, 21(3): 307-311.
[10]李兴德. 转化血型用蕃茄α-D-半乳糖苷酶的分离、纯化及理化性质[J]. 应用与环境生物学报, 1996, 2(4): 369-375.
[11]蔡国林, 杨旭, 曹钰, 等. 黑曲霉产α-半乳糖苷酶发酵条件研究[J]. 中国酿造, 2007, 170(5): 11-14.
[12]刘彩琴, 何国庆, 陈启和, 等. 一株产α-半乳糖苷酶菌的分离与选育[J]. 食品与发酵工业, 2006, 32(11): 32-35.
[13]王春林, 李淑兰, 陆文清, 等. 饲用青霉α-半乳糖苷酶固态发酵底物的优化[J]. 中国农业大学学报, 2008, 13(1): 51-54.
[14]杨冠东, 刘芳, 李荷. α-半乳糖苷酶的研究进展概况[J]. 现代食品科技, 2006,3(22): 275-279.
[15]高新, 杨军, 李素波, 等. 基因工程α-半乳糖苷酶的制备及其性质研究[J]. 生物工程学报, 2003,19(2): 223-226.
[16]张丞斌, 傅正伟. 影响外源蛋白在巴斯德毕赤酵母中表达和分泌的研究进展[J]. 现代生物医学进展, 2008, 8(7):1382-1384.
High Expression of Trichoderma reesei α-Galactosidase gene agl2 in Pichia pastoris
ZHONG Kai-xin, CHEN Li-zhi, LI Yang-yuan
(GuangdongVTRBio-techCo.,LTD.,Zhuhai519060)
α-galactosidase gene (agl2), amplified by reverse transcription PCR from genome ofTrichodermareeseiwas cloned into plasmid pPIC9K which had a strong promoter AOX1 and transferred and expressed inPichiapastorisGS115 after linearization. The results showed thatagl2 gene was highly expressed inP.pastoris. The α-galactosidase activity reached at the highest value of 1 106 U/mL methanol induction in 50 L high-density fermentation for 168 h. Theagl2 gene had integrated into the genome ofP.pastorisGS115 proved by PCR.
α-galactosidase;Trichodermareesei;Pichiapastoris; high density fermentation
珠海市科技计划项目(2011A0201011002)
钟开新 男,硕士。研究方向为基因工程。E-mail: nhappy_z@163.com
* 通讯作者。男,博士。研究方向为基因工程与发酵工程。E-mail: liyangyuanvtr@163.com
2014-07-17;
2014-10-26
Q936
A
1005-7021(2015)04-0024-05
10.3969/j.issn.1005-7021.2015.04.005
猜你喜欢
中草药(2022年20期)2022-11-15
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02
食品与机械(2019年1期)2019-03-30
食品与机械(2019年1期)2019-03-30
中成药(2018年5期)2018-06-06
中成药(2017年8期)2017-11-22
天然产物研究与开发(2014年3期)2014-04-27
中国酿造(2014年9期)2014-03-11
