
不同强度言语声诱发听性脑干反应及听处理偏侧化分析△
2016-04-13 05:11符秋养梁勇邹岸王涛
听力学及言语疾病杂志 2016年2期
关键词:强度
符秋养 梁勇 邹岸 王涛
·临床研究·
不同强度言语声诱发听性脑干反应及听处理偏侧化分析△
符秋养1梁勇2邹岸3王涛3
【摘要】目的探讨不同强度言语声诱发听性脑干反应(speech evoked auditory brainstem response,s-ABR)的时域和频域变化以及脑干水平是否存在听处理偏侧化现象。方法分别以40、60和80 dB HL言语声对29例健康青年行双耳s-ABR检测,分析双耳时域参数中的主波潜伏期、幅值以及频域参数中的基频(F0)、共振峰(F1~F5)。结果三种刺激强度下,左右耳各主波潜伏期及幅值之间差异均无统计学意义(P>0.05);随言语刺激声强增大,计算强度每降低20 dB时s-ABR各主波潜伏期延长平均值,瞬态性主波V、A、C、O波及周期性主波D、E、F波的潜伏期均显著缩短(P<0.05),幅值显著增加(P<0.05);周期性主波潜伏期延长更明显(P<0.05)。s-ABR频域分析显示,从F0及F1~F5幅值依次降低,与强度变化一致;双耳记录的s-ABR具有很高的相似度,其时域参数和频域参数耳别间差异无统计学意义(P>0.05)。结论s-ABR较好编码了言语的时域和频域信息,且时域和频域参数高度对应并具有相同的强度响应性;瞬态性和周期性成分的s-ABR各主波不同潜伏期特性可能提示更多编码信息;言语在听性脑干水平的处理并无明显偏侧化现象。
【关键词】言语声;强度;听性脑干反应;偏侧化
言语作为人类社会中最重要的复杂声,是声音和意义的集合体,常被称为语音,携带着非常丰富的声学信息[1],频率和强度是其中最重要的特征信息。目前关于言语频率和强度的听觉处理机制仍知之甚少,对于言语认知行为的诸多细节亦不清楚。
言语认知行为的完成除依赖于听觉系统将言语声信息转换为神经电信号并连续加工编码外,还需要经过复杂的高级心理活动。在这一过程中大脑半球表现出了一侧偏向现象,称为听处理偏侧化[2,3]。目前皮层水平的听处理偏侧化研究较为深入,发现主要由一侧大脑半球控制着一定的言语功能,瞬态性的词义性信息分析主要在左半球听区,而周期性的非词义性信息处理可能在右半球[2]。有学者认为耳蜗水平可能也具有类似大脑半球的不对称性处理,其短声刺激的瞬态耳声发射信噪比右耳显著高于左耳,而纯音刺激则左耳显著高于右耳[3]。
脑干作为听觉皮层下中枢,接受双侧耳蜗的传入信息后经过处理整合如实表达了刺激声的信息,保证了高级中枢更复杂、精细的信息处理,是听处理的重要环节。因此,了解言语频率和强度信息在脑干的表达对了解听觉处理过程及了解言语认知行为的神经电生理机制有重要意义。近年研究发现言语声诱发听性脑干反应(speech evoked auditory brainstem response,s-ABR)与刺激声信息有高度的对应关系,在言语认知机制研究及相关临床应用方面引起关注[2,4~8]。s-ABR最常用的刺激声是合成辅-元音单音节/da/[7],/da/包含清辅音/d/和元音/a/两部分,/a/部分时域上有3个周期性波:d(15.27 ms)、e(23.88 ms) 和f(32.38 ms),周期约为9 ms;频域含有基频(F0)及5个共振峰(F1~F5),频率范围分别是:F0= 103~121 Hz,F1= 220~720 Hz, F2= 1700~1240 Hz,F3=2 580~2 500 Hz,F4,5=3 600~4 500 Hz;/da/诱发s-ABR为一系列成分波,由起始反应(onset response, OR)及频率跟随反应(frequency following response, FFR)组成[4~8]。有学者[5]进一步将其分为OR、过渡部分、FFR和终止部分;OR包含V波和A波,过渡部分包含C波,FFR包含D波、E波和F波,终止部分则包含O波。s-ABR的OR、过渡部分以及终止部分属于瞬态性成分,而FFR属于周期性成分,这些反应成分在不同人群中表现出不同的电生理特点,为言语认知机制的基础及临床研究提供了良好的工具[4~7]。
本研究通过分析s-ABR随刺激声强度的变化和耳侧关系,探讨脑干水平言语编码机制及其听处理偏侧化现象,希望有助于进一步认识言语认知行为的神经电生理机制。
1资料与方法
1.1研究对象健康成人29例58耳,男15例,女14例,年龄24~39岁,平均33.19±5.18岁。纳入标准:右利手,具有高中以上教育背景,能流利进行普通话交流,无听觉系统、神经系统疾病及精神障碍病史,500~8 000 Hz纯音平均听阈≤20 dB HL,click-ABR检测结果正常。
1.2s-ABR检测方法测试仪器为美国Intelligent Hearing Systems公司生产的Smart EP诱发电位仪,测试在声电屏蔽室内(GB/T16403)进行。测试前告知受试者闭目,保持安静,尽可能避免眼睑抖动,忽视测试声。测试时受试者平卧位,参考电极置于两侧耳垂,记录电极置于前额正中发际处,地极置于眉心,极间电阻均≤3 kΩ;插入式耳机分别固定于两侧外耳道口深部0.5 cm处;采样率8 000 Hz,带通滤波100~3 000 Hz,伪迹拒绝阈值35 μV,采用同侧给声、同侧记录模式;刺激声采用合成辅-元音单音节/da/[7](图1),刺激率为11.1次/秒,分别以40、60和80 dB HL强度测试,每个受试者每个刺激强度均记录三组数据,每组数据连续记录64 ms,三组数据做平均处理提高信噪比。测试时刺激强度的给声顺序以及刺激耳别随机决定,调整强度及耳别时受试者休息2分钟,每组数据测试间隔1分钟。
1.3s-ABR的时域(主波潜伏期、幅值)和频域(基频、共振峰)分析
1.3.1s-ABR时域分析s-ABR主波判定采用软件结合人工方法,首先由研究者在Matlab平台上开发的软件模块进行初步判定,然后由不了解反应参数的s-ABR研究者对软件判定主波进行确定或调整。软件模块主要功能包括读取Smart EP记录的3组原始数据,做平均叠加去噪处理得到一个平均波形,然后,根据设定的主波判定准则,对平均波形进行主波判定,并在人工确定后读取出主波潜伏期和幅值。综合听脑干反应特性、/da/声学参数及同类研究结果[1,4,5,7],软件模块中平均波形的主波判定准则为:潜伏期≤10 ms的幅值最大正性波峰判为V波,紧接的负性波峰判为A波,10 ms<潜伏期≤20 ms的幅值最大负性波峰判为C波,20 ms<潜伏期≤30 ms的幅值最大负性波峰判为D波,30 ms<潜伏期≤40 ms的幅值最大负性波峰判为E波,40 ms<潜伏期≤50 ms的幅值最大负性波峰判为F波;50 ms<潜伏期≤55 ms的幅值最大负性波峰判为O波。
1.3.2s-ABR频域分析对58耳3种强度的s-ABR平均波形进行快速傅立叶变换,提取/da/基频及5个共振峰信息并计算其幅值(对应/da/基频和共振峰频率范围的平均峰值),即F0及F1~F5的幅值。
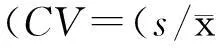
2结果
2.1s-ABR的波形29例受试者双耳s-ABR时域及频域图形与刺激声具有很高相似性,反应时域波形由一系列成分波组成,其主波包含正波V、负波A、负波C、负波D、负波E、负波F和负波O(图1),频域图中FFR与元音/a/具有非常准确的对应关系,而辅音/d /没有很好地被表达(图2)。
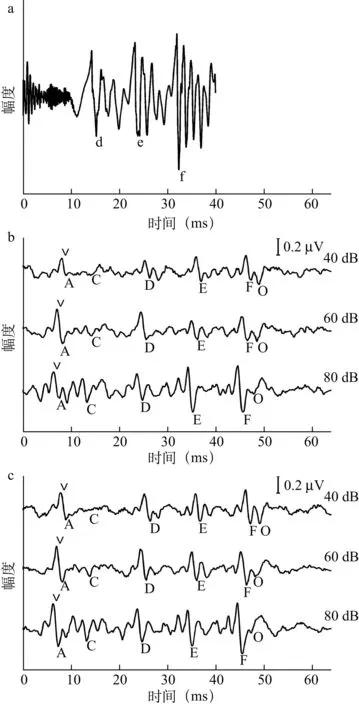
图1 /da/的时域波形及双耳s-ABR的时域平均波形
a为/da/时域波形,b、c分别为左右耳s-ABR时域平均波形
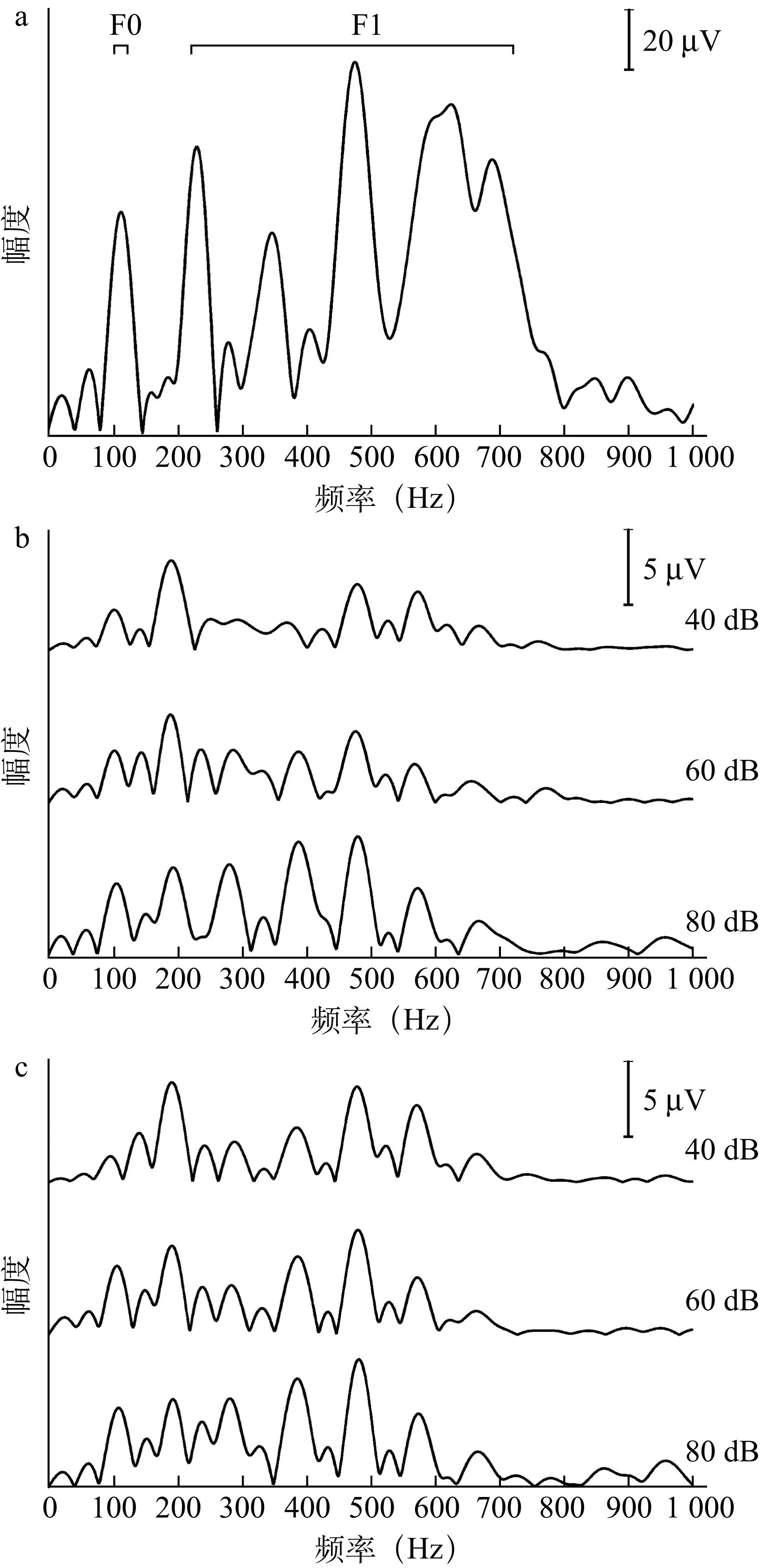
图2 /a/的频域波形及双耳FFR的频域平均波形
a为/a/频域波形,b、c分别为左右耳FFR频域平均波形
2.2不同刺激强度下双耳s-ABR各主波潜伏期及幅值由表1~4可见,三种刺激强度下,双耳间各主波潜伏期及幅值的差异无统计学意义(P>0.05),说明/da/声诱发的s-ABR无偏侧化现象;不同强度下各主波潜伏期差异有统计学意义(P< 0.05),多重比较显示40与60 dB HL、60与80 dB HL、40与80 dB HL强度下,各波潜伏期差异均有统计学意义(P<0.05);不同强度下各主波幅值(除D波外)差异有统计学意义(P<0.05),多重比较,仅在40与80 dB HL强度下部分主波幅值差异有统计学意义(P<0.05);潜伏期CV=1%~14%,幅值CV=31%~83%。计算强度每降低20 dB时s-ABR各主波潜伏期延长平均值[(40 dB强度下的潜伏期值-80 dB强度下的潜伏期值)/2],各瞬态性主波V、A、C及O波潜伏期变化均值分别为0.79±0.32、0.69±0.28、0.72±0.35及0.68±0.40 ms;而各周期性成分D、E和F波潜伏期变化均值分别为1.21±0.59、1.14±0.61和1.17±0.67 ms,可见,周期性主波潜伏期延长更明显(P<0.05)。

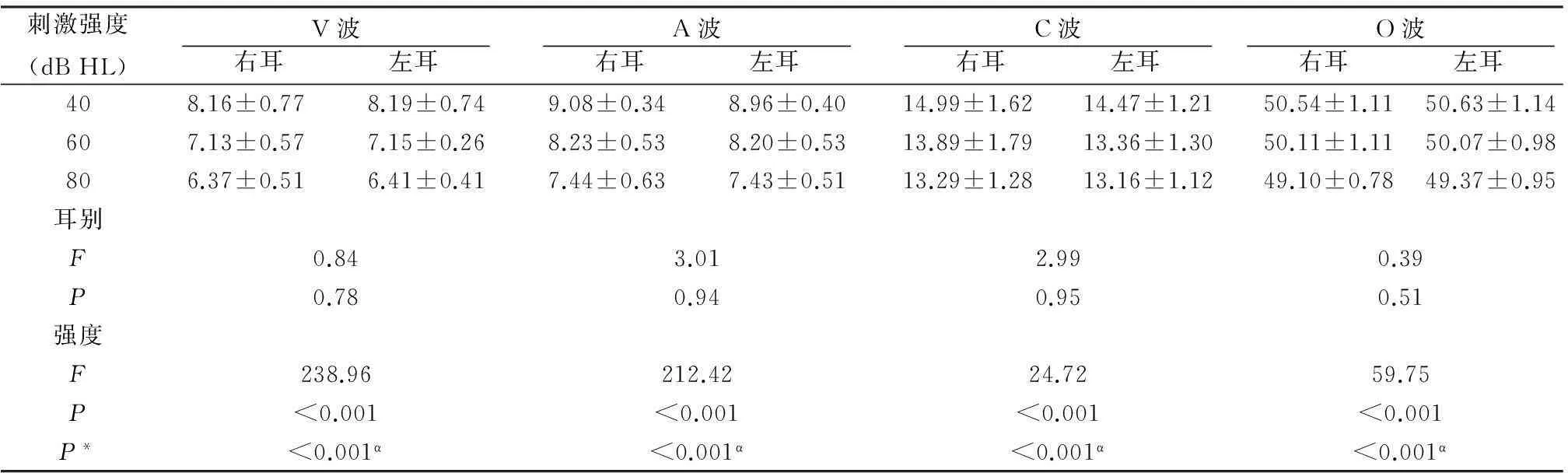
刺激强度(dBHL)V波 右耳 左耳 A波 右耳 左耳 C波 右耳 左耳 O波 右耳 左耳 408.16±0.778.19±0.749.08±0.348.96±0.4014.99±1.6214.47±1.2150.54±1.1150.63±1.14607.13±0.577.15±0.268.23±0.538.20±0.5313.89±1.7913.36±1.3050.11±1.1150.07±0.98806.37±0.516.41±0.417.44±0.637.43±0.5113.29±1.2813.16±1.1249.10±0.7849.37±0.95耳别F0.843.012.990.39P0.780.940.950.51强度F238.96212.4224.7259.75P<0.001<0.001<0.001<0.001P*<0.001α<0.001α<0.001α<0.001α
注:P*为各强度多重比较,α为所有两两比较

±s)
注:P*为各强度多重比较,α为所有两两比较

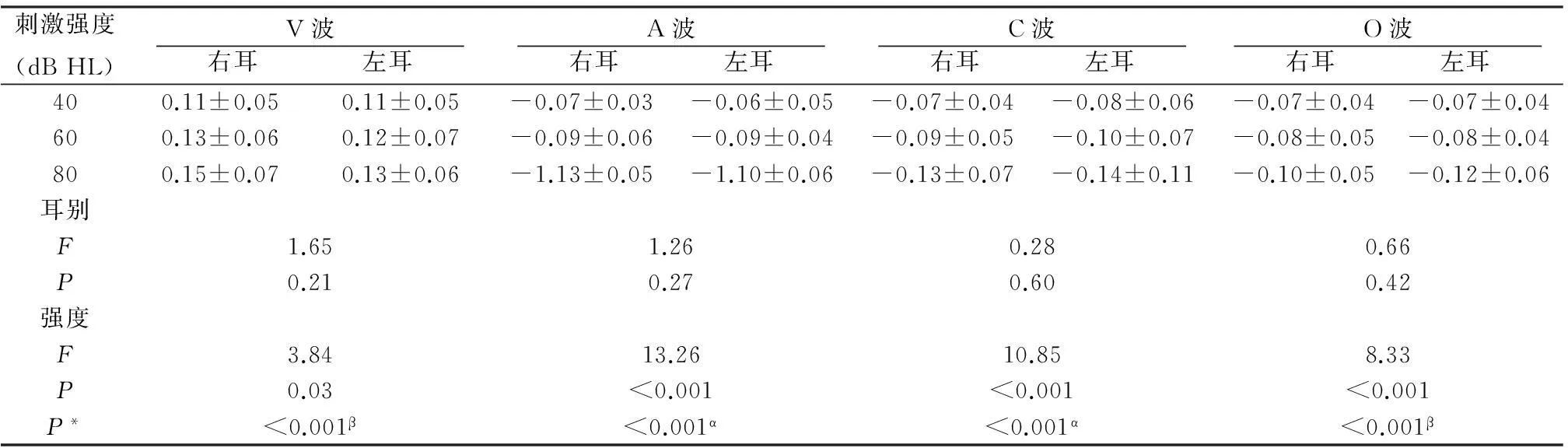
刺激强度(dBHL)V波 右耳 左耳 A波 右耳 左耳 C波 右耳 左耳 O波 右耳 左耳 400.11±0.050.11±0.05-0.07±0.03-0.06±0.05-0.07±0.04-0.08±0.06-0.07±0.04-0.07±0.04600.13±0.060.12±0.07-0.09±0.06-0.09±0.04-0.09±0.05-0.10±0.07-0.08±0.05-0.08±0.04800.15±0.070.13±0.06-1.13±0.05-1.10±0.06-0.13±0.07-0.14±0.11-0.10±0.05-0.12±0.06耳别F1.651.260.280.66P0.210.270.600.42强度F3.8413.2610.858.33P0.03<0.001<0.001<0.001P*<0.001β<0.001α<0.001α<0.001β
注:P*为各强度多重比较,α为所有两两比较,β为仅40与80 dB HL强度比较


刺激强度(dBHL)D波 右耳 左耳 E波 右耳 左耳 F波 右耳 左耳 40-0.12±0.06-0.11±0.05-0.12±0.05-0.12±0.05-0.12±0.05-0.11±0.0460-0.14±0.05-0.13±0.06-0.13±0.05-0.13±0.04-0.14±0.06-0.13±0.0480-0.13±0.05-0.13±0.05-0.14±0.05-0.15±0.06-0.15±0.06-0.16±0.05耳别F1.660.090.55P0.210.770.50强度F1.693.288.46P0.190.04<0.001P*0.03β<0.001β
注:P*为各强度多重比较,α为所有两两比较,β为仅40与80 dB HL强度比较
2.3s-ABR频域分析结果/da/基频(F0)及共振峰(F1~F5)见表5,表达幅值从F0到F1~F5依次降低(F4~5数值接近0,未列出),幅值变化与言语强度改变一致。
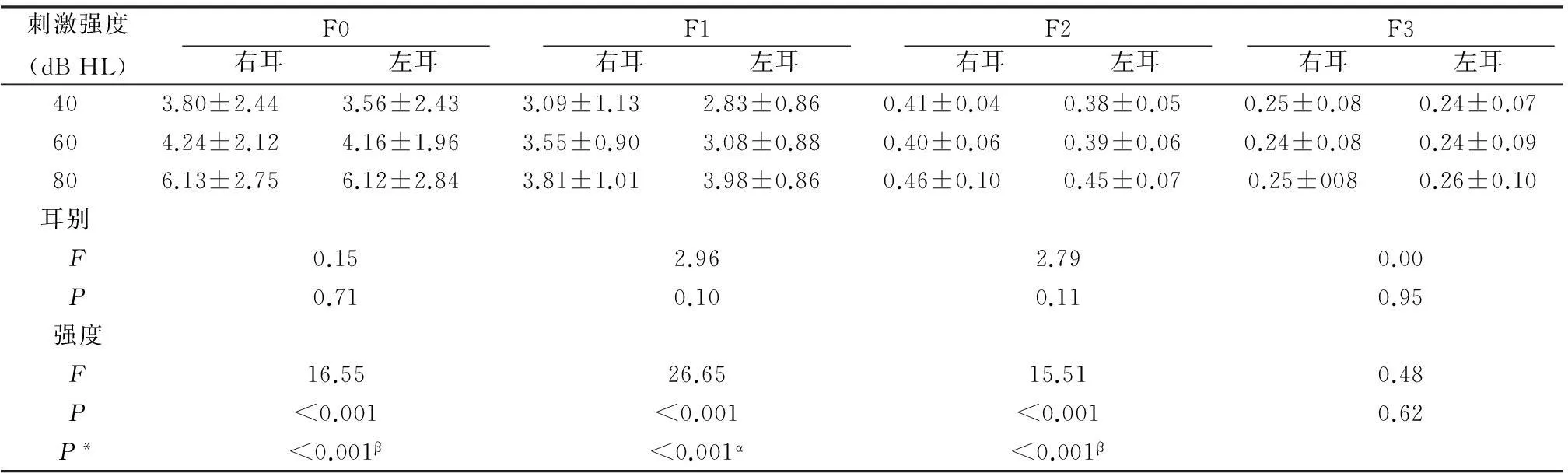
刺激强度(dBHL)F0 右耳 左耳 F1 右耳 左耳 F2 右耳 左耳 F3 右耳 左耳 403.80±2.443.56±2.433.09±1.132.83±0.860.41±0.040.38±0.050.25±0.080.24±0.07604.24±2.124.16±1.963.55±0.903.08±0.880.40±0.060.39±0.060.24±0.080.24±0.09806.13±2.756.12±2.843.81±1.013.98±0.860.46±0.100.45±0.070.25±0080.26±0.10耳别F0.152.962.790.00P0.710.100.110.95强度F16.5526.6515.510.48P<0.001<0.001<0.0010.62P*<0.001β<0.001α<0.001β
注:P*为各强度多重比较,α为所有两两比较,β为仅40与80 dB HL比较
3讨论
3.1s-ABR时域特性分析听脑干是听皮层下接受、分析和发送听觉信息的多功能整合中心,其内部众多纤维联系形成多种反馈回路,对听觉信息的提取和调制有着重要作用[6,9,10]。s-ABR的产生与言语信号特征密切相关,反应中的V波及A波为言语信号开始诱发,C波为言语信号中元音开始诱发,O波则为言语信号结束所诱发[7],这些瞬态性波可能是脑干核团中章鱼细胞(ocopus cells)的活动结果,这类听神经元具有强度响应性,并对刺激中的瞬态性信息产生爆发式反应[8]。而周期性波D、E、和F则是言语信号中元音信息的跟随反应,可能来源于脑干核团中对1 kHz以下频率表现出显著锁相特性的神经元[11],如“给声”反应细胞、“斩波”细胞、初始样细胞等。本研究发现,随着言语强度增大,s-ABR瞬态性波及周期性波均表现出了潜伏期显著缩短以及幅值显著增大,这可能与强度变化影响了这些脑干神经元首个动作电位的发放时间(first spike latency,FSL)和发放数(first spike count,FSC)有关,一般认为强度增大可使听神经元FSC增加,产生较大反应幅值[12,13]。然而,听觉诱发电位幅值稳定性差,本研究中s-ABR各强度下幅值变异系数为31%~83%,波动性较大,这可能与听神经元FSC的差异和不稳定有关。研究发现听觉神经元存在3种FSC模式,一种随着强度增加而持续增加,一种随强度增加到一定水平后趋于稳定,另一种则随强度增加到一定水平后反而下降[12];这3种FSC模式的神经元广泛存在于听觉系统各级核团[13],目前尚不清楚这些神经元在不同核团的分布情况。此外,同一听神经元FSC也不稳定,相同强度刺激引起的发放数变化很大[13]。从本研究结果看,强度变化引起s-ABR幅值改变的数值较小,可能是因为20 dB和40 dB的强度差距尚不能引起听神经元总体FSC明显增加;研究发现强度刺激引起的听神经元总体FSC在60 dB强度差内变化不大[14],这与本文结果符合。有学者认为听神经元FSL是声音强度编码更可靠的机制,随着强度增大表现为逐渐减小[12,15~17];听神经元FSL具有高度稳定性,同一刺激FSL标准差常在零点几毫秒之内,在较高声强时更为明显[18];此外,相同频率的FSL-强度曲线可准确反映两者关系,而不同频率的FSL-声强曲线经时间和强度等效后也可以完全重叠,不受刺激时程的影响[13],因而听觉诱发电位的反应潜伏期稳定性非常高,并且随刺激强度增大而逐渐减小。本研究结果显示s-ABR各强度下潜伏期变异系数较小(1%~14%),这与听神经元FSL的稳定性符合。尽管FSC和FSL可能各自解释了声音强度的编码机制,但FSL可达数毫秒甚至微秒的精确度携带着更多信息[15],因此,听觉电位反应潜伏期具有更高的临床应用价值。
从文中结果看,当言语刺激强度以相同程度增加时,s-ABR瞬态性主波之间的潜伏期变化值接近,而周期性成分潜伏期的变化值也接近,但周期性主波潜伏期延长更明显,提示s-ABR瞬态性成分可能具有相同的神经元来源,而周期性主波的神经元也可能具有同源性,再次证实了学者关于s-ABR神经起源的观点[6,7]。然而尽管s-ABR瞬态性成分和周期性主波的来源神经元都位于同一听觉通路,但彼此功能不同,可能构成了2条平行传导途径[6,7,9,10]。有学者提出听脑干信号处理具有特征提取和平行处理原则,声信息的某种特征被一类特定听神经元提取和识别后由许多并列的信息传递通路进行平行处理[6,9],保证了信息被如实表达到高级中枢[10]。what-where通路可能是听脑干中平行神经通路之一,where通路主要处理词义信息,而what通路主要负责非词义信息[6,7,9]。研究发现有些儿童对言语的词义识别能力受损,表现出不同程度的诵读及听写困难;但对言语的非词义信息识别不受影响,如对说话者身份及情感等方面识别能力正常[2,4,7];这类儿童的s-ABR测试表现出周期性成分无改变,但瞬态性主波明显异常[2],在噪声下更显著[7];提示s-ABR瞬态性主波来源神经元可能构成了where通路,周期性成分神经元则可能组成了what通路。s-ABR瞬态性主波和周期性成分不同的生理特性可能提供更多解密言语认知行为的线索,有待进一步研究。
3.2s-ABR频域特性分析本研究结果显示s-ABR反应和刺激声在时域上相似度非常高;频域中FFR与元音/a/也具有非常准确的对应关系,但辅音/d/似乎没有很好地被表征,与言语声/ba/诱发脑干反应的研究结果[8]符合。FFR对/a/的精确表达提示元音信息在脑干水平的编码可能已经达到了很高程度,辅音信息未“非编码”则可能因为辅音识别需要借助语境[19],然而语境信息是否被表达在s-ABR的瞬态性成分中尚有待进一步证实。声波传入耳蜗后在基底膜传播,不同频率在基底膜不同部位发生最大振动,引起该处内毛细胞产生动作电位,从而将机械振动转变成电信号传入中枢;在这一过程中,每个部位的内毛细胞响应一个最佳频率,形成了一对一的传导通道,并且在基底膜波动周期的特定相位发放冲动,因而使频率及其时间特征以锁相发放形式被传输[10]。听觉通路各级中枢均存在与此对应的频率拓扑结构,不同频率响应特性的听神经元按其特征频率由高到低或者由低到高排列[9,10];这种频率拓扑结构使得声信号不同频率信息可以通过不同通道传导,很可能是平行神经通路的基础。基频是声调识别的主要特征,共振峰是言语辨别的重要特征,对元音尤为重要,文中结果显示,/da/基频F0和5个共振峰F1~F5编码幅度从基频到各个共振峰依次降低,幅度变化与言语强度改变一致,提示s-ABR频域编码幅值变化可能也表达了言语的强度变化信息;有学者提出s-ABR忠实编码了言语信息[1,2,6,7],本研究结果与此相符。
3.3s-ABR无偏侧化现象分析听觉系统各个水平的听处理偏侧化是听觉研究热点。听觉系统为双侧性的神经通路,由于对侧听投射更具优势,起始耳的对侧半球常得到更强的激活[20],因而出现了耳侧优势,在加工数字、词汇等言语材料时表现为右耳优势,而加工音乐、 环境声等非言语材料时表现为左耳优势[2,21]。此外,在对辅音处理中也表现出右耳优势,元音处理则无明显偏侧化现象,原因可能是辅音信息变化更快速,因而右耳-左半球交叉优势得到更多激活,但这种左半球优势并非绝对[2]。研究发现声调加工也存在右耳-左半球优势,在声调语种人群中更为显著,而非声调语种人群则没有显著耳侧优势或偏向右半球[22]。目前研究认为听处理偏侧化是解剖不对称性、左半球言语优势以及右半球非言语优势共同所致[2],右耳言语刺激时,听信号可直接投射到左半球进行处理;而左耳声信号需先投射到右半球,并经过胼胝体中继站处理后方转入左半球处理中心;同样,左耳非言语信号可直接投射到右半球进行处理,而右耳则经过左半球及胼胝处理后才能转入右半球处理中心[20,23]。从文中结果看,不同刺激强度下左、右耳的s-ABR无显著差异,/da/时域信息或频域信息的听脑干编码未表现出耳侧优势,而辅音、元音、声调以及词义信息在脑干水平的加工亦无偏侧化现象,其原因可能是听脑干信息处理功能性通路是相互平行的,what-where通路可能平行分布于解剖结构中的对侧和同侧通路;不同耳侧及不同听信息经过听脑干平行处理后投射到对应听皮层,然后在半球优势作用下才表现出耳侧优势;但听脑干平行处理和听皮层偏侧优势在言语认知行为中的相互关系尚有待进一步研究。
4参考文献
1Akhoun I, Moulin A, Jeanvoine A, et al. Speech auditory brainstem response (speech ABR) characteristics depending on recording conditions and hearing status: an experimental parametric study[J]. Journal of Neuroscience Methods,2008, 175:196.
2Vanvooren S, Poelmans H, Hofmann M, et al. Hemispheric asymmetry in auditory processing of speech envelope modulations in prereading children[J]. The Journal of Neuroscience,2014, 34: 1523.
3Sininger YS, Cone-Wesson B. Asymmetric cochlear processing mimics hemispheric specialization[J]. Science,2004, 305:1581.
4Erika S, Nina K. Musical training heightens auditory brainstem function during sensitive periods in development[J]. Frontiers in Psychology,2013, 4: 622.
5符秋养,梁勇,苏园园,等.健康青年人言语诱发听性脑干反应的成分及特性[J].中华耳鼻咽喉头颈外科杂志,2009,44:900.
6Kraus N, Nicol TG. Brainstem origins for cortical "what" and "where" path ways in the auditory system[J]. Trends in Neurosciences,2005, 28: 176.
7Johnson KL, Nicol TG, Kraus N. Brain stem response to speech: a biological marker of auditory processing[J]. Ear and Hearing,2005, 26: 424.
8Akhoun I, Galle′go S, Moulin A, et al. The temporal relationship between speech auditory brainstem responses and the acoustic pattern of the phoneme /ba/ in normal-hearing adults[J]. Clinical Neurophysiology,2008, 119: 922.
9Rauschecker JP, Biao T. Mechanisms and streams for processing of "what"and "where"in auditory cortex[J]. Proceeding of the National Academy of Sciences of the United States of America,2000,97: 11800.
10Rauschecker JP, Shannon RV. Sending sound to the brain[J]. Science,2002, 295: 1025.
11Wang J, Qin L, Chimoto S, et al. Response characteristics of primary auditory cortex neurons underlying perceptual asymmetry of ramped and damped sounds[J]. Neuroscience,2014, 256: 309.
12Phillips DP, Sark SA. Separate mechanisms control spike numbers and inter-spike intervals in transient responses of cat auditory cortex neurons[J]. Hearing Research,1991, 53: 17.
13Tan X, Wang X, Yang W, et al. First spike latency and spike count as functions of tone amplitude and frequency in the inferior colliculus of mice[J]. Hearing Research,2008, 235: 90.
14Kitzes L. Binaural interactions shape binaural response structures and frequency response functions in primary auditory cortex[J]. Hearing Research,2008, 238: 68.
15Heil P. First spike latency of auditory neurons revisited[J]. Current Opinion in Neurobiology,2004, 14: 461.
16VanRullen R, Guyonneau R, Thorpe SJ. Spike times make sense[J]. Trends in Neurosciences,2005, 28: 1.
17Johansson RS, Birznieks I. First spikes in ensembles of human tactile afferents code complex spatial fingertip events[J]. Nature Neuroscience,2004, 7: 170.
18Wiener MC, Richmond BJ. Decoding spike trains instant by instant using order statistics and the mixture-of-Poissons model[J]. The Journal of Neuroscience,2003, 23:2394.
19Chandrasekaran B, Hornickel J, Skoe E, et al. Context-dependent encoding in the human auditory brainstem relates to hearing speech in noise: Implications for developmental dyslexia[J]. Neuron,2009, 64: 311.
20Hashimoto R, Homae F, Nakajima K, et al. Functional differentiation in the human auditory and language areas revealed by a dichotic listening task[J]. Neuroimage, 2000, 12: 147.
21Itoh K, Miyazaki K, Nakada T. Ear advantage and consonance of dichotic pitch intervals in absolute-pitch possessors[J]. Brain and Cognition, 2003, 53: 464.
22Wang Y, Jongman A, Sereno JA.Dichotic perception of Mandarin tones by Chinese and American listeners[J]. Brain and Language,2001, 78: 332.
23Westerhausen R, Hugdahl K. The corpus callosum in dichotic listening studies of hemispheric asymmetry: a review of clinical and experimental evidence[J]. Neuroscience & Biobehavioral Reviews,2008, 32: 1044.
(2014-09-11收稿)
(本文编辑李翠娥)
Speech-ABR Representations and Non-lateralization Effect for the Stimulation Intensity
Fu Qiuyang*, Liang Yong, Zou An, Wang Tao
(*Department of Otolaryngology, Guangdong NO.2 Provincial People’s Hospital, Guangzhou,510515, China)
【Abstract】ObjectiveTo investigate the effects of stimulation intensity on subcortical auditory processing of speech and assess the subcortical asymmetry of speech elements at the brainstem level using speech evoked auditory brainstem response (s-ABR), so as to provide more clues for the mechanism of speech cognitive behaviors.MethodsSpeech syllables /da/ with three intensities(40 dB HL,60 dB HL and 80 dB HL) were presented monaurally to 29 normal hearing adults, and corresponding s-ABRs were recorded. The stimulus and response parameters, including latency and amplitude in time domain, fundamental frequency and formants in frequency domain were analyzed statistically.ResultsWith the increase of stimulation intensity, significant shorter latency and larger amplitude were observed for the feature peaks of s-ABR (P<0.05). The latency changes corresponding to the same intensity increment were at variance with the periodic components and the transient components. The fundamental frequency and formants of the stimulus syllables that extracted preferably from s-ABRs were encoded less vigorously in ascending order which was the same as that for the intensity. The responses to the right and left monaural stimulation resembled without significant lateralized difference.ConclusionThe s-ABR is able to encode faithfully the speech sound in terms of timing and spectrum components with high correspondence with the stimulus intensity.
The different latency characteristics of periodic components and transient responses may imply more underlying coding information of speech sound. However, lateral asymmetry speech processing is not considerable at the brainstem level.
【Key words】Speech sound;Stimulation intensity;Auditory brainstem response;Lateralization
【中图分类号】R764.04
【文献标识码】A
【文章编号】1006-7299(2016)02-0116-07
DOI:10.3969/j.issn.1006-7299.2016.02.002
作者简介:符秋养,男,广东人,副主任医师,主要研究方向为耳科学及言语听觉科学临床研究。通讯作者:梁勇(Email:yliang@fimmu.com);王涛(Email:taowang@smu.edu.cn)
网络出版时间:2015-3-314:39
网络出版地址:http://www.cnki.net/kcms/detail/42.1391.R.20150303.1439.027.html
△国家自然科学基金项目(61172033)、2014年广东省科技计划立项(411141963067)和广东省医学科研基金(A2015074)联合资助
1广东省第二人民医院耳鼻咽喉-头颈外科(广州510515);2南方医科大学附属南方医院耳鼻咽喉-头颈外科;3南方医科大学生物医学工程学院
猜你喜欢
中国棉花(2021年4期)2021-12-05
建材发展导向(2021年7期)2021-07-16
重型机械(2019年3期)2019-08-27
新高考·高一物理(2016年7期)2017-01-23
新高考·高一物理(2016年7期)2017-01-23
新高考·高一物理(2016年7期)2017-01-23
中学物理·高中(2016年8期)2016-08-08
中学生数理化·高二版(2016年9期)2016-05-14
中国惯性技术学报(2015年1期)2015-12-19
