
真菌诱导子对龙眼胚性愈伤组织生长和多糖积累的影响
2016-07-09 18:39徐小萍赖瑞联林玉玲赖钟雄
热带作物学报 2016年11期
徐小萍 赖瑞联 林玉玲 赖钟雄

摘 要 为探讨真菌诱导子对龙眼胚性愈伤组织生长及多糖含量的影响。以龙眼胚性愈伤组织为材料,采用单因素分析法探究了真菌诱导子种类、诱导时间和浓度对龙眼胚性愈伤组织生长和多糖积累的影响。结果表明,不同真菌诱导子处理下龙眼胚性愈伤组织中多糖含量由高到低排序分别为:龙眼拟茎点霉>胶孢镰刀菌>尖孢镰刀菌;采用100 mg/L浓度的龙眼拟茎点霉诱导子诱导15 d,龙眼胚性愈伤组织多糖含量最高,比对照提高了61.34%,且不同处理间存在显著差异。但真菌诱导子处理下龙眼胚性愈伤组织生长状态较差,且增殖率显著下降。龙眼拟茎点霉作为龙眼病原内生菌,其活性诱导物能促进龙眼胚性愈伤组织中多糖积累,从而一定程度上提高植物的免疫防御能力。研究结果为探究真菌诱导子对龙眼多糖合成和代谢机理的研究奠定基础。
关键词 龙眼;真菌诱导子;多糖;胚性愈伤组织;增殖
中图分类号 S667.2 文献标识码 A
龙眼(Dimocarpus longan Lour.)是中国南方热带亚热带名贵果树。研究表明,龙眼果肉、果皮及枝叶等各组织器官中含有大量多糖[1-3],尤其是龙眼果肉多糖成分具有较高的生物药理活性,其含量高达64.27%,极具开发前景。多糖是生物体中重要初生代谢物质,具有清除自由基、调节免疫系统、抗癌抗菌和抑制酪氨酸酶等生化功能[4-5]。作为一种重要活性成分,多糖还能增强神经功能、治疗神经损伤、降血糖和抗肿瘤作用[6-8]。然而,以果肉等组织器官为原材料提取龙眼多糖易受季节、气候等因素限制,开发一种高效多糖生产技术具有重要的现实意义。
龙眼胚性愈伤组织具有较强的增殖和体细胞胚胎发生能力,为龙眼药用活性成分的研究生产提供了有效平台[9-10]。且大量研究表明,植物愈伤组织在多糖生产中具有很好的应用前景[11-13]。因此,笔者认为研究龙眼胚性愈伤组织中的多糖积累有重要意义。真菌诱导子是来自微生物的生物活性物质,能使植物产生过敏反应,并诱导植物积累特定的次生代谢产物[14],提高植物的防御抗病能力。植物多糖在特定条件下参与某些次生代谢物合成,多糖合成代谢可能受真菌诱导子影响[15-16],探究龙眼的致病内生真菌与非龙眼内生菌分别制备的诱导子对提高龙眼胚性愈伤组织多糖积累的影响有重要意义。研究表明龙眼果肉中可分离获得龙眼拟茎点霉、镰刀菌和炭疽菌,其中龙眼拟茎点霉分离率最高,达44%,其他2种分离率较低[17]。笔者推测真菌诱导子对龙眼胚性愈伤组织多糖诱导具有重要作用。因此,本试验以龙眼胚性愈伤组织为材料,以3种真菌制备的诱导子为处理因子。采用单因素分析方法研究不同真菌诱导子种类、诱导时间和处理浓度对胚性愈伤组织生长和多糖积累的影响,并通过相关性和显著性分析,为真菌诱导子对龙眼多糖合成代谢、信号识别和信号转导等作用機理的研究提供科学依据。
1 材料与方法
1.1 材料
龙眼‘红核子(Dimocarpus longan Loμr. cv.Honghezi)胚性愈伤组织由福建农林大学园艺植物生物工程研究所提供[18]。3种真菌分别是龙眼拟茎点霉(Phomopsis longanae Chi)、尖孢镰刀菌(Fusarium oxysporum)、胶孢镰刀菌(Fusarium subglutinans Wollenw. et Reinking),均由福建农林大学植物保护学院赠送。
1.2 方法
1.2.1 不同种类诱导子处理龙眼胚性愈伤组织
参考Wang等[19]、林艳君等[20]的方法,获得3种真菌诱导子。往龙眼胚性愈伤组织增殖培养基L1分别添加100 mg/L的3种真菌诱导子处理龙眼胚性愈伤组织,暗培养20 d,通过多糖含量测定筛选最适宜的诱导子类型。
1.2.2 不同诱导时间和诱导子浓度处理龙眼胚性愈伤组织 试验进一步采用单因素法分别探究处理15、20、25 d后龙眼胚性愈伤组织多糖积累情况,并设置对照试验;其次在最适宜天数处理试验的基础上,结合前人研究报道[20],分别采用0、50、100、200 mg/L浓度的诱导子进行最佳浓度筛选。每个处理进行3个重复,分别采用苯酚硫酸法进行多糖含量测定[21]、愈伤组织状态观察和增殖系数统计。
1.2.3 标准曲线制作及多糖测定 称取0.1 g蔗糖干样作为标准品,溶解定容至1 000 mL,依次稀释成不同倍数的浓度,以紫外分光光度计在485 nm波长下测定的吸光值为纵坐标,以糖浓度为横坐标,利用吸光值与总糖量成线性关系,制作多糖含量标准曲线如图1,线性回归方程为y=0.018 3x+0.004 6,相关系数R2=0.999 8。
多糖提取测定,采用苯酚硫酸法。称取龙眼胚性愈伤组织干样0.03 g,加1 mL的双蒸水,沸水浴30 min,7 800 r/min常温离心10 min,取上清液后重复该步骤2次,双蒸水漂洗残渣,离心,定容至10 mL;加5 mL石油醚,台式恒温振荡器常温200 r/min,10 min;静置分层,弃上层溶液,下层溶液加2 mL氯仿 ∶ 正丁醇(5 ∶ 1)溶液,混匀至乳浊状,静置分层;取50 μL上层溶液,双蒸水定容至2 mL;加1 mL 9%苯酚溶液,摇匀,加5 mL浓硫酸,摇匀。室温下冷却静置30 min,紫外分光光度计485 nm波长测定吸光值。多糖含量计算公式:多糖含量(%)=X/(M/V1)*V2/V3×100%。其中X:样品中多糖浓度(mg/mL),M:样品质量g,V1:首次样品定容的体积10 mL,V2:稀释时吸取的样品体积50 μL,V3:稀释后定容的体积2 mL,见图1。
1.3 数据统计
采用Excel 2003进行数据统计和图表制作,并采SPSS20.0进行相关分析、方差分析和显著性测验。
2 结果与分析
2.1 真菌诱导子对多糖含量的影响
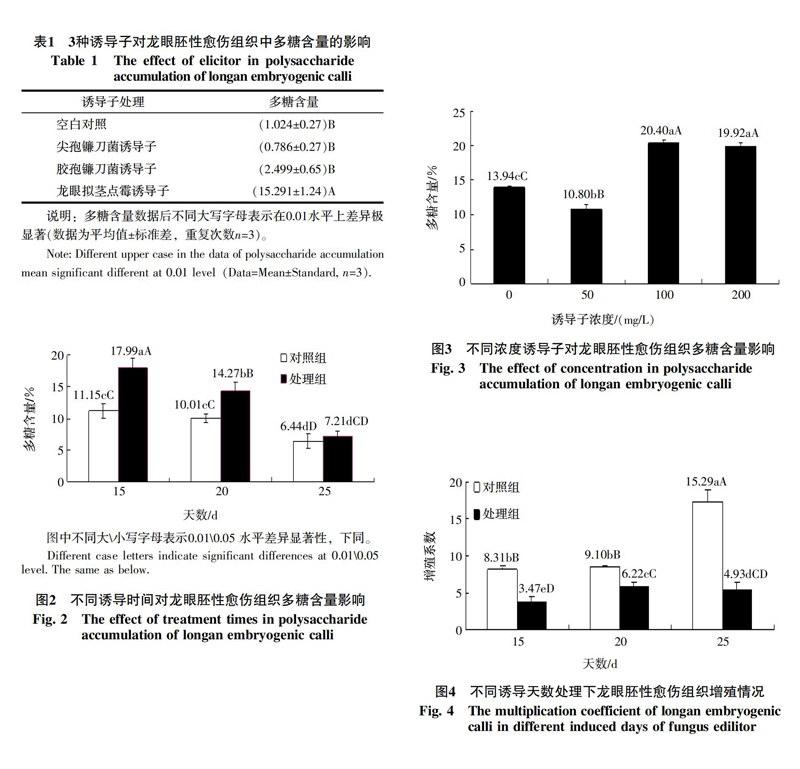
2.1.1 真菌诱导子种类对多糖含量的影响 对所得数据进行方差齐性检验、单因素分析结合Duncan法进行两两比较,如表1,结果表明3种真菌诱导子中龙眼拟茎点霉诱导子对龙眼胚性愈伤组织中多糖含量的影响达到极显著性水平(α=0.01),多糖含量高达15.29%,尖孢镰刀菌诱导子不利于胚性愈伤组织多糖积累,胶孢镰刀菌诱导子对多糖积累诱导效果不显著。因此,选用龙眼拟茎点霉诱导子对龙眼胚性愈伤组织进行不同诱导天数和不同浓度的处理,进一步探究多糖含量的积累情况。
2.1.2 诱导时间对龙眼胚性愈伤组织多糖含量的影响
筛选出最适的诱导子种类后,试验进一步对最适诱导时间进行筛选。结果表明,龙眼拟茎点霉诱导子处理15 d时,最能促进胚性愈伤组织多糖积累,处理25 d时对多糖积累也有促进作用,但积累量最少。经方差齐性检验、单因素方差分析、F检验及LSD多重比较数据分析表明,与对照相比,α=0.05显著性水平下,龙眼拟茎点霉诱导子诱导不同时间对愈伤组织多糖积累具有显著性影响。结合图2分析,龙眼胚性愈伤组织在相同浓度不同诱导天数的龙眼拟茎点霉诱导子处理下,多糖含量随诱导天数增加而减少,其中15 d时的多糖含量达到最高为17.99%,其次是20 d(14.27%),25 d时最低,仅为7.22%;与对照相比,多糖含量分别提高了61.34%、42.56%、12.04%。可见,龙眼胚性愈伤组织多糖积累与诱导时间密切相关,而15 d为最佳处理时间。
2.1.3 龙眼拟茎点霉诱导子浓度对多糖含量的影响
继不同诱导天数处理后,进一步探究诱导子浓度对龙眼胚性愈伤组织多糖含量影响。结果表明100 mg/L龙眼拟茎点霉诱导子最能促进龙眼胚性愈伤组织多糖积累,其次是200 mg/L,多糖含量分别达到20.40%和19.92%,二者无显著差异;而添加浓度为50 mg/L时对多糖积累作用不明显(图3),其他处理间的差异显著性较为明显。
2.2 龙眼拟茎点霉诱导子对龙眼胚性愈伤组织生长的影响
2.2.1 诱导时间对龙眼胚性愈伤组织生长的影响
用相同浓度100 mg/L龙眼拟茎点霉诱导子处理不同时间后,龙眼胚性愈伤组织生长情况如图4、5所示。从形态上看,诱导20 d时胚性愈伤组织形态较松散淡黄,颗粒较小,15 d时愈伤组织形态松散浅黄,颗粒较小,可能是龙眼胚性愈伤组织正处于旺盛生长阶段,25 d时形态较为紧实,出现颗粒化现象;25 d增殖系数最高,15 d时增殖系数较小,20 d时增殖系数相对较高处于中间水平。SPSS20.0分析可知,不同诱导天数的诱导子对龙眼胚性愈伤组织增殖生长均具有显著性影响,且经诱导子处理后的龙眼胚性愈伤组织增殖系数整体较低,说明龙眼拟茎点霉诱导子明显抑制龙眼胚性愈伤组织增殖生长。
2.2.2 誘导子浓度对龙眼胚性愈伤组织生长的影响
对不同浓度诱导子处理15 d时的龙眼胚性愈伤组织生长状态和增殖系数进行分析(图6和图7)。从总体上看,与对照相比,不同浓度诱导子对龙眼胚性愈伤组织生长均有抑制作用。而50 mg/L的诱导子有利于龙眼胚性愈伤组织正常形态维持,200 mg/L的诱导子不利于龙眼胚性愈伤组织正常生长,100 mg/L的诱导子处理下龙眼胚性愈伤组织虽颗粒稍大,但色泽较为正常。方差齐次性检验得p=0.11,大于0.05,单因素方差分析的F检验中p=0,小于0.05,说明不同浓度诱导子对龙眼胚性愈伤组织增殖生长具有显著性影响,且添加100和200 mg/L的诱导子对龙眼胚性愈伤组织增殖生长均有极显著性差异,但100 mg/L诱导子相对抑制作用最小,增殖系数为4.94,与对照相比减小40.48%。可见,50~200 mg/L的诱导子对龙眼胚性愈伤组织增殖生长均有抑制作用,而均衡多糖含量与龙眼胚性愈伤组织生长情况,选择添加100 mg/L龙眼拟茎点霉诱导子,暗培养诱导15 d作为本试验龙眼胚性愈伤组织多糖积累的最佳条件。
3 讨论与结论
3.1 龙眼拟茎点霉诱导子促进龙眼胚性愈伤组织多糖积累
本试验研究认为,龙眼拟茎点霉诱导子半活体营养型龙眼内生真菌,其活性诱导物能促进龙眼胚性愈伤组织中多糖含量的积累。白桦悬浮细胞中三萜系化合物会受拟茎点霉诱导子的影响[22],进一步证实了真菌诱导子能有效促进植物次生代谢物合成。真菌诱导子能通过信号识别、信号转导及信号转录介导的胞内应答等途径,调控相关基因的表达进而影响植物次生代谢物积累。龙眼拟茎点霉诱导子来源于龙眼果肉致病内生菌-龙眼拟茎点霉[15],该霉能产生细胞壁降解酶,降解细胞壁多糖,诱发龙眼果腐病[23-24]。陈艺辉等[25]研究发现龙眼拟茎点霉可以促进酚类物质积累使果皮褐变,病原菌侵染早期能引起果实细胞氧化迸发的应激反应,增强植物体免疫防御能力;多糖作为初生代谢产物,也是植物细胞壁主要成分,不仅能作为众多植物次生代谢物合成前体,还具有免疫、防御及抗逆等相关功能[26]。龙眼拟茎点霉诱导子可能通过诱导调控龙眼胚性愈伤组织中某些病原抗性基因,在病原侵染初期通过合成多糖来提高植物自身的免疫防御能力,这与江曙等[27]研究的明党参内生真菌能显著促进明党参多糖积累一致。
研究表明,生物诱导子具有高效、专一及浓度效应等特点[28-29]。本试验认为龙眼拟茎点霉诱导子较其他2种非龙眼内生真菌诱导子更能显著提高龙眼胚性愈伤组织多糖含量。尖孢镰刀菌诱导子对龙眼胚性愈伤组织多糖积累有一定抑制作用,与刘长军等[30]的研究结论并不一致,可能由于同种诱导子响应植物中同种次生代谢物合成的细胞受体的结合位点存在差异。也可能通过钙调素、磷酸肌醇、G蛋白等诱导信号,以IP3-DAG-Ca2+/PKC系统、腺苷酸环化酶调节的cAMP/PKA胞内信号传导的第二信使传递系统,影响次生代谢物合成相关基因的表达,改变次生代谢通路,从而调节次生代谢物积累[31]。因此,研究龙眼本身致病内生菌的生物活性物质适当的诱导时间、浓度及添加时间对龙眼本身多糖和次生代谢物的积累具有重要意义。
3.2 龙眼拟茎点霉诱导子抑制龙眼胚性愈伤组织生长
试验中,添加100 mg/L龙眼拟茎点霉诱导子时发现,龙眼愈伤组织增殖生长抑制率达40.48%,这与陶金华等[32]研究的100 mg/L内生真菌抑制茅苍术细胞生长结果类似。诱导子对次生代谢物调节具有特异性,同时由于诱导产生的抗毒素对细胞自身大多具有毒性,浓度过高有可能抑制细胞生长。且粗诱导物中诱导子的活性成分和拮抗成分相互制约,加量少时拮抗作用小,诱导作用也小,反之亦然。
在前人的研究中,刘长军等[30]研究发现尖孢镰刀菌诱导子在西洋参悬浮细胞培养前期能促进细胞生长,而Zhu等[33]发现蛋白类诱导子在促进灵芝酸合成的同时抑制了细胞生长。可见,诱导子促进细胞生长与次生代谢物合成还可能和诱导子本身的活性成分种类有关,其可能和细胞间信号传导、基因表达调控等机制有关,这与添加非生物因子如激素、光质、营养物质等因素对次生代谢物的诱导有所不同[34]。试验研究结果可为在龙眼细胞组织培养生产中提高多糖含量提供科学依据。
参考文献
[1] Yang B, Jiang Y M, Zhao M M, et al. Structural characterisation of polysaccharides purified from longan(Dimocarpus longan Lour.)fruit pericarp[J]. Food Chemistry, 2009, 115(2): 609-614.
[2] Liu X X, Luo Y Y, Zha C J. Rheological properties of polysaccharides from longan(Dimocarpus longan Lour.)fruit[J]. International Journal of Polymer Science, 2015: 1-5
[3] 蔡英卿, 赖钟雄, 伍妙聪, 等. 龙眼不同品种各营养器官多糖含量分析[J]. 中国生态农业学报, 2007, 15(4): 86-88.
[4] Zhong K, Wang Q, He Y, et al. Evaluation of radicals scavenging, immunity-modulatory and antitumor activities of longan polysaccharides with ultrasonic extraction on in S180 tumor mice models[J]. International Journal of Biological Macromolecules, 2010, 47(33): 56-60.
[5] Yi Y, Zhang M W, Liao S T , et al. Structural features and immunomodulatory activities of polysaccharides of longan pulp[J]. Carbohydrate Polymers, 2012, 87(87): 636-643.
[6] Zhang M, Tang X L, Wang F, et al. Characterization of Lycium barbarum polysaccharide and its effect on human hepatoma cells[J]. International Journal of Biological Macromolecules, 2013, 61(10): 270-275.
[7] 陳建国, 步文磊, 来伟旗, 等. 桑叶多糖降血糖作用及其机制研究[J]. 中草药, 2011, 42(3): 515-520.
[8] 向艳霞, 肖凌云, 张 菊, 等. 黄芪多糖治疗神经系统疾病的作用及其机制研究进展[J]. 中国医院药学杂志, 2016(8): 1-5.
[9] 董慧雪, 周燕蓉, 田奇琳, 等. 不同光质对龙眼胚性愈伤组织类黄酮含量的影响[J]. 热带作物学报, 2014, 35(12): 2 374-2 377.
[10] Lai Z X, Lin Y L. Analysis of the global transcriptome of longan(Dimocarpus longan Lour.)embryogenic callus using Illumina paired-end sequencing[J]. BMC Genomics, 2013(14): 561
[11] Liang W B, Deng B L, Xiao J, et al. Effects of basic media and culture conditions on callus growth and polysaccharide content in Schisandra sphenanthera[J]. Journal of Central South University of Forestry & Technology, 2011, 31(3): 130-133.
[12] Tian Q Q, Lu C M, Li X, et al. Low temperature treatments of rice(Oryza sativa L.)anthers changes polysaccharide and protein composition of the anther walls and increases pollen fertility and callus induction[J]. Plant Cell Tissue & Organ Culture, 2015, 120(1): 89-98.
[13] 金晓锋, 杨永健, 朱莲莲, 等. 铁皮石斛悬浮培养条件筛选及生产多糖[J]. 兰州大学学报: 医学版, 2012, 38(4): 27-30.
[14] Sibylle K, Marilena L, Christne H, et al. Fungal elicitors combined with a sucrose feed significantly enhance triterpene production of a Salvia fruticosa cell suspension[J]. Applied Microbiology and Biotechnology, 2016, 100(16): 7 071-7 082.
[15] 杨 慧, 陈晓梅, 郭顺星. 真菌诱导子对铁皮石斛原球茎多糖含量的影响[J]. 世界科学技术: 中医药现代化, 2009, 11(5): 719-722.
[16] 高兴喜, 姚 强, 王 磊, 等. 真菌激发子对灵芝液体发酵生产多糖和三萜类物质的影响[J]. 食品科学, 2009, 30(23): 309-313.
[17] 张居念, 林河通, 谢联辉, 等. 龙眼果实潜伏性病原真菌的初步研究[J]. 热带作物学报, 2006, 27(4): 78-82.
[18] 赖钟雄, 潘良镇. 龙眼胚性细胞系的建立与保持[J]. 福建农业大学学报, 1997, 26(2): 160-167.
[19] Wang C G, Wu J Y, Mei X G. Enhancement of taxol production andexcretion in Taxus chinensis cell culture by fungal elicitation andmedium renewal[J]. Applied microbiology and Biotechnology, 2001, 55(4): 404-410.
[20] 林艳君, 赖钟雄. 铁皮石斛HDR基因克隆及真菌诱导子对其表达和生物碱含量的影响[J]. 热带作物学报, 2015, 36(4): 680-686.
[21] 毕和平, 周 贤, 陈光英, 等. 苯酚-硫酸法测定破布叶叶的多糖[J]. 海南师范大学学报: 自然科学版, 2010, 23(4): 400-402.
[22] Fan G Z, Zhai Q, Zhan Y. Gene expression of lupeol synthase and biosynthesis of Nitric Oxide in cell suspension cultures of Betula platyphylla, in response to a Phomopsis, elicitor[J].Plant Molecular Biology Reporter, 2012, 31(2): 296-302.
[23] 张居念, 陈艺晖, 林艺芬, 等. 龙眼拟茎点霉(Phomopsis longanae Chi)的生物学特性研究[J]. 热带作物学报, 2013, 34(4): 695-699.
[24] 张洪祥. 龙眼拟茎点霉细胞壁降解酶及其致病机理的研究[D].福州: 福建农林大学, 2010.
[25] 陈艺晖, 林河通, 林艺芬, 等. 拟茎点霉侵染对龙眼果实采后果皮褐变和活性氧代谢的影响[J]. 中国农业科学, 2011, 44(23): 4 858-4 866.
[26] Vorwerk S, Somerville S, Somerville C. The role of plant cell wall polysaccharide composition in disease resistance[J]. Trends in Plant Science, 2004, 9(4): 203-209.
[27] 江 曙, 段金廒, 陶金华, 等. 明党参内生真菌种群的生态分布及其诱导子活性研究[J]. 中草药, 2010, 41(1): 121-125.
[28] 崔晋龙, 付少彬, 高 芬, 等. 真菌诱导植物次生代谢产物积累的信号机制及在药用植物中的应用[J]. 中草药, 2012, 43(8): 1 647-1 651.
[29] Xiao J L, Lai Y C, Li W, et al. Research development of endophytic fungus and defence response mechanism of plant[J]. Botanical Research, 2013, 2(1): 13-17.
[30] 刘长军, 侯嵩生, 李新明, 等. 真菌诱导子对悬浮培养西洋参细胞的生理效应[J]. 植物科学学报, 1996(3): 240-246.
[31] Andrea V, RICARDO B. Molecular aspects of the early stages of elicitation of secondary metabolites in plants[J]. Plant Science, 2007, 172(5): 861-875.
[32] 陶金華, 濮雪莲, 江 曙. 内生真菌诱导子对茅苍术细胞生长及苍术素积累的影响[J]. 中国中药杂志, 2011, 36(1): 27-31.
[33] Zhu L W, Zhong J J, Tang Y J. Significance of fungal elicitors on the production of ganoderic acid and Ganoderma polysaccharides by the submerged culture of medicinal mushroom Ganoderma lucidum[J]. Process Biochemistry, 2008, 43(12): 1 359-1 370.
[34] 徐小萍, 谢燕萍, 慕 尧, 等. HPLC法测定LED不同光质下迷迭香愈伤组织中鼠尾草酸含量[J]. 广东农业科学, 2015, 42(15):77-81.
猜你喜欢
科学与财富(2021年36期)2021-05-10
作文大王·低年级(2019年4期)2019-05-13
世界热带农业信息(2018年5期)2018-11-09
科技视界(2018年1期)2018-03-15
世界热带农业信息(2018年6期)2018-03-05
湖北农业科学(2017年1期)2017-03-09
山东农业科学(2016年12期)2017-01-21
中国现代医生(2016年27期)2016-12-21
学苑创造·A版(2016年9期)2016-10-10
科技视界(2016年5期)2016-02-22
