
昆虫卵黄原蛋白受体(VgRs)及其主要功能综述
2016-08-23 03:13王加伟邹明民杨一帆尤民生
环境昆虫学报 2016年4期
王加伟,彭 露,邹明民,杨一帆,汪 蕾,尤民生
(福建农林大学应用生态研究所, 闽台特色作物病虫生态防控协同创新中心,农业部闽台作物有害生物综合治理重点实验室,福州350002)

昆虫卵黄原蛋白受体(VgRs)及其主要功能综述
王加伟*,彭露*,邹明民,杨一帆,汪蕾,尤民生**
(福建农林大学应用生态研究所, 闽台特色作物病虫生态防控协同创新中心,农业部闽台作物有害生物综合治理重点实验室,福州350002)
卵黄原蛋白受体(VgRs)属于低密度脂蛋白受体家族成员,具有该家族典型的保守结构域,包括配体结合域,表皮生长因子前体同源域,跨膜域,O-联糖功能域,以及胞质尾域。昆虫VgRs通常具有卵巢特异性,是卵黄原蛋白Vg的专一性胞吞作用受体,可介导Vg进入昆虫卵母细胞,而后沉淀积累形成昆虫生殖必须的卵黄蛋白YP。VgRs介导的胞吞作用是一个动态循环过程,它是卵黄发生的基础,对昆虫卵母细胞发育起着至关重要的作用。近年来的研究表明,VgRs不仅与卵巢激活、卵黄发生与卵子形成密切相关,而且在昆虫信息交流、社会分化、行为构建以及免疫调控等中也起到了至关重要的作用,已成为潜在的害虫控制新靶标。本文首次对昆虫VgRs基因的序列信息,分子结构,系统进化,表达模式以及调控功能等方面进行了综述,旨在为了解VgRs基因的研究进展及前景提供参考,对进一步改进害虫生态控制的策略和措施也具有指导意义。
昆虫;卵黄原蛋白受体;表达模式;生殖调控;功能
众所周知,昆虫是动物界中种类最多、数量最大、分布范围最广的类群,除了拥有个体小、食性广、生命周期短、适应进化快等特点外,更为主要的是其强大的生殖能力和特殊的生殖适应性(雷朝亮和荣秀兰,2003)。昆虫作为一种卵生动物,其胚胎发育主要依靠发育中的卵母细胞积累充足的卵黄蛋白(yolk protein, YP)等物质作为生命必须的营养物质(氨基酸、蛋白质、脂类、磷酸盐、碳水化合物、离子和维生素等),该过程也被称为卵黄发生(vitellogenesis),是昆虫生殖调控的核心(Amdametal.,2010; 戈林泉和吴进才,2010)。迄今为止,卵黄原蛋白(vitellogenin,Vg)是所有昆虫中最为丰富的一类YP前体,主要在激素的调控下由雌性昆虫脂肪体细胞合成,并释放到血淋巴,通过受体作用被卵母细胞摄取,最终通过剪切、修饰、加工后以晶体形式沉淀为YP(Matozzoetal., 2008; Tufail and Takeda,2008,2009)。
卵黄蛋白原受体(vitellogenin receptors,VgRs)具有卵巢特异性,是Vg的专一性胞吞作用受体(Tufail and Takeda,2009),与卵生动物,尤其是昆虫的生殖过程密切相关。在VgR介导的Vg胞吞作用过程中,VgR与Vg结合形成的受体—配体复合物引发细胞质膜局部内化(internalization),随后在网格蛋白(clathrin)的作用下形成被膜小窝(coated pit)(Brown and Goldstein,1986),包裹着Vg的被膜小窝逐步深陷,直至脱离质膜形成被膜小泡(coated vesicle),最后转运至卵母细胞内,进入细胞内的被膜小泡脱离包被,Vg/VgR复合体在核内酸化作用下解离,Vg被胞内体包裹后沉淀形成YP,VgR重新回到卵母细胞表面,再次介导Vg转运(Thomas and Alexander,1998; Tufail and Takeda,2008)。可以看出,VgR介导的胞吞作用是一个动态循环过程,且具有专一特异性,既保证了卵母细胞对Vg的高效摄取,又可避免吸入过多的细胞外溶液。
VgR是卵黄发生的基础,对昆虫卵巢的成熟起着至关重要的作用,也是研究控制害虫的潜在靶标(Linetal.,2013)。开展VgR的研究对深入了解昆虫的生殖生理机制、挖掘有效控制害虫的新方法具有重要意义。近年来,昆虫VgRs的研究逐步成为探讨昆虫生殖调控机制的热点,从Ferenz(1993)在飞蝗卵母细胞中纯化、鉴定出第一个VgR蛋白以来,迄今为止,已有包括鳞翅目、蜚蠊目、双翅目、膜翅目、半翅目、鞘翅目,以及虱目在内的30种昆虫的VgRs序列被克隆、测序或预测。为了进一步促进昆虫VgR基因的研究,拓展该领域研究的广度与深度,本文首次全面的对昆虫VgR基因的序列信息、分子结构、系统进化、表达模式以及生殖调控功能等方面进行了综述,旨在为了解VgR基因的研究进展及前景提供参考,对进一步改进害虫生态控制的策略和措施也具有指导意义。
1 昆虫卵黄原蛋白受体基因的克隆
随着分子生物学技术与基因组学数据的不断丰富与发展,目前对于昆虫VgR基因的克隆主要采用3种方式:第一种为genome walking,即分步法获得基因全长。首先根据已有的基因组或转录组信息,筛选获得VgR基因的预测序列,通过分段设计引物,逐段扩增基因全长,最后将获得的各段序列进行整合与拼接。值得注意的是,在进行分段引物设计时,必须包含至少50 bp的重合序列,这样进行拼接时才能保证获得序列的完整性。第二种为常规的RACE (Rapid-Amplification of cDNA Ends)扩增方法,此方法大多应用在目标物种的基因组或转录组信息还未发布,但序列已知的基因克隆,即直接将前人报道和上传的基因序列作为自己克隆的依据。首先设计通用引物获得中间片段,然后根据中间片段分别设计针对5’RACE的下游引物与3’RACE的上游引物,分别获得5端与3端序列,最后将三段序列进行拼接即可获得全长。例如,红火蚁Solenopsisinvicta(Chenetal., 2004)、美洲大蠊Periplanetaamericana(Tufail and Takeda,2005)、马德拉蜚蠊Rhyparobiamaderae(Tufail and Takeda,2007)、斜纹夜蛾Spodopteralitura(Shuetal.,2011)、柞蚕Antheraeapernyi(Liuetal., 2011)、柳蚕Actiasselene(Xuetal., 2012)、家蚕Bombyxmori(Linetal., 2013)、烟粉虱Bemisiatabaci(郭建洋等, 2010)、褐飞虱Nilaparvatalugens(Luetal., 2015)和桔小实蝇Bactroceradorsalis(Congetal.,2015; Linetal.,2015),均采用此法。值得注意的是这种方法对cDNA的质量和引物的特异性要求很高。第三种为根据已知探针进行基因克隆,以物种VgR的cDNA 片段作为分子探针,筛选cDNA 文库,从而获得基因的cDNA 序列,此方法仍需要基因组信息的支持。双翅目的埃及伊蚊AedesaegyptiVgR的基因克隆则采用此方法(Cho and Raikhel,2001)。
自Ferenz首次从飞蝗卵母细胞中纯化、鉴定出第一个VgR蛋白以来(Ferenz,1993),VgR基因的研究越来越成为热点。目前NCBI中已收录的昆虫VgR信息共有385条,已报道的有30种昆虫VgRs基因(表1),其中已克隆验证获得全长序列的有15 种昆虫,通过基因组和转录组信息预测的VgR序列有15种昆虫。
续上表
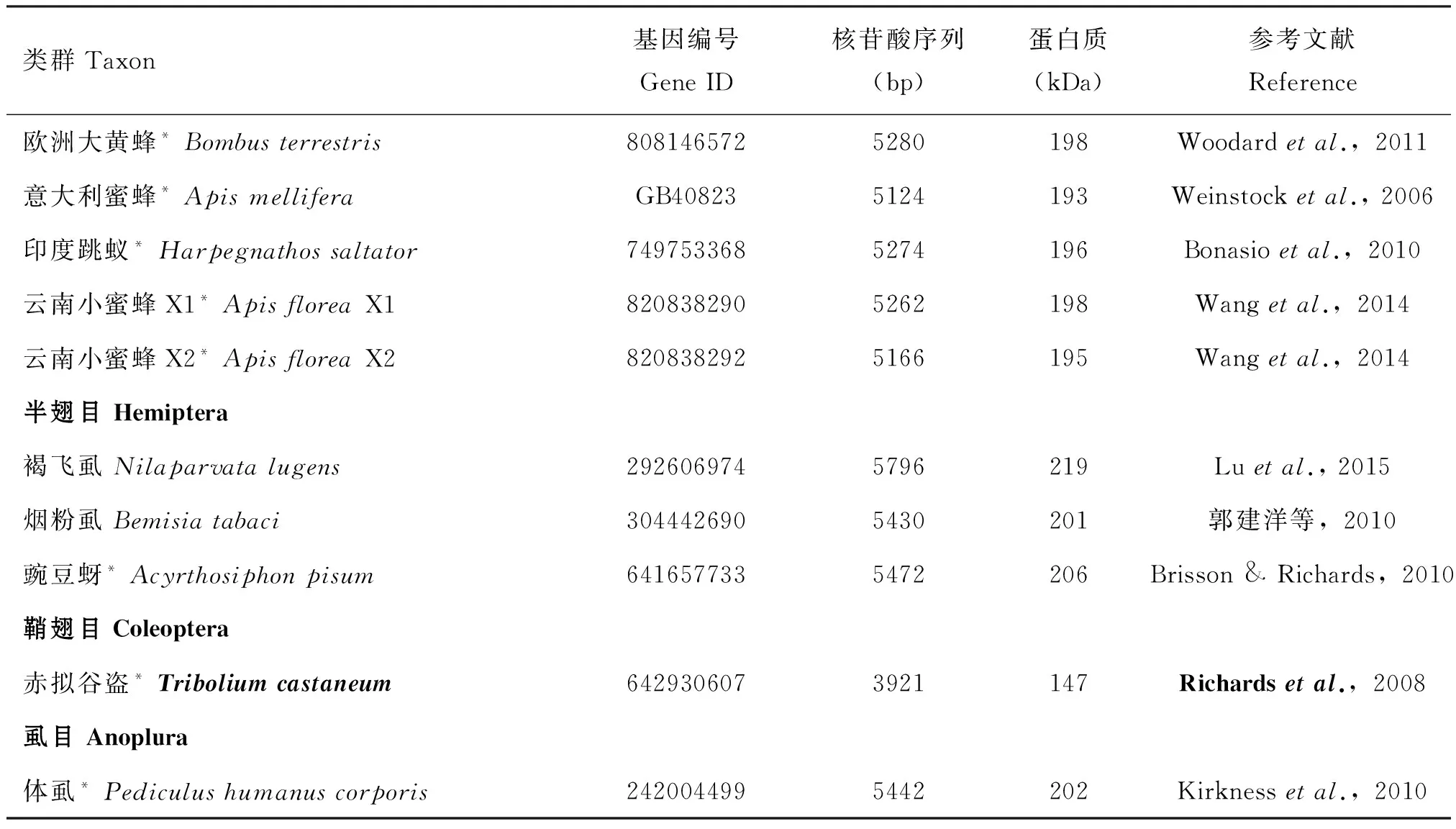
类群Taxon基因编号GeneID核苷酸序列(bp)蛋白质(kDa)参考文献Reference欧洲大黄蜂*Bombusterrestris8081465725280198Woodardetal.,2011意大利蜜蜂*ApismelliferaGB408235124193Weinstocketal.,2006印度跳蚁*Harpegnathossaltator7497533685274196Bonasioetal.,2010云南小蜜蜂X1*ApisfloreaX18208382905262198Wangetal.,2014云南小蜜蜂X2*ApisfloreaX28208382925166195Wangetal.,2014半翅目Hemiptera褐飞虱Nilaparvatalugens2926069745796219Luetal.,2015烟粉虱Bemisiatabaci3044426905430201郭建洋等,2010豌豆蚜*Acyrthosiphonpisum6416577335472206Brisson&Richards,2010鞘翅目Coleoptera赤拟谷盗*Triboliumcastaneum6429306073921147Richardsetal.,2008虱目Anoplura体虱*Pediculushumanuscorporis2420044995442202Kirknessetal.,2010
注:*代表基因组预测获得的VgR序列;基因编号中含前缀字母的来自物种基因组,其他来自NCBI。Note:The asterisk (*) indicates the predictedVgRsequences collecting from the insect genomes; Gene IDs,containing prefix letters are from species genome databases,others from NCBI.
2 昆虫卵黄原蛋白受体的分子结构和特点
利用已经获得或预测的昆虫VgR序列进行保守结构域分析(图1),结果显示昆虫VgRs属于低密度脂蛋白受体(low density lipoprotein receptor, LDLR)家族(Sappington and Raikhel, 1998)。此外,该家族还包括脂蛋白受体(lipoprotein receptor, LpR),超低密度脂蛋白受体(very LDLR,VLDLR)(Sakaietal., 1994),LDLR相关蛋白(LDLR-related protein, LRP)(Herzetal., 1988),以及megalin分子(Saitoetal., 1994)。昆虫VgRs具有一些LDLR家族典型的保守结构域,如配体结合域(ligand-binding domain, LBD),表皮生长因子前体同源域(epidermal growth factor precursor homology domain, EGFD),跨膜域(transmembrane domain, TMD),O-联糖功能域(O-linked sugar domain, OLSD),以及胞质尾域(cytoplasmic domain, CPD)。同一物种不同的LDLR家族成员以及不同物种同一家族成员之间,LBD与EGFD的数量并不相同,可以此作为区分亚家族的标准之一,例如VLDLRs(Takahashietal., 1992; Sakaietal., 1994),脊椎动物VgRs(Bujoetal., 1994; Okabayashietal., 1996)、线虫VgRs(Grant and Hirsh,1999)、以及昆虫LpRs仅含有单个LBD与EGFD结构域,而人类与鸡的LRPs以及magalin分子则含有两个以上的LBD与EGFD结构域,由于昆虫VgRs特异地含有两个LBD和EGFD结构域,因此被划分为一个独立的LDLR亚家族(Tufail and Takeda,2005, 2007)。
配体结合域(LBD)是一段由40个氨基酸左右组成的介导配体和受体相互作用的功能域,含有A型重复序列,每个重复序列包含能够结合钙离子的保守性酸性残基区域和6个以二硫键结合的半胱氨酸残基(Yamamotoetal.,1984 )。昆虫LBD N-末端的A型重复区域用来结合Vg蛋白分子。昆虫VgR的两个LBD配体结合域中分别含有4-5个或7-8个重复基序。
表皮生长因子前体同源域(EGFD)通常位于LBD之后,约40个氨基酸,其中包含B型重复序列与β螺旋域,B型重复序列仍然由6个以二硫键结合的半胱氨酸残基组成,但其绑定模式不同于LBD (Tufail and Takeda,2009)。昆虫VgRs具有2个 EGF同源域,由7个位置相对固定的B型重复序列构成,其中4个位于第一个同源域,3个位于第二个同源域(Tufail and Takeda,2005,2007,2009)。而β螺旋域约由260个氨基酸组成,其中含有6个YWTD重复基序,昆虫VgRs具有3个YWTD基序群集(Jeonetal., 2001)。EGF结构域突变的VgRs虽然可以结合配体蛋白,但不能在酸性条件下发生解离,对家蚕Bombyxmori胚胎具有明显的致死效应(Linetal., 2013)。
O-联糖功能域(OLSD)位于质膜表面,由30个左右的氨基酸组成,富含苏氨酸和丝氨酸残基。大部分昆虫VgRs均含有OLSD,但也有例外,已有的序列预测显示,果蝇、北美大黄蜂、印度跳蚁、豌豆蚜、红火蚁,帝王斑蝶和棉铃虫均不含OLSD(Luetal., 2015)。目前OLSD的功能仍不明确。
跨膜域(TMD)是位于O-联糖功能域的羧基末端的一段短序列,由约24个氨基酸组成,在调节受体功能,以及质膜与胞吞小泡之间的循环通路中具有重要作用,功能类似膜锚(Herz and Bock,2002)。昆虫VgRs大多只有一个TMD,位于C末端,但也有例外,序列预测显示,埃及伊蚊、地中海实蝇、佛罗里达弓背蚁、豌豆蚜、烟粉虱、体虱VgRs等均具有两个TMD,另一个位于N端。TMD大部分为疏水性氨基酸,比如丙氨酸、甘氨酸、异亮氨酸、亮氨酸、苯丙氨酸和缬氨酸等(Tufail and Takeda,2009)。
胞质尾域(CPD)是细胞内的一段区域, 主要通过NPXY(冬酰胺-脯氨酸-X-酪氨酸)内化信号(internalization signal)将受体定位到被膜小窝上,所有LDLR家族成员的胞质尾区至少含有一个NPXY拷贝。NPXY序列呈紧密的发夹结构,主要作为一些连接蛋白与信号分子的结合位点。而大多数昆虫VgRs胞质尾区域的NPXY信号由特殊的LI(亮氨酸-异亮氨酸)内吞信号代替,但也有例外,红火蚁VgRs同时具有NPXY和LI信号(Chenetal.,2004),蜚蠊目昆虫同时具有NPTF及LI信号。除此之外,昆虫VgRs的其他内化信号还包括蟑螂中发现的NPFD基序,该基序与酵母NPFX(1,2)D基序结构类似,能够直接有效的吸收Kex2p受体(Dunn and Hicke,2001; Howardetal., 2002),蚊子中发现的Src同源体3(Src-homology 3,SH3)结合基序PXXP(Hjalmetal., 1996; Sun and Scoutar,1999),以及果蝇VgR胞质尾区存在的AKSAGQF基序,该基序与甘露糖6-磷酸盐受体(mannose 6-phosphate receptor)基序AKGMEQF类似,能够与LL和YXXø (其中X 表示任何氨基酸,ø表示带有疏水侧链的氨基酸)一起发挥功能(Danzeretal., 1997)。昆虫VgRs中内化信号的多样性,说明在受体介导胞吞作用过程中不同昆虫可能会运用不同的信号。由此推断昆虫VgR的结构与蛋白修饰特征可能与其转运过程和效应密切相关。
除了保守结构域分析外,本文还预测了昆虫已有VgR序列的信号肽区域(http://www.cbs.dtu.dk/services/SignalP/)(图1),不同昆虫均含有一条信号肽,位于肽链N端,多从第一个氨基酸序列开始,且长度在18-30 bp不等,主要用于引导VgR蛋白分子进入到卵母细胞膜表面,从而结合Vg。
3 昆虫卵黄原蛋白受体的进化关系
对NCBI中收录的和已发表的15个昆虫基因组中获得的29种昆虫VgRs的氨基酸序列,应用MEGA软件进行了系统进化树分析(图2),结果显示,昆虫VgRs主要分为三大分支,其中鳞翅目昆虫VgRs归属于第I支(下方区域),双翅目昆虫VgRs归属于第II支(中间区域),而大多数昆虫VgRs则归属于第III支(上方区域),第III支又可分为3个亚分支,在进化树上自上而下为膜翅目,鞘翅目、蜚蠊目、虱目、半翅目蚜虫与半翅目。可以看出,同属的进化关系更为接近,E值均在50以上,说明VgR分子进化与物种的进化较一致。

图1 不同昆虫VgR典型特征结构域的比较
注:序列前字母为代表物种VgR,S为信号肽,LBD为A型富含半胱氨酸残基的重复序列,EGF为B型富含半胱氨酸残基的重复序列,O为O-联糖结构域,T为跨膜域,C为胞质尾区。Note:S, signal peptide;LBD, ligand-binding domain;EG, epidermal growth factor precursor homology domain;O, O-linked sugar domain;T, transmembrane domain;C, cytoplasmic domain.Sequences were retrieved from GenBank protein database and genomic database. These include the VgR ofDrosophilamelanogaster(DmVgR),Aedesaegypti(AaVgR),Solenopsisinvicta(SiVgR),Periplanetaamericana(PaVgR),Blattellagermanica(BgVgR),Rhyparobiamaderae(RmVgR),Antheraeapernyi(ApeVgR),Spodopteralitura(SlVgR),Actiasselene(AsVgR),Bemisiatabaci(BtaVgR),Bombyxmori(BmVgR),Nilaparvatalugens(NlVgR),Bactroceradorsalis(BdVgR)Helicoverpaarmigera(HaVgR),Bombusimpatiens(BiVgR),Camponotusfloridanus(CfVgR),Apisdorsata(AdVgR),Nasoniavitripennis(NvVgR),Megachilerotundata(MrVgR),Bombusterrestris(BteVgR),Apismellifera(AmVgR),Harpegnathossaltator(HsVgR),Apisflorea(AfVgR),Ceratitiscapitata(CcVgR),Anophelesgambiae(AgVgR),Acyrthosiphonpisum(ApiVgRs),Danausplexippus(DpVgR),Triboliumcastaneum(TcVgR),Pediculushumanuscorporis(PhcVgR).
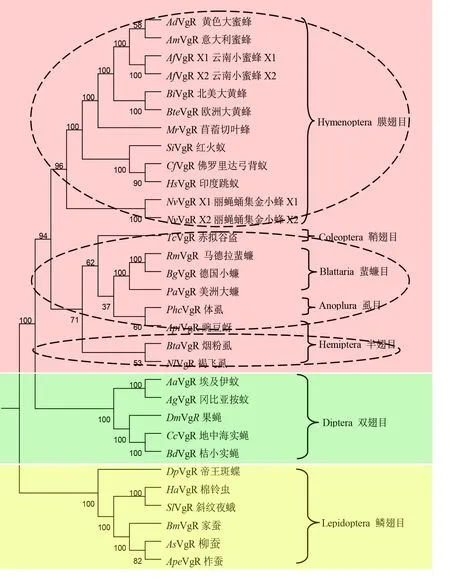
图2 昆虫VgRs系统发育进化树Fig.2 Phylogenetic tree of vitellogenin receptors(VgRs)in insects
4 昆虫VgRs在不同龄期和组织的表达模式
大多数昆虫VgRs的表达发生在初羽化成虫之后,例如埃及伊蚊(Sappingtonetal., 1996)、美洲大蠊(Sappingtonetal., 1996)、马德拉蜚蠊(Tufail and Takeda,2005)、斜纹夜蛾(Shuetal.,2011)、褐飞虱(Luetal., 2015)、烟粉虱(郭建洋, 2010; 程璐等,2013)、桔小实蝇(Linetal.,2015)等。但不同昆虫种类其表达高峰有所差异,埃及伊蚊VgR的转录产物在成虫羽化1 d内急剧增加,吸血24 h后达到峰值(Sappingtonetal., 1996)。美洲大蠊VgR在羽化后1-2 d表达量最高,之后3-9 d逐渐下降,在13 d时已检测不到VgR的表达,然而SDS-PAGE研究发现,VgR蛋白在羽化后5-7 d的积累量达到最高(Tufail and Takeda,2005)。马德拉蜚蠊VgR的表达模式与美洲大蠊较为相似,其转录产物在羽化后第一天达到最高,之后慢慢降低;用SDS-PAGE分析发现成虫羽化后1-2 d内VgR蛋白的表达量较低,之后开始缓慢增加,直到6-9 d时达到峰值(Tufail and Takeda,2007)。这可能由于蛋白开始表达到积累至含量最高存在一定的延时性。斜纹夜蛾VgR的表达在羽化后36 h达到峰值,之后逐渐下降(Shuetal.,2011)。褐飞虱的VgR表达则较延迟,直至成虫第6天达到最高值,之后逐渐下降(Luetal., 2015)。而烟粉虱VgR的表达则在成虫第7天达到最高值(Chengetal.,2013)。以上差异可能与昆虫生殖腺的发育速率以及产卵前期长短密切相关。除此之外,也有研究发现,VgR在雌性家蚕的整个发育周期均有表达,卵期至5龄幼虫的第5天时,VgRs的表达水平较低,随后逐渐升高,在成虫期达到最大值(Linetal., 2013),昆虫VgRs可能识别多种配体和转运其他脂蛋白,由于突变体“缺少Vn”,为了提供足够的营养物质使卵巢发育,VgR可能参与转运了Vg和30 kDa的蛋白(Tufail and Takeda,2009);并且红火蚁的VgR也只在婚飞(mating flights)期交配后表达, 24 h达到峰值,之后逐渐下降,无翼红火蚁VgR不表达,也没有受精现象;红火蚁中存在的婚飞现象是一种繁殖与传播的策略,获得足够的营养物质与发育出翅膀是红火蚁成功婚飞的必备条件(Bryant and Alexander, 2011)。以上研究结果表明,VgR主要出现在交配与生殖前,旨在为昆虫卵巢发育与卵子产生奠定基础(Luetal.,2009)。另外,VgRs可能与一次成功的生殖相偶联,只有在成功的生殖中,VgRs会大量表达,反之则不会,昆虫会维持生殖与生存的平衡,当拥有能够维持生存的能量后,才会进行生殖等一系列基因表达与活动。
大多数研究证实,昆虫VgRs属于卵巢特异性表达蛋白,例如,对斜纹夜蛾、家蚕、褐飞虱、红火蚁不同组织的研究发现,仅在卵巢中检测到VgRs的表达(Shuetal.,2011; Linetal., 2013; Luetal., 2009; Luetal., 2015)。然而现有研究证明,昆虫VgRs的表达并不仅仅局限于卵巢中。桔小实蝇羽化后第4天,VgR则在脂肪体中开始表达,第6天达到最高值,之后开始下降,与卵巢VgR的表达相比较为延迟,但增加速率更快,其卵巢VgRs从雌虫初羽化开始表达,1-3 d内表达水平较低,随后开始缓慢增加,直至10 d后达到峰值(Linetal.,2015)。Liu等(2011)与Qian等(2015)证实,柞蚕与野蚕的VgRs在成虫期时仅在卵巢中表达,而在幼虫期时则在脂肪体与卵巢中均有明显表达。除此之外,对意大利蜜蜂的VgR研究发现,其不仅在卵巢中表达,在头部、中肠及咽下腺也有微量表达(Guidugli-Lazzarinietal.,2008)。而对亚社会性昆虫红斑尼葬甲Nicrophorusvespilloides的研究发现,VgR在其头部大量表达(Roy-Zokanetal.,2015)。VgRs的组织表达差异可能表明了昆虫VgRs的功能多效性。
5 昆虫卵黄原蛋白受体的功能多效性
昆虫卵黄原蛋白Vg在卵子内沉积是卵子发生与胚胎发育的关键,作为Vg的专一性受体,VgR保证了昆虫在卵黄发生过程中获得足够的营养物质,对昆虫卵巢的成熟起着至关重要的作用(Linetal., 2013)。Ciudad等(2006)利用免疫荧光技术监测了德国小蠊卵泡细胞发育过程中的VgR变化,研究发现,6龄幼虫的卵泡细胞中便可检测到微弱的VgR荧光信号,成虫第3天时VgR荧光信号显著加强,并且卵泡饱满;然而,当沉默德国小蠊VgR后,脂肪体中的Vg积累增加,且卵母细胞发育停滞,这说明由于卵母细胞表面VgR减少,导致配体Vg蛋白无法转运至卵母细胞内,从而囤积在脂肪体中。Shu等(2011)研究证实,干扰斜纹夜蛾VgR后,其卵巢发育受到明显抑制,分别下调了82.6%(24 h)、78.8%(48 h)、86.7%(72 h),表现为卵巢干瘪,卵黄蛋白含量极低,并且卵巢管数目明显减少,产卵量显著下降。褐飞虱VgRRNAi后其表达水平显著下降(Luetal., 2015)。未交配的红火蚁VgRRNAi 5 d时,其表达量显著低于对照组,10 d后极显著低于对照组,观察其产卵行为发现干扰后的红火蚁虽然也能产卵,但产卵量与孵化率极显著低于对照组(Luetal.,2009)。综上所述,VgRs在昆虫的生殖中起着不可替代的作用,可能直接影响昆虫卵母细胞的生理与形态发育、卵子和卵巢管的数量以及生殖力。
已有研究证实,昆虫VgRs不仅在卵巢中表达,在脂肪体、甚至头部、中肠、咽下腺都有表达,这说明VgRs不仅在昆虫繁殖中起着至关重要的作用,也可能参与了其他生理与行为活动,尤其是在社会性昆虫中。意大利蜜蜂VgR研究发现,工蜂卵巢产后0-6 h,VgR表达水平较高,由于其配体Vg功能的多样性,作者推测VgR可能协同Vg蛋白参与了后代繁育,蜂王调控、工蜂免疫响应以及激素调控等,但具体的功能仍有待进一步证实(Guidugli-Lazzarinietal.,2008)。对亚社会性昆虫红斑尼葬甲的研究发现,VgR在其头部大量表达,并且当红棕尼葬甲的社会环境发生改变时,比如交配前有无进食,有无进行亲代抚育(parental care);或者从一种社会行为改变至另一种社会行为时,如从交配到亲代抚育,VgR的表达水平会出现显著性变化,由此证实VgR可能在维持红斑尼葬甲生存以及进行正常行为活动中具有重要作用(Roy-Zokan,2015)。桔小实蝇VgR研究发现,其不仅在卵巢中表达,羽化后5-6 d雌虫脂肪体VgR的mRNA表达水平显著升高,可能行使一种自分泌的作用,吸收脂肪体分泌至血淋巴的Vg蛋白,从而保证脂肪体中Vg的贮存,这可能由于Vg不仅具有生殖调控作用(Congetal.,2015),还可能与食物贮存(Guidugli-Lazzarinietal.,2008)、免疫响应(Singhetal.,2013)以及耐受性(Zhangetal.,2014)相关。
6 小结与展望
随着新一代测序技术与生物信息学分析方法的快速发展,推动了昆虫基因组的研究(彭露等,2015),这不仅成功解决了许多受到广泛关注的种群遗传学和进化生态学的热点问题,与此同时,人们对于重要农业害虫的适应性和致害性变异有了更新更全面的认识,为明确关键基因的功能以及寻找控制害虫的新靶标提供了新的思路和研究方向(Heetal., 2012; Youetal., 2013; Xiaetal., 2013; Wangetal., 2014;彭露等,2015)。近年来,昆虫卵黄原蛋白及其受体的研究已成为昆虫生殖生理学领域研究的热点。因此,VgRs基因的分子结构、系统进化、表达模式以及生殖调控功能等也越来越明确。
昆虫VgRs 属低密度脂蛋白LDLR家族,具有该家族典型的结构特征,如配体结合域、表皮生长因子前体同源域、跨膜域、O-联糖功能域、胞质尾域。但与其他LDLR家族成员相比也存在一定差异,如胞质尾域独特的内化信号序列LI。除此之外,不同昆虫同一保守结构域的拷贝数以及内部的重复基序也可能具有差异,例如昆虫VgRs的两个配体结合域中分别含有4-5个或7-8个重复基序(图1);而已有的序列预测显示,果蝇、熊峰、印度跳蚁、豌豆蚜、红火蚁、帝王班蝶,以及棉铃虫均不含O-联糖功能域(图1,Luetal., 2015);并且昆虫VgRs大多只有一个跨膜域,除了埃及伊蚊、地中海实蝇、佛罗里达弓背蚁、豌豆蚜、烟粉虱、体虱VgRs等具有两个跨膜域;其次,昆虫VgRs中内化信号的也具有明显的多样性,包括NPTF、NPFD PXXP、AKSAGQF等。然而,这些结构差异可能产生的VgRs功能多效性仍不清楚,是否与其转运过程和效应密切相关?是否参与了与其对配体的识别等仍需进一步深入研究,前人推测,昆虫VgRs还可识别与Vg结构相似的配体(Chenetal., 2004; Tufail and Takeda,2005; Ciudadetal., 2006; Tufail and Takeda,2007),而果蝇VgR甚至可识别一些不相关的配体(Schonbaumetal., 1995)。也有许多研究证实,LDLR家族基因不仅可作为内吞受体,还可执行细胞信号转导的功能(Herzetal., 2000),而昆虫VgRs胞质尾域多样性的内化信号可能正好证实了其信号转导的功能。因此,明确不同结构的昆虫VgRs的功能多效性可为寻找害虫防治的新靶标提供理论基础。
大多数昆虫VgRs的表达发生在初羽化成虫之后,但不同昆虫种类其表达高峰有所差异,这个可能与昆虫生殖腺的发育速率以及产卵前期长短密切相关。除此之外,还有研究发现,昆虫VgRs的龄期表达模式也有差异,雌性家蚕的整个发育周期均有VgR表达,在成虫期达到最大值(Linetal., 2013);而红火蚁的VgR只在婚飞(mating flights)期交配后表达,婚飞结束后表达消失。同时,不同昆虫VgRs的组织表达模式也有差异。大多认为昆虫VgRs属于卵巢特异性表达蛋白,如斜纹夜蛾、家蚕、褐飞虱、红火蚁(Shuetal.,2011; Linetal., 2013; Luetal., 2009; Luetal., 2015),但也有研究发现,桔小实蝇、柞蚕与野蚕的脂肪体也有表达,尤其是在幼虫期;而一些社会性昆虫的头部、中肠及咽下腺均有VgR的表达。不同昆虫VgR的不同表达模式可能在特定的龄期或组织中具有特定功能,但明确的相关性研究仍有待进一步深入,如VgR在幼虫期脂肪体的表达是否与其能量与营养的获得与积累有关,从而促进了幼虫的生长发育。而VgR在社会性昆虫头部、中肠等的表达是否说明VgR可能参与了其种群的信息交流、社会分工、行为活动等。阐释以上机制将有利于我们开展害虫的生长发育与生殖调控、遗传调控、行为调控等方面的技术研发,以寻求和创新害虫生态治理和可持续控制的策略和手段。
卵黄原蛋白Vg是昆虫卵黄发生的关键物质,也是昆虫生殖调控的基础。随着功能基因组研究的应用与发展,已发现Vg特别是社会性昆虫的Vg具有功能多效性,包括卵巢激活、生殖竞争、社会分化、行为构建、延长寿命、食物利用等多种功能,而VgR作为Vg的专一性受体,是否也行使了多种功能仍不明确,目前,昆虫VgRs的研究主要集中在生殖调控方面,且主要关注了其对昆虫卵巢发育、卵子形成、以及生殖能力的影响,而VgR在社会性昆虫中的研究虽然已涉及行为调控等,但相比Vg而言亟待进一步深入探讨。我们相信,在功能基因组的飞速发展下,将大大提高人们对昆虫VgRs功能机制的认识,并为害虫治理提供新思路。
References)
Amdam GV,Page RE,Fondrk MK,etal. Hormone response to bidirectional selection on social behavior [J].Evol.Dev., 2010, 12: 428-436.
Bryant B,Alexander SR. Programmed autophagy in the fat body ofAedesaegyptiis required to maintain egg maturation cycles [J].PLoSONE,2011,6(11): e25502.
Bonasio R,Zhang G,Ye C,etal. Genomic comparison of the antsCamponotusfloridanusandHarpegnathossaltator[J].Science,2010,329: 1068-1071.
Brisson JA,Richards S. Genome sequence of the pea aphidAcyrthosiphonpisum[J].PLoSBiol., 2010, 8(2): e1000313.
Brown MS, Goldstein JL. A receptor-mediated pathway for cholesterol homeostasis [J].Science,1986, 232: 34-47.
Bujo H,Hermann M,Kaderli MO,etal. Chicken oocyte growth is mediated by an eight ligand repeat member of the LDL receptor family [J].EMBOJ., 1994,13: 5165-5175.
Chen ME,Lewis DK,Keeley LL,etal. cDNA cloning and transcriptional regulation of the vitellogenin receptor from the fire ant,SolenopsisinvictaBuren (Hymenoptera: Formicidae) [J].InsectMol.Biol.,2004, 13: 195-204.
Cheng L, Guo JY,Liu SS,etal. Molecular cloning,sequence analysis and developmental expression profile of vitellogenin receptor gene in the whiteflyBemisiatabaciMiddle East-Asia Minor 1 (Hemiptera: Aleyrodidae) [J].ActaEnt.Sin., 2013,56(6): 584-593. [程璐, 郭建洋, 刘树生, 等. 烟粉虱MEAM1隐种卵黄原蛋白受体基因cDNA的克隆、序列分析及在不同发育时期的表达[J]. 昆虫学报, 2013, 56(6): 584-593.]
Cho KH, Raikhel AS. Organization and developmental expression of the mosquito vitellogenin receptor gene [J].InsectMol.Biol., 2001, 10(5): 465-474.
Ciudad L,Piulachs MD,Bellés X. Systemic RNAi of the cockroach vitellogenin receptor results in a phenotype similar to that of the Drosophila yolkless mutant [J].FEBSJ.,2006, 273: 325-335.
Cong L,Yang WJ,Jiang XZ,etal. The essential role of vitellogenin receptor in ovary development and vitellogenin uptake inBactroceradorsalis[J].Int.J.Mol.Sci.,2015,16: 18368-18383.
Danzer K,Weber B,Hille-Rehfeld A,etal. Identification of three internalization sequences in the cytoplasmic tail of the 46 kDa mannose 6-phosphate receptor [J].Biochem.J.,1997,326: 497-505.
Davis CG,Goldstein JL,Südhof TC,etal. Acid-dependent ligand dissociation and recycling of LDL receptor mediated by growth factor homology region [J].Nature,1987,326: 760-765.
Dunn R,Hicke L. Multiple roles for Rsp5p-dependent ubiquitination at the internalization step of endocytosis [J].J.Biol.Chem.,2001,276: 25974-25981.
Ferenz HJ. Yolk protein accumulation inLocustamigratoria(Orthoptera: Acrididae) oocytes [J].Int.J.InsectMorphol.&Embryol.,1993,22: 295-314.
Ge LQ,WU JC. Research progress in insect vitellin and its hormone regulation [J].ChineseBulletinofEntomology,2010,47(2): 236-246. [戈林泉, 吴进才. 昆虫卵黄蛋白及其激素调控的研究进展[J]. 昆虫知识,2010, 47(2): 247-253.]
Grant B,Hirsh D. Receptor-mediated endocytosis in theCaenorhabditiselagansoocyte [J].Mol.Bio.Cell, 1999,10: 4311-4326.
Guidugli-Lazzarini KR. Expression analysis of putative vitellogenin and lipophorin receptors in honey bee (ApismelliferaL.) queens and workers [J].J.InsectPhysiol.,2008, 54(7): 1138-1147.
Guo JY. Vitellogenesis,Vitellogenin and its Receptor Sequence Analysis of Whiteflies,Bemisiatabaci[D]. Zhejiang:Zhejiang University, Doctoral Dissertation, 2010. [郭建洋. 烟粉虱卵黄发生、卵黄蛋白及其受体基因序列的分析[D]. 浙江: 浙江大学,博士学位论文,2010]
He WY,You MS,Vasseur L,etal. Developmental and insecticide-resistant insights from the de novo assembled transcriptome of the diamondback moth,Plutellaxylostella[J].Genomics,2012,99(3): 169-177.
Herz J,Bock HH. Lipoprotein receptors in the nervous system.Annu.Rev.Biochem., 2002,71: 405-434.
Herz J,Gotthardt M,Willnow TE. Cellular signalling by lipoprotein receptors [J].Curr.Opin.Lipidol., 2000,11: 161-166.
Herz J,Hamann U,Rogne S,etal. Surface location and high affinity for calcium of a 500-kd liver membrane protein closely related to the LDL-receptor suggest a physiological role as lipoprotein receptor [J].EMBOJ.,1988,7(13): 4119-4127.
Hjalm G,Murray E,Crumley G,etal. Cloning and sequencing of human gp330,a Ca2+-binding receptor with potential intracellular signaling properties [J].Eur.J.Biochem.,1996,239: 132-137.
Holt RA,Subramanian GM,Halpern A,etal. The genome sequence of the malaria mosquitoAnophelesgambiae[J].Science,2002,298: 129-149.
Howard JP,Huton JL,Olson JM,etal. Sla1p serves as the targeting signal recognition factor for NPXY(1,2)D-mediated endocytosis [J].J.CellBiol.,2002,157: 315-326.
Jeon H,Meng W,Takagi J,etal. Implications for familial hypercholesterolemia from the structure of the LDL receptor YWTD-EGF domain pair [J].Nat.Struct.Biol., 2001,8: 499-504.
Kirkness EF,Haas BJ,Sun WL,etal. Genome sequences of the human body louse and its primary endosymbiont provide insights into the permanent parasitic lifestyle [J].Proc.Natl.Acad.Sci.,2010,107(27): 12168-12173.
Lei CL,Rong XL,2003. General Entomology[M]. Beijing: China Agriculture Press [M]. [雷朝亮, 荣秀兰, 2003. 普通昆虫学[M]. 北京: 中国农业出版社.]
Lin WJ,Chien CY,Tsai CL,etal. A nonovary-specific vitellogenin receptor from the oriental fruit fly,Bactroceradorsalis[J].Arch.InsectBiochem.,2015, 90(4): 169-180.
Lin Y,Meng Y,Wang YX,etal. Vitellogenin receptor mutation leads to the oogenesis mutant phenotype “scantyvitellin” of the silkworm,Bombyxmori[J].J.Biol.Chem.,2013, 288: 13345-13355.
Liu QN,Zhu BJ,Liu CL,etal. Characterization of vitellogenin receptor(VgRs)from the Chinese oak silkworm,Antheraeapernyi[J].B.Insectol.,2011, 64(2): 167-174.
Lowe TM,Eddy SR. tRNA scan-SE: a program for improved detection of transfer RNA genes in genomic sequence [J].NucleicAcidsRes.,1997,25(5): 955-64.
Lu HL,Vinson SB,Patricia VP. Oocyte membrane localization of vitellogenin receptor coincides with queen flying age,and receptor silencing by RNAi disrupts egg formation in fire ant virgin queens [J].FEBSJ.,2009,276: 3110-3123.
Lu K,Shu YH,Zhou JL,etal. Molecular characterization and RNA interference analysis of vitellogenin receptor fromNilaparvatalugens[J].J.InsectPhysiol., 2015, 73: 20-29.
Matozzo V,Gagnéb F,Marina MG,etal. Vitellogenin as a biomarker of exposure to estrogenic compounds inaquatic invertebrates: A review [J].Environ.Int., 2008,34: 531-545.
Okabayashi K,Shoji H,Nakamura T,etal. cDNA cloning and expression of theXenopuslaevisvitellogenin receptor [J].Biochem.Biophys.Res.Commun., 1996,224: 406-413.
Peng L,He WY,Xia XF,etal. Prospects for the management of insect pests in the genomic era [J].ChineseJournalofAppliedEntomology,2015,52(1): 1-22. [彭露, 何玮毅, 夏晓峰, 等. 基因组学时代害虫治理的研究进展及前景.应用昆虫学报[J], 2015,52(1): 1-22.]
Qian C,Fu WW,Wei GQ,etal. Identification and expression analysis of vitellogenin receptor from the wild silkworm,Bombyxmandarina[J].Arch.InsectBiochem.Physiol.,2015,89(4): 181-192.
Richards S,Gibbs RA,Weinstock GM,etal. The genome of the model beetle and pestTriboliumcastaneum[J].Nature,2008,452: 949-955.
Roy-Zokan EM,Cunningham CB,Hebb LE,etal. Vitellogenin and vitellogenin receptor gene expression is associated with male and female parenting in a subsocial insect [J].Proc.R.Soc.B., 2015, 282: 0787.
Sadd BM,Barribeau SM,Bloch G,etal. The genomes of two key bumblebee species with primitive eusocial organization [J].GenomeBiol.,2015,16: 76.
Sakai J,Hirano A,Takahashi H,etal. Structure,chromosome location and expression of the human very low density lipoprotein receptor gene [J].J.Biol.Chem., 1994,269: 2173-2182.
Saito A,Pietromonaco S,Loo AK-C,etal. Complete cloning and sequencing of rat gp 330/‘megalin’,a distinctive member of the low density lipoprotein receptor gene family [J].Proc.Natl.Acad.Sci., 1994,91: 9725-9729.
Salvemini M,Robertson M,Aronson B,etal.Ceratitiscapitatatransformer-2 gene is required to establish and maintain the autoregulation of Cctra,the master gene for female sex determination [J].Int.J.Dev.Biol.,2009,53(1): 109-120.
Sappington TW,Kokoza VA,Cho WL,etal. Molecular characterization of the mosquito vitellogenin receptor reveals unexpected high homology to the Drosophila yolk protein receptor [J].Proc.Natl.Acad.Sci., 1996, 93: 8934-8939.
Sappington TW, Raikhel AS. Ligand-binding domains in vitellogenin receptors and other LDL-receptor family members share a common ancestral ordering of cysteine-rich repeats [J].J.Mol.Evol.,1998, 46(4): 476-487.
Schonbaum CP,Lee S,Mahowald AP. The Drosophila yolkless gene encodes a vitellogenin receptor belonging to the low density lipoprotein receptor superfamily [J].Proc.Natl.Acad.Sci.,1995, 92: 1485-1489.
Shu YH,Wang JW,Lu K,etal. The first vitellogenin receptor from a Lepidopteran insect: Molecular characterization,expression patterns and RNA interference analysis [J].InsectMol.Biol.,2011, 20: 61-73.
Singh NK, Pakkianathan BC,Kumar M,etal. Vitellogenin from the silkworm,Bombyxmori: An effective anti-bacterial agent [J].PLoSONE,2013,8(9): e73005.
Sun XM,Scoutar AK. Expression in vitro of alternatively spliced variants of the messenger RNA for human apolipoprotein E receptor-2 identifies in human tissues by ribonuclease protection assays [J].Eur.J.Biochem., 1999,262: 230-239.
Takahashi S,Kawarabayasi S,Nakai T,etal. Rabbit low density lipoprotein receptor-like protein with distinct ligand specificity [J].Proc.Natl.Acad.Sci., 1992,89: 9252-9256.
Thomas WS,Alexander SR. Molecular characteristics of insect vitellogenins and vitellogenin receptors [J].InsectBiochem.Mol.Biol., 1998,28: 277-300.
Tufail M,Takeda M. Molecular cloning,characterization and regulation of the cockroach vitellogenin receptor during oogenesis [J].InsectMol.Biol., 2005, 14: 389-401.
Tufail M,Takeda M. Molecular cloning and developmental expression pattern of the vitellogenin receptor from the cockroach,Leucophaeamaderae[J].InsectBiochem.Mol.Biol., 2007, 37: 235-245.
Tufail M,Takeda M. Molecular characteristics of insect vitellogenins [J].J.InsectPhysiol.,2008,54: 1447-1458.
Tufail M,Takeda M. Insect vitellogenin/lipophorin receptors: Molecular structures,role in oogenesis,and regulatiory mechanisms [J].J.InsectPhysiol.,2009, 55: 87-103.
Wang AR,Kim MJ,Lee JY,etal. The mitochondrial genome of the black dwarf honey bee,Apisandreniformis(Hymenoptera: Apidae) [J].Mitochondr.Dna, 2014, 24(3): 208-210.
Wang XH,Fang XD,Yang PC,etal. The locust genome provides insight into swarm formation and long-distance flight [J].Nat.Commun., 2014,5: 2957.
Weinstock GM,Robinson GE,Gibbs RA,etal. Insights into social insects from the genome of the honeybeeApismellifera[J].Nature,2006,443: 931-949.
Woodard SH,Fischman BJ,Venkat A,etal. Genes involved in convergent evolution of eusociality in bees [J].Proc.Natl.Acad.Sci.,2011,108(18): 7472-7.
Xia XF,Zheng DD,Zhong HZ,etal. DNA sequencing reveals the midgut microbiota of diamondback moth,Plutellaxylostella(L.) and a possible relationship with insecticide resistance [J].PLoSONE,2013,8(7): e68852.
Xu YY,Zhu BJ,Liu QN,etal. Prokaryotic expression of vitellogenin receptor gene ofActiasseleneHubner [J].Afr.J.Agr.Res.,2012,7(21): 3166-3174.
Yamamoto T,Davis CG,Brown MS,etal. The human LDL receptor: A cysteine-rich protein with multiple Alu sequences in its Mrna [J].Cell, 1984, 39(1): 27-38.
You MS,Yue Z,He WY,etal. A heterozygous moth genome provides insights into herbivory and detoxification [J].Nat.Genet., 2013,45(2): 220-225.
Zhan S,Merlin C,Boore JL,etal. The monarch butterfly genome yields insights into long-distance migration [J].Cell,2011,147(5): 1171-1185.
Zhang WN, Xiao HJ,Liang GM,etal. Tradeoff between reproduction and resistance evolution to Bt-toxin inHelicoverpaarmigera: Regulated by vitellogenin gene expression [J].Bull.Entomol.Res.,2014,104: 444-452.
A review of insect vitellogenin receptors (VgRs) and their fundamental functions
WANG Jia-Wei*, PENG Lu*,ZOU Ming-Min,YANG Yi-Fan,WANG Lei, YOU Min-Sheng**
(Institute of Applied Ecology,Fujian Agriculture and Forestry University, Fujian-Taiwan Joint Innovation Centre for Ecological Control of Crop Pests,Key Laboratory of Integrated Pest Management of Fujian and Taiwan, China Ministry of Agriculture,Fuzhou 350002,China)
Insect vitellogenin receptors (VgRs) belong to the low-density lipoprotein receptor (LDLR) gene superfamily,which share a group of five structural domains: ligand-binding domain (LBD),epidermal growth factor precursor homology domain (EGFD), transmembrane domain (TMD), O-linked sugar domain (OLSD),and cytoplasmic domain (CPD). The expression studies demonstrate that insect VgRs are usually ovary-bound receptors involved in mediating dynamic endocytosis of the major yolk protein precursors Vg. VgR is the basic substance for vitellogenesis,and dominates the oocyte development of insects. In addition,recent advances in this area have shown that insect VgRs are not only associated with the ovary activation, vitellogenesis,and oogenesis,but also play a critical role in information exchange,labor differentiation,behavior formation,and immune regulation,which has become a potential new target for pest control. This article presents an overview of the research progress in insectVgRs, including their genomic information, molecular structures, phylogenetic relationships,expression profiles and regulating functions. We believe that this review article addresses available literature, and provides potential prospects with the hope to improve the genetic strategies and tactics currently employed in ecologically-based pest management.
Insect; vitellogenin receptor; expression profiling; reproductive regulation; functions
国家自然科学基金(31320103922, 31230061, 31401744)
*共同第一作者, 王加伟,男,硕士生,主要从事昆虫分子生物学方面的研究,E-mail: wangjiawei0908@sina.com;彭露,女,讲师,主要从事昆虫分子生物学与基因组学方面的研究,E-mail: penglu819@fafu.edu.cn
**
Author for correspondence,E-mail:msyou@iae.fjau.edu.cn
2016-03-31;接受日期Accepted:2016-05-18
Q963
A
1674-0858(2016)04-0801-12
猜你喜欢
中国畜牧杂志(2021年5期)2021-12-05
中国生殖健康(2020年8期)2021-01-18
中国生殖健康(2020年2期)2021-01-18
工业催化(2020年9期)2020-11-13
无机化学学报(2020年7期)2020-07-20
中国畜牧杂志(2020年6期)2020-07-11
中国生殖健康(2018年1期)2018-11-06
中国生殖健康(2018年2期)2018-11-06
无机化学学报(2018年8期)2018-08-01
河南畜牧兽医(2017年8期)2017-11-24
