
鬃异源棘尾虫的形态学及细胞发生学研究*
2016-11-17 09:55罗晓甜李俐琼马洪钢胡晓钟
中国海洋大学学报(自然科学版) 2016年11期
罗晓甜, 李俐琼,2, 马洪钢, 胡晓钟**
(1.中国海洋大学水产学院,山东 青岛 266003; 2.中国农业大学烟台研究院海洋学院,山东 烟台 264670)
鬃异源棘尾虫的形态学及细胞发生学研究*
罗晓甜1, 李俐琼1,2, 马洪钢1, 胡晓钟1**
(1.中国海洋大学水产学院,山东 青岛 266003; 2.中国农业大学烟台研究院海洋学院,山东 烟台 264670)
利用活体观察和蛋白银染色技术对鬃异源棘尾虫(Tetmemenapustulata(Müller, 1786) Eigner, 1999)漠河种群的形态学及细胞发生学进行了详尽的研究。研究表明:本种群与其他种群在活体形态和纤毛图式特征方面十分吻合。无性生殖期间细胞发生学表现出的主要特征如下:1)老口围带为前仔虫所继承,老波动膜仅在前部发生更新;2)后仔虫口原基为表层独立发生;3)老棘毛II/2、III/2与IV/3、IV/2、V/4分别参与前、后仔虫额-腹-横棘毛原基的构建;4)前、后仔虫额-腹-横棘毛原基I-VI以“1∶3∶3∶3∶4∶4”的方式分化形成棘毛;5)前、后仔虫的缘棘毛原基分别由老结构反分化形成;6)背触毛发生为尖毛虫模式,但2列背缘触毛原基源于右缘棘毛原基前端断裂。本研究为该种背缘触毛原基起源的首次揭示。
纤毛虫; 腹毛亚纲; 散毛目; 尖毛虫科; 细胞发生
尖毛虫科(Oxytrichidae)纤毛虫广泛分布在海洋、淡水、土壤及高盐高碱等极端生境中,目前已知近50属200余种,是腹毛亚纲(Hypotrichia)中包含种数最多的阶元之一[1-5]。尖毛类纤毛虫因属、种间具有极相似的形态特征(例如体形、大小、纤毛图式、皮层结构等)而导致种类划分相当困难。然而,许多诸如额-腹-横棘毛的产生方式、口原基的发生位置,缘棘毛的演化方式,特别是背触毛的发生模式等细胞发生学特征在不同属种间存在差异,而成为阶元划分、种类区分的重要依据[1,5-6]。
其中棘尾虫属Stylonychia由Ehrenberg于1830年建立,模式种为贻贝棘尾虫Stylonychiamytilus(Müller, 1773) Ehrenberg, 1830。在Berger有关尖毛虫科的专著中,该属包含11个种[1]。21世纪以来,又有4个新的物种:安默曼棘尾虫StylonychiaammermanniGupta et al., 2001,哈尔滨棘尾虫StylonychiaharbinensisShi and Ammermann, 2004,双峰棘尾虫StylonychiagibberaFoissner and heber, 2016和类南方棘尾虫StylonychianotophoridesFoissner, 2016先后被报道, Foissner 的研究认为哈尔滨棘尾虫Stylonychiaharbinensis可能与安默曼棘尾虫Stylonychiaammermanni同物异名[3, 7-8]。Eigner根据细胞发生学特征及分子系统学信息为其中的2个种建立了新属,异源棘尾虫属TetmemenaEigner, 1999,包括模式种鬃异源棘尾虫Tetmemenapustulata(基原异名:Keronapustulata)和贪食异源棘尾虫Tetmemenavorax(基原异名:Stylonychiavorax)[9-10]。棘尾虫属与异源棘尾虫属的主要区别就在于:前者体前端较体后端明显宽阔,虫体坚硬且横棘毛明显分为2组;而后者虫体椭圆形或卵圆形,虫体半坚硬,横棘毛不分组呈“J”形排布。鬃异源棘尾虫最早由 Müller于1786年报道;随后,国外多位学者对该种进行了研究,其中Wirnsberger等.对奥地利萨尔斯堡月亮湖种群的研究因具有详细的活体形态和纤毛图式及细胞发生学特征、统计数据而最为可靠[11]。国内对该种有过3次报道:珠江种群、武汉东湖种群及哈尔滨种群,其中珠江种群有详细的活体形态、纤毛图式及统计数据但无细胞发生学信息,哈尔滨种群有发生学信息,但是形态描述十分简单,武汉东湖种群形态特征描述简单且无发生学信息[12-14]。
本文为该种在黑龙江省漠河县的首次报道,给出无性生殖期间皮层纤毛器和核器的演化过程进行了详细的跟踪观察,以期为纤毛虫的地理分布等研究提供信息支持。
1 材料与方法
纤毛虫样品于2007年8月17日采自黑龙江省漠河县一淡水池塘,分离后成功建立了实验室纯培养,培养方法详见文献[15]。显微镜明视野及微分干涉下进行活体观察及显微拍照。蛋白银制片方法详见文献[16]。在40~1 000×放大倍数下测量活体大小,在1 000×放大倍数下测量蛋白银制片后的细胞并绘图。细胞发生过程中出现的新结构在线条图中用黑色阴影代表,老结构则用轮廓线表示。文中所用名词术语参照了详细的活体形态和纤毛图式特征及统计数据并对其文献[1]。
2 实验结果
2.1 形态学(见图1和表1)
鬃异源棘尾虫Tetmemenapustulata(Müller, 1786) Eigner, 1999属于(腹毛亚纲 Hypotrichia Stein, 1859)(尖毛虫科 Oxytrichidae Ehrenberg, 1838)(异源棘尾虫属TetmemenaEigner, 1999)
虫体活体大小的变化幅度较大,约(45~125) μm×(25~85) μm,通常为阔椭圆形或卵圆形,前后两端钝圆(见图1A,G)。虫体背腹扁平,厚度:体宽比率约为2∶3~1∶2(见图1F)。皮膜薄而坚实,不具伸缩性。表膜下未见皮层颗粒。虫体内质透明,充满大小不等(直径2~6 μm)的食物泡及内储颗粒。伸缩泡单一,直径约为10 μm,位于虫体左侧中部靠前部位(见图1A、G,短箭头)。包囊直径约40~55 μm,皮膜表面多棘刺状突起(图1D、E、I)。
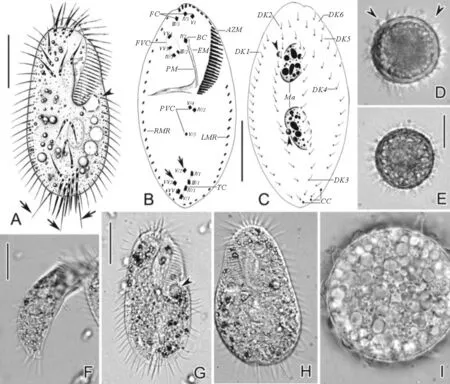
(A.典型个体腹面观,长箭头示尾棘毛,短箭头示伸缩泡;B、C.纤毛图式腹面观及背面观,长箭头示横前腹棘毛,短箭头示小核;D、E.包囊,短箭头示皮膜表面棘刺状突起;F.侧面观,示厚幅比例;G.典型个体腹面观,短箭头示伸缩泡;H.稍受挤压个体背面观;I.包囊内质。AZM:口围带;BC:口棘毛;CC:尾棘毛;DK1-6:背触毛列1-6;EM:口内膜;FC:额棘毛;FVC:额腹棘毛;LMR:左缘棘毛列;Ma:大核;PM:口侧膜;PVC:口后腹棘毛;RMR:右缘棘毛列;TC:横棘毛;图B中I1-VI4,每列额-腹-横棘毛原基产生的棘毛。比例尺:40 μm(A-C,F,G);30 μm(E)。A. Ventral view of a representative individual, arrows indicate the stiff caudal cirri, arrowhead shows the contractile vacuole; B, C. Ventral and dorsal view of infraciliature, arrows indicate pretransverse ventral cirri, arrowheads indicate micronuclei; D, E. Resting cysts, arrowheads mark the spiky surface; F. Lateral view, to show the dorsal-ventrally flattened ratio; G. Ventral view of a typical individual, arrowhead indicates the contractile vacuole; H. Dorsal view of a slightly squeezed specimen; I. Cytoplasm of the resting cyst. AZM: Adoral zone of membranelles; BC: Buccal cirrus; CC: Caudal cirri; DK1-6: Dorsal kineties 1-6; EM: Endoral membrane; FC: Frontal cirri; FVC: Frontoventral cirri; LMR: Left marginal row; Ma: Macronuclear nodules; PM: Paroral membrane; PVC: Postoral ventral cirri; RMR: Right marginal row; TC: Transverse cirri; I-VI: Frontal-ventral-transverse cirral anlagen I-VI, 1-4 (in B), cirri within each anlage. Scale bars = 40 μm (in A-C, F, G); 30 μm (in E).)
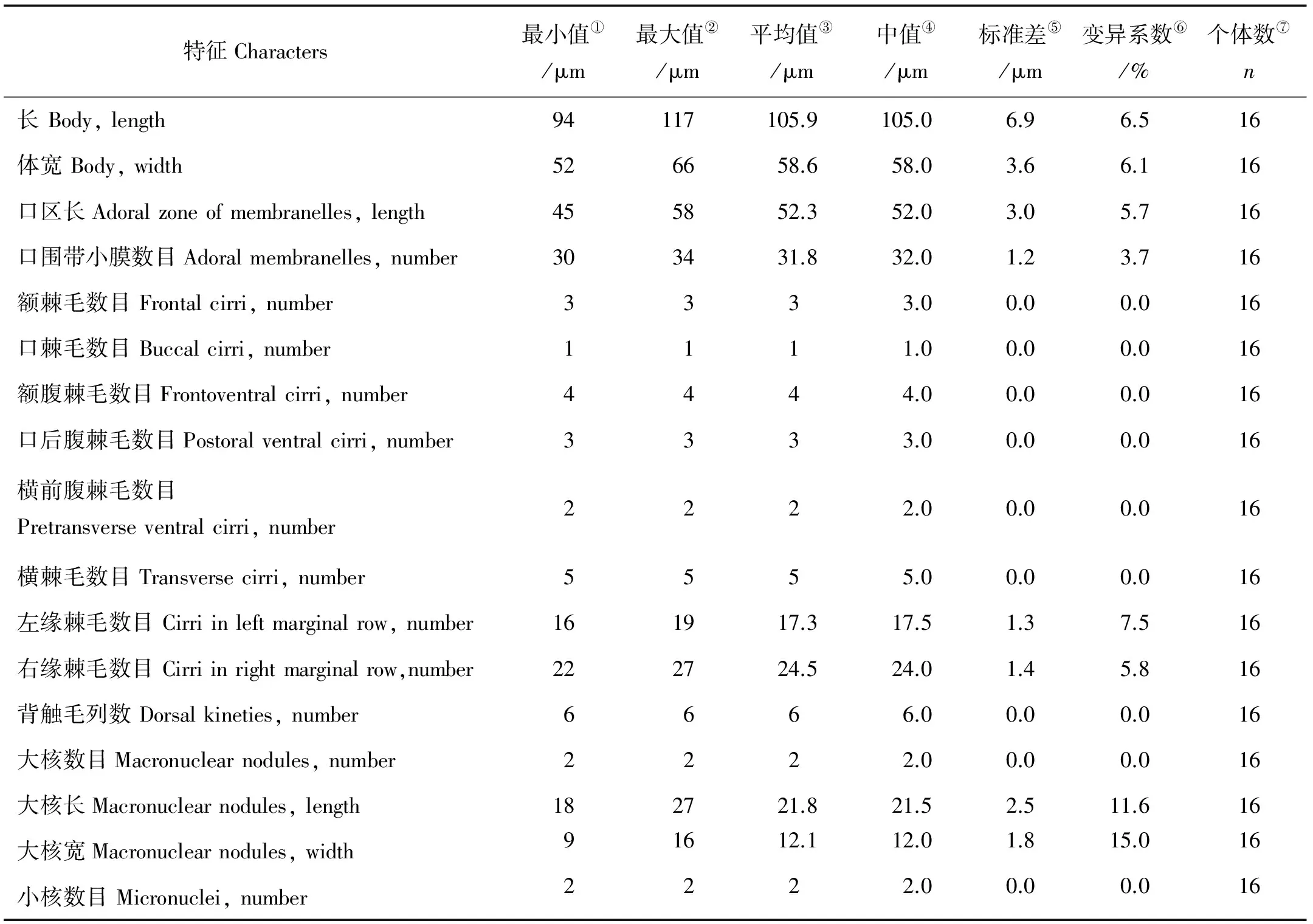
图1 鬃异源棘尾虫的活体形态(A、D~I)和纤毛图式(B、C)
注:所有数据均自蛋白银制片。All data are based on protargol impregnated specimens.
①Minimum;②maximum;③arithmetic mean; ④median value;⑤standard deviation;⑥coefficient of variation in; ⑦number of specimens examined
口区约占体长的50%,口围带远端绕至虫体顶端右侧,含小膜30~34片。波动膜包括口内膜和口侧膜,两者直且平行排布,近等长,前者略前置。额-腹-横棘毛数目及排布十分稳定:额棘毛(FC)恒为3根(I/1,II/3,III/3),较粗壮,其中右侧额棘毛邻近口围带远端;单一口棘毛(BC,II/2)位于波动膜顶端,额腹棘毛(FVC)4根(III/2,IV/3,VI/3,VI/4),呈“V”形排布,且III/2与IV/3之间的距离比IV/3与VI/3之间的距离小;口后腹棘毛(PVC)3根(IV/2,V/3,V/4),略呈“>”形排列,且IV/2与V/4之间距离较小;横前腹棘毛2根(V/2,VI/2);横棘毛(TC)5根(II/1-VI/1),呈“J”形排布。左、右缘棘毛各一列,分别包含16~19根和22~27根棘毛,其后端几联合。背触毛为“4+2”模式,即包含完整的第1~4列背触毛和体右上方的2列短的背缘触毛列。针状尾棘毛(CC)3根,位于虫体尾端,活体长约25 μm因而在200 ×放大倍数下明显可见。大核椭球状,恒为2枚,大小约22 μm × 12 μm,位于体中部,呈前后排布;每一枚大核旁伴随一枚球形小核,直径约2 μm(见图1B、C)。
2.2 细胞发生(见图2~5)
2.2.1 口器 发生早期,口后腹棘毛左侧与左缘棘毛列右侧独立出现了一列小的无序排列的毛基粒群,即后仔虫的口原基(见图2A、B;图4A-C;长箭头)。随后,此原基因毛基粒的增殖而扩大并开始由前至后、由右至左组装形成新的小膜(见图2C,长箭头)。进一步发育,新小膜逐渐增多,新的口围带前端逐渐向右弯曲(见图2D、E、G;图3A)。至发生中期,小膜组装完毕(见图3C)。最终,后仔虫新形成的口围带迁移到既定位置(见图3E、G)。
在整个细胞发生过程中,老口围带小膜不发生变化,由前仔虫完全继承(见图2A~E、G;图3A、C、E、G)。
在后仔虫口原基发生不久,其右前方分化出波动膜原基(额-腹-横棘毛原基I)(见图4E,长箭头)。伴随着毛基粒的增殖,波动膜原基逐渐拉长,并在原基前端产生1根额棘毛(见图2E,G;图3A)。随后,波动膜原基逐渐纵裂形成口内膜和口侧膜(见图3C、E、G)。
发生早中期,老的波动膜前端1/5~1/4的部分开始反分化形成前仔虫的波动膜原基(额-腹-横棘毛原基I)(见图2C、D,短箭头)。伴随着毛基粒的增殖,波动膜原基逐渐拉长并纵裂形成前仔虫口内膜和口侧膜的前段,并在原基的前端产生1根额棘毛(见图2G;3A)。随后,新形成的波动膜前端与未发生变化的下端的老结构拼接形成前仔虫完整的波动膜(见图3C、E、G)。
2.2.2 腹面棘毛 发生早中期,后仔虫口原基右前方形成额-腹-横棘毛原基(FVTA)I-III,老口后腹棘毛IV/2和V/4分别反分化形成原基IV和V-VI;前仔虫中,口棘毛(II/2)反分化形成FVTA II,最左侧额腹棘毛(III/2)反分化形成FVTA III,最下方额腹棘毛(IV/3)参与形成FVTA IV-VI(见图2D、E)。随后,FVTA伸长为长条带状并分段化(见图2E、G;4H~J;5C~F)。随着进一步分化,FVTA I-VI最终按1∶3∶3∶3∶4∶4的方式分化为18根棘毛并迁移到既定位置:FVTA I,II,III的前端各贡献出1根棘毛,并向前迁移至口围带顶端附近而形成额棘毛。原基II中部分化出1根棘毛,在发生的后期会沿着波动膜原基迁移至波动膜前端形成口棘毛。原基III,IV和VI分别贡献1、1、2根棘毛(III/2,IV/3,VI/3,VI/4),最终迁移到额区形成额腹棘毛。原基IV和V分别贡献1、2根棘毛(IV/2,V/3,V/4)而形成口后腹棘毛。原基V和VI各贡献1根棘毛(V/2,VI/2)形成横前腹棘毛。原基II-VI最下端各贡献出1根粗壮的横棘毛(II/1-VI/1),定位于虫体亚尾端(见图3A、C、E、G;5I~L)。原基发育期间,未参与形成原基的老棘毛逐渐被重吸收(见图3A、C、E、G)。

(A~C.早期发生个体腹面观,长箭头示后仔虫口原基;D.早中期发生个体腹面观,长箭头示前后仔虫额-腹-横棘毛原基,短箭头示反分化的老波动膜;E、F.同一早中期发生个体腹面及背面观,示反分化的棘毛(见图E长箭头)及背触毛原基(见图F长箭头);G、H.同一发生个体腹面及背面观,示左(见图G短箭头)右(见图G长箭头)缘棘毛原基及大核复制带(见图H长箭头)。图G中I-VI表示额-腹-横棘毛原基I-VI。比例尺:50 μm。A-C. Ventral views of early dividers, arrows indicate oral primordium of the opisthe; D. Ventral view of an early-middle divider, arrows mark the fronto-ventral-transverse cirral anlagen, arrowhead indicates the dedifferentiating undulating membranes; E, F. Ventral and dorsal view of an early-middle divider, showing the dedifferentiating cirri (arrows in E) and the dorsal kinety anlagen (arrows in F); G, H. Ventral and dorsal view of the same divider, to show the left (arrowheads in G) and right (arrows in G) marginal row anlagen and the replication bands of macronuclear segments (arrows in H). I-VI in G, fronto-ventral-transverse cirral anlagen I-VI. Scale bar=50 μm.)
图2 鬃异源棘尾虫发生早期(A~C)至早中期(D~H)纤毛图式
Fig.2 Morphogenesis ofTetmemenapustulataafter protargol impregnation from early to early-middle stages
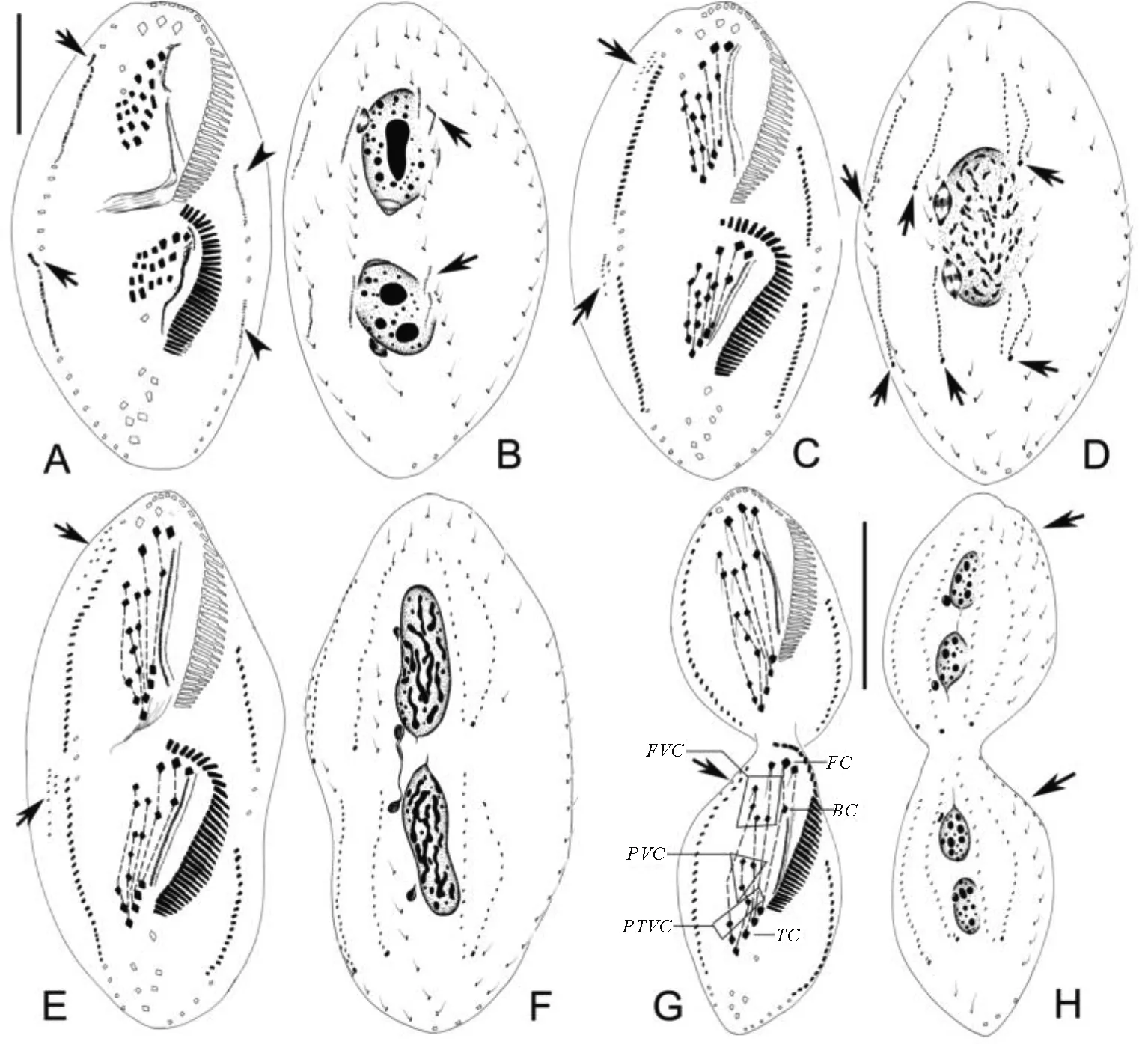
(A、B.同一发生个体腹面及背面观,示右缘棘毛原基向前分裂出的小片断(见图A长箭头),左缘棘毛原基(短箭头)及第3列背触毛原基的断裂(见图B长箭头);C、D.中期发生个体腹面及背面观,示新产生的背缘触毛原基(见图C长箭头)及新的尾棘毛(见图D长箭头);E、F.同一晚期发生个体腹面及背面观,示背缘触毛列(见图E长箭头);G、H.即将分裂的末期发生个体腹面及背面观,示新棘毛的迁移及迁移到背面的背缘触毛(长箭头)。BC:口棘毛;FC:额棘毛;FVC:额腹棘毛;PTVC:横前腹棘毛;PVC:口后腹棘毛; TC:横棘毛。比例尺:30 μm(A,B);50 μm(G,H)。A, B. Ventral and dorsal view of the same divider, showing the small segments split anteriorly from right marginal row anlagen (arrows in A), left marginal row anlagen (arrowheads), and the fragmentation of 3rd dorsal kinety anlagen (arrows in B); C, D. Ventral and dorsal view of a middle divider, to show the newly formed dorsomarginal kinety anlagen (arrows in C) and the new caudal cirri (arrows in D); E, F. Ventral and dorsal view of a late divider, to show the newly formed dorsomarginal kineties (arrowheads in E); G, H. Ventral and dorsal view of the same divider which is about to separate, to show the migration of new cirri and the dorsomarginal kineties migrated to the dorsal side (arrows). BC: Buccal cirri; FC: Frontal cirri; FVC: Frontoventral cirri; PTVC: Pretransverse ventral cirri; PVC: Postoral ventral cirri; TC: Transverse cirri. Scale bars=30 μm (in A, B); 50 μm (in G, H).)
图3 鬃异源棘尾虫发生早中期至末期纤毛图式
Fig.3 Morphogenesis ofTetmemenapustulataafter protargol impregnation from early-middle to late stages
2.2.3 缘棘毛 发生早中期,右缘棘毛列前端1根棘毛反分化形成前仔虫右缘棘毛原基(见图2E;图4G)。随后,前、后仔虫的左、右缘棘毛原基均由老结构反分化形成,且右缘棘毛原基(见图2G;图5E,长箭头)发育比左边(见图2G,短箭头;图5F,长箭头)更早更快。缘棘毛原基不断增殖,分化为棘毛并向两端延伸从而最终取代老结构,成为前、后仔虫新的缘棘毛列(见图3A、C、E、G)。
2.2.4 背触毛 新的背触毛来自2组原基:发生早中期,第1、2、3列老背触毛中部前后分别出现一小列原基,即前、后仔虫的第1、2、3列背触毛原基(见图2F;图4F;长箭头)。这些原基逐步增殖,向前后两端延伸(见图2H)。随着原基的进一步增殖,第3列背触毛原基在末端断裂形成第4列背触毛原基(见图3B;图4K;图5G;长箭头),同时前、后仔虫右缘棘毛原基前端分离出来一小段原基,即背缘触毛原基(见图3A;图5A;长箭头)。发生中期,第1、2、4列背触毛原基的末端各形成1根尾棘毛,同时背缘触毛原基纵向分化成一长一短2列原基,即背缘触毛原基5和6(见图3C、D;图4L;图5B)。随后,背触毛原基1-4继续发育并向前后两端延伸,背缘触毛原基5、6向虫体背部迁移,老背触毛也逐渐被吸收(见图3E~H)。
2.2.5 核器 核器的发生过程与其他腹毛类纤毛虫非常相似,即在早期2枚大核的远端各出现一条DNA复制带,并向细胞的中央移动(见图2H,长箭头),至发生中后期2枚大核融合(见图3D;图5H;长箭头),后逐渐拉伸,分裂2次后随着细胞分裂分配到前、后仔虫中,从而保证2个新仔虫各有2枚大核(见图3F、H;图5I~L)。小核可见有丝分裂(见图3B、D、F、H)。
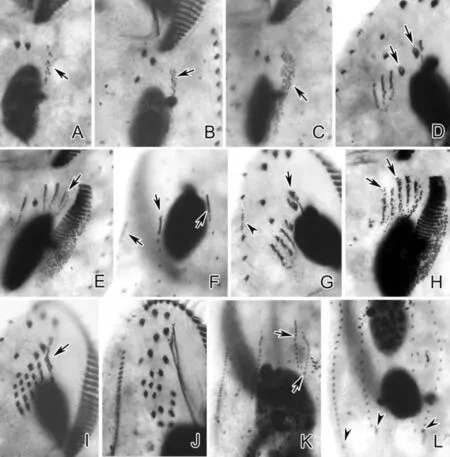
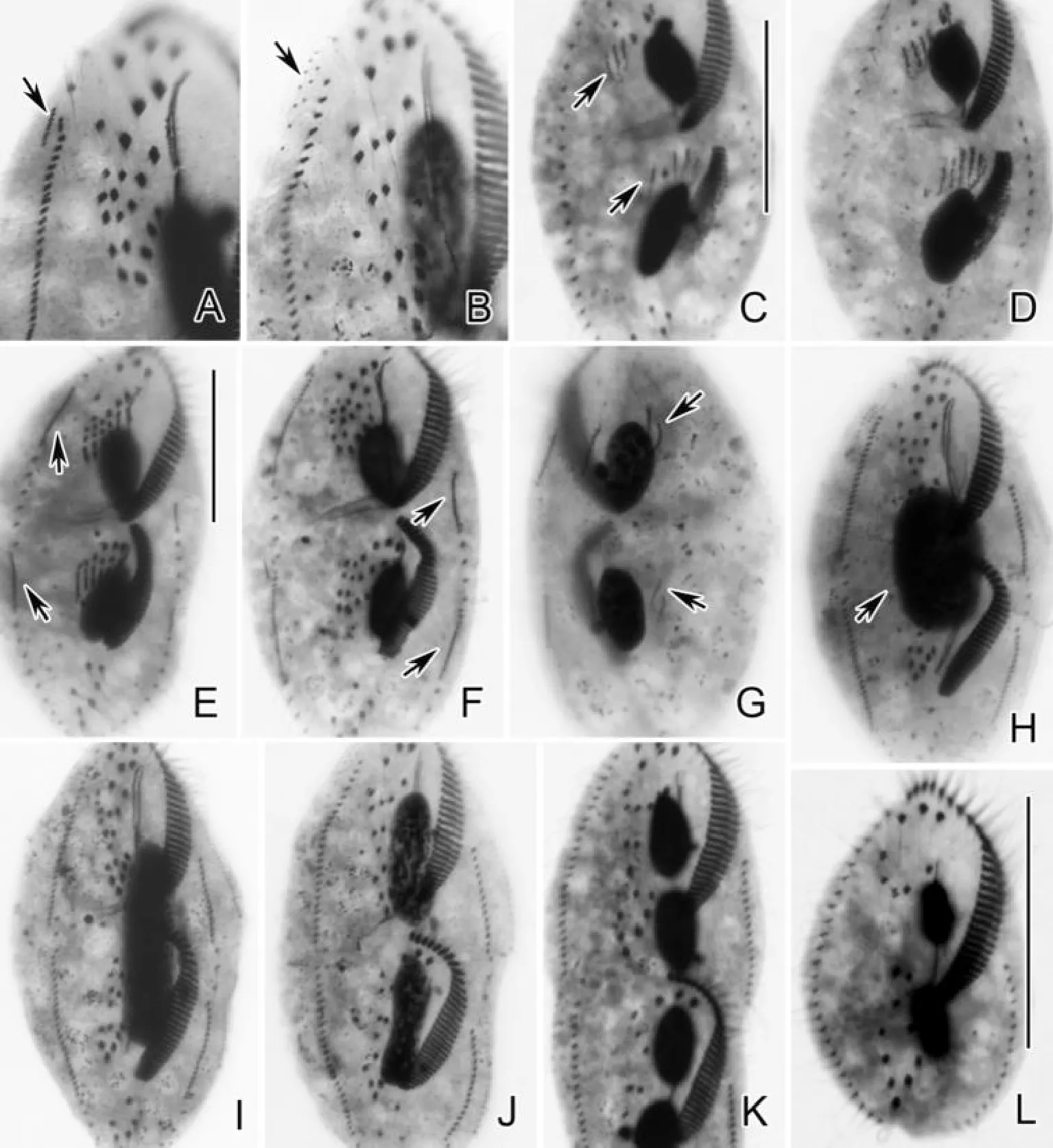
(A、B、C.早期发生个体腹面观,示后仔虫口原基(长箭头);D.前仔虫腹面观,示正在反分化的老棘毛(长箭头);E.后仔虫腹面观,示波动膜原基(长箭头);F.背面观,示背触毛原基(长箭头);G.前仔虫腹面观,示老的波动膜前端反分化的部分(长箭头)及右缘棘毛原基(短箭头);H.后仔虫腹面观,示条状额-腹-横棘毛原基(长箭头);I.前仔虫腹面观,示新形成的部分波动膜(长箭头);J.前仔虫腹面观,示新的额-腹-横棘毛;K.背面观,示第3列背触毛原基的断裂(长箭头);L.背面观,示新的尾棘毛(短箭头)。A, B, C. Ventral views of early dividers, showing oral primordium of the opisthe (arrows); D. Ventral view of proter, arrows mark the dedifferentiating parental cirri; E. Ventral view of opisthe, arrow indicates the undulating membranes anlagen; F. Dorsal view, showing dorsal kinety anlagen (arrows); G. Ventral view of proter, arrow marks the dedifferentiating part of the old undulating membranes, arrowhead indicates the right marginal row anlage; H. Ventral view of opisthe, arrows indicate the streak-like fronto-ventral-transverse cirral anlagen; I. Ventral view of proter, arrow indicates the newly formed part of undulating membranes; J. Ventral view of proter, to show the new fronto-ventral-transverse cirri; K. Dorsal view, arrows indicate the fragmentation of the 3rd dorsal kinety anlage; L. Dorsal view, arrowheads mark the new caudal cirri.)
图4 鬃异源棘尾虫细胞发生时期蛋白银染色照片细节特点
Fig.4 Detailed features of morphogenesis ofTetmemenapustulataafter protargol impregnation
(A.前仔虫腹面观,示由右缘棘毛原基向前分裂出的小片断(长箭头);B.前仔虫腹面观,示背缘触毛原基(长箭头);C、D.腹面观,示条状额-腹-横棘毛原基(见图C长箭头);E.腹面观,示右缘棘毛原基(长箭头);F、G.同一发生个体腹面及背面观,示左缘棘毛原基(见图F长箭头)及第3列背触毛原基的断裂(见图G长箭头);H.腹面观,示单一的大核融合体(长箭头);I~K.晚期发生个体腹面观,示大核的分裂及新棘毛的迁移;L.新分裂出的前仔虫腹面观。比例尺:50 μm(C,E);60 μm(L)。A. Ventral view of proter, to show the small segments split anteriorly from right marginal row anlage (arrow); B. Ventral view of proter, arrow notes the dorsomarginal kinety anlagen; C, D. Ventral views, to show the streak-like fronto-ventral-transverse cirral anlagen (arrows in C); E. Ventral view, arrows mark the right marginal row anlagen; F, G. Ventral and dorsal view of the same divider, showing the left marginal row anlagen (arrows in F) and the fragmentation of the 3rd dorsal kinety anlagen (arrows in G); H. Ventral view, noting the single mass of macronuclei (arrow); I~K. Ventral views of the late dividers, to show the division of macronuclei and migration of new cirri; L. Ventral view of newly separated proter. Scale bars=50 μm (in C, E); 60 μm (in L).)
图5 鬃异源棘尾虫细胞发生时期蛋白银染色照片
Fig.5 Morphogenesis ofTetmemenapustulataafter protargol impregnation
3 讨论
3.1 形态学比较
鬃异源棘尾虫的模式种群仅有简单的活体形态信息,漠河种群与之相比在体型、伸缩泡位置等方面基本一致,因此两者可以被认为是同种[17]。在纤毛图式特征及形态统计学数据等方面本种群与月亮湖及珠江种群非常吻合,并且3个种群活体大小变化幅度都较大:本种群(45~125) μm×(25~85) μm,月亮湖种群(48~124) μm×(26~83) μm,珠江种群(75~115) μm×(40~60) μm[11, 14]。但其它研究所涉种群个体均较大,特别是Bhatia(1936),Bürger(1905)和Roux(1901)种群个体明显较大(150~230 μm,230 μm × 96 μm,(180~220) μm×(85~105) μm)[1]。这些因生境不同而在个体大小方面产生较大变异的现象在淡水和土壤纤毛虫中是很普遍的[1, 18-19]。
3.2 发生学比较
与月亮湖种群相比,本种群发生学特征与其完全一致,进一步证实了本种具有非常稳定的细胞发生学特征[1, 11],从而表明该类特征可作为种类区分的一个指标。本种无性二分裂过程中皮层纤毛器演化所表现的特征,诸如:后仔虫口原基在细胞表面发生,前仔虫继承老口围带,额腹横棘毛以“1∶3∶3∶3∶4∶4”的方式分化形成18根棘毛,缘棘毛原基在老结构中产生,背触毛为2组发生式等是大多数尖毛科纤毛虫所共有的特征[1, 5]。但同时,本种亦表现出下列独征:1)前仔虫中波动膜为拼接式发生,即老波动膜只是其前端反分化产生新的原基,然后与未发生变化的后部老波动膜一起形成前仔虫的波动膜,这一发生特征仅在棘尾虫属(Stylonychia)内发现,与其它绝大多数腹毛类纤毛虫中的完全更新不同[1, 20-22];2)背缘触毛原基源于右缘棘毛原基的前端断裂,此为该种背缘触毛原基起源的首次发现[1, 20-22]。
本种的唯一同属种,贪食异源棘尾虫目前无发生学报道。本种与相近的贻贝棘尾虫(棘尾虫属的模式种)的发生学特征相比,主要区别在于:1)前者新口原基于口后腹棘毛和左缘棘毛列之间产生,而后者则靠近横棘毛左侧产生;2)前仔虫中,前者口棘毛(II/2)反分化形成FVTA II,FVTA IV-VI来自额腹棘毛IV/3,后者FVTA II来自后仔虫的口原基,老口棘毛(II/2)未发生变化,FVTA IV来自额腹棘毛IV/3,FVTA V-VI来自右侧口后腹棘毛V/4;3)后仔虫中,前者口后腹棘毛IV/2和V/4分别形成FVTA IV和V-VI,后者仅由口后腹棘毛V/4形成FVTA IV-VI。以上这些不同之处进一步证实了异源棘尾虫属建立的必要性。
3.3 系统地位
鬃异源棘尾虫的研究历史十分久远,且其系统地位一直不明。其最早由Müller于1786年归入Kerona属,随后的200年一直被视为棘尾虫属(Stylonychia)中的一个种[1],直至Eigner(1997)将此种转移到Clara属[9]。但随后发现属名Clara早被占用,于是Eigner(1999)用属名Tetmemena代替Clara[10]。本研究及前人的研究表明,尽管本种与棘尾虫在形态和发生学特征具有相似之处,这正好说明它们应隶属于同一科级阶元,但与此同时,也存在不少关键的不同,因此支持将其从棘尾虫属移出另建新属[9-10,14]。最近有关该种的分子系统学研究同样支持此形态分类的观点[23-24]。
4 结论
本文对鬃异源棘尾虫漠河种群的形态学及细胞发生学进行了详尽的研究,首次揭示了该种背缘触毛原基的起源,同时验证了鬃异源棘尾虫种内稳定的细胞发生学特征,并进一步证实了异源棘尾虫属建立的必要性,此外,还为本种的地理分布等研究提供了信息支持。
[1] Berger H. Monograph of the Oxytrichidae (Ciliophora, Hypotrichia) [J]. Monographiae Biologicae,1999, 78: 1-1080.
[2] Fan Y B, Zhao X L, Hu X Z, et al. Taxonomy and molecular phylogeny of two novel ciliates, with establishment of a new genus,Pseudogastrostylan. g. (Ciliophora, Hypotrichia, Oxytrichidae) [J]. European Journal of Protistology, 2015, 51: 374-385.
[3] Foissner W. Terrestrial and semiterrestrial ciliates (Protozoa, Ciliophora) from Venezuela and Galápagos [J]. Denisia, 2016, 35: 1-914.
[4] Luo X T, Gao F, Yi Z Z, et al. Taxonomy and molecular phylogeny of two new brackish hypotrichous ciliates, with the establishment of a new genus (Ciliophora, Spirotrichea) [J]. Zoological Journal of the Linnean Society, 2016,177.doi: 10.1111/zoj.12451.
[5] Shao C, Lu X T, Ma H G. A general overview of the typical 18 frontal-ventral-transverse cirri Oxytrichidae s. l. genera (Ciliophora, Hypotrichia) [J]. Journal of Ocean University of China, 2015, 14(2): 522-532.
[6] 王梅,胡晓钟,朱明壮,等. 尖毛虫科纤毛虫的细胞发生模式 [J]. 淄博学院学报, 2000, 2(2): 81-85.
Wang M, Hu X Z, Zhu M Z, et al. Morphogenetic patterns of oxytrichid ciliates [J]. Journal of Zibo University, 2000, 2(2): 81-85.
[7] Gupta R, Kamra K, Arora S, et al.Stylonychiaammermannisp. n., a new oxytrichid (Ciliophora: Hypotrichida) ciliate from the river Yamuna, Delhi, India [J]. Acta Protozoologica, 2001, 40: 75-82.
[8] Shi X B, Ammermann D.Stylonychiaharbinensissp. n., a new oxytrichid ciliate (Ciliophora, Hypotrichia) from the Heilongjiang Province, China [J]. Protistology, 2004, 3: 219-222.
药学生应加强药事法规的学习,他们应知道违法红线绝对不能踩,违法的行为必会受到严惩。药学院校应加强药学实践、药患沟通实践、药事管理等课程比重,强化人文素养和法律法规知识的教育。
[9] Eigner P. Evolution of morphogenetic processes in the Orthoamphisiellidae n. fam., Oxytrichidae, and Parakahliellidae n. fam., and their depiction using a computer method (Ciliophora, Hypotrichida) [J]. Journal of Eukaryotic Microbiology, 1997, 44: 553-573.
[10] Eigner P. Comparison of divisional morphogenesis in four morphologically different clones of the genusGonostomumand update of the natural hypotrich system (Ciliophora, Hypotrichida) [J]. European Journal of Protistology, 1999, 35: 34-48.
[11] Wirnsberger E, Foissner W, Adam H. Morphological, biometric, and morphogenetic comparison of two closely related species,StylonychiavoraxandS.pustulata(Ciliophora: Oxytrichidae) [J]. The Journal of Protozoology, 1985, 32: 261-268.
[12] 施心路, 宋微波, 史新柏. 腹毛目纤毛虫无性分裂期间的形态发生模式 [M]. ∥宋微波,徐奎栋,施新路,等. 原生动物学专论. 青岛: 青岛海洋大学出版社, 1999: 189-210.
Shi X L, Song W B, Shi X B. Morphogenetic modes of hypotrichous ciliates [M]. // Song W B, Xu K D, Shi X L, et al. Progress in Protozoology. Qingdao: Qingdao Ocean University Press, 1999: 189-210.
[13] 朱飞云,余育和,沈韫芬. 下毛目分子系统进化研究和鬃棘尾虫再描述 (纤毛动物门:下毛目:尖毛虫科) [J]. 动物分类学报,2002, 27(1): 24-32.
Zhu F Y, Yu Y H, Shen Y F. Molecular phylogenetic analysis of order Hypotrichida with redescription ofStylonychiapustulata(Ciliophora: Hypotrichida: Oxytrichidae) [J].Acta Zootaxonomica Sinica, 2002, 27(1): 24-32.
[14] Shao C, Pan X M, Jiang J M, et al. A redescription of the oxytrichidTetmemenapustulata(Müller, 1786) Eigner, 1999 and notes on morphogenesis in the marine urostylidMetaurostylopsissalinaLei et al., 2005 (Ciliophora, Hypotrichia) [J]. European Journal of Protistology, 2013, 49: 272-282.
[15] 宋微波,徐奎栋. 现代原生动物学研究的常用方法[M].∥宋微波,徐奎栋,施新路,等. 原生动物学专论. 青岛: 青岛海洋大学出版社, 1999: 9-25.
Song W B, Xu K D. Protocols in protozoological studies[M].// Song W B, Xu K D, Shi X L, et al. Progress in Protozoology. Qingdao: Qingdao Ocean University Press, 1999: 9-25.
[16] Wilbert N. Eine verbesserte Technik der Protargolimprägnation für Ciliaten [J]. Mikrokosmos, 1975, 64: 171-179.
[17] Müller O F. Animalcula infusoria fluviatilia et marina, quae detexit, sytematice descripsit et ad vivum delineari curavit [M]. Mölleri: Hauniae, 1786.
[18] Foissner W, Agatha S, Berger H. Soil ciliates (Protozoa, Ciliophora) from Namibia (Southwest Africa), with emphasis on two contrasting environments, the Etosha region and the Namib Desert [J]. Denisia, 2002, 5: 1-1459.
[19] Ricci N, Riggio D C. Cannibals ofOxytrichabifaria(Ciliata, Hypotrichida): I. A crowding dependent cell differentiation [J]. Journal of Experimental Zoology, 1984, 229: 339-347.
[20] Berger H. Monograph of the Urostyloidea (Ciliophora, Hypotricha) [J]. Monographiae Biologicae, 2006, 85: 1-1304.
[21] Berger H. Monograph of the Amphisiellidae and Trachelostylidae (Ciliophora, Hypotricha) [J]. Monographiae Biologicae, 2008, 88: 1-737.
[22] Berger H. Monograph of the Gonostomatidae and Kahliellidae (Ciliophora, Hypotricha) [J]. Monographiae Biologicae, 2011, 90: 1-741.
[23] Lu X T, Shao C, Yu Y H, et al. Reconsideration of the ‘well-known’ hypotrichous ciliatePleurotrichacurdsi(Shi et al., 2002) Gupta et al., 2003 (Ciliophora, Sporadotrichida), with notes on its morphology, morphogenesis and molecular phylogeny[J]. International Journal of Systematic and Evolutionary Microbiology, 2015, 65: 3216-3225.
[24] Shao C, Chen L Y, Pan Y, et al. Morphology and phylogenetic position of the oxytrichid ciliates,Urosomasalmastra(Dragesco and Dragesco-Kerneis, 1986) Berger, 1999 andU.karinaesinensenov. sspec. (Ciliophora, Hypotrichia) [J]. European Journal of Protistology, 2014, 50: 593-605.
责任编辑 朱宝象
Morphology and Ontogeny of Tetmemena pustulata(Müller, 1786) Eigner, 1999 from Mohe, China
LUO Xiao-Tian1, LI Li-Qiong1, 2, MA Hong-Gang1, HU Xiao-Zhong1
(1.College of Fisheries, Ocean University of China, Qingdao 266003, China; 2.School of Ocean, Yantai Academy, China Agricultural University, Yantai 264670, China)
The morphology and morphogenesis ofTetmemenapustulata(Müller, 1786) Eigner, 1999 isolated from Mohe, China, were investigated using live observation and protargol impregnation. Our populations resembled the other populations in terms of their live characters and ciliary patterns. The main morphogenetic features during division are described as follows: 1) the parental adoral zone of membranelles is kept by the proter, but the parental undulating membranes are retained after division, except for the anterior part dedifferentiating into undulating membranes anlagen; 2) the oral primordium of the opisthe developsdenovoon cell surface; 3) the parental cirri II/2 and III/2, IV/3, IV/2, and V/4 contribute to the formation of fronto-ventral-transverse cirral anlagen for the proter and opisthe, respectively; 4) the fronto-ventral-transverse cirral anlagen I~VI forms cirri in the mode of “1∶3∶3∶3∶4∶4”; 5) marginal cirri anlagen are generated within the parental marginal rows; 6) dorsal kineties are formed inOxytrichapattern, and the 5th and 6th dorsomarginal kineties originate from small segments split anteriorly from right marginal row anlagen, which is the first reveal for the origination of the dorsomarginal kineties forTetmemenapustulata.
ciliate; Hypotrichia; Sporadotrichida; Oxytrichidae; morphogenesis
国家自然科学基金项目(31430077);青岛市应用基础研究计划项目(15-12-1-1-jch)资助
2016-07-29;
2016-09-07
罗晓甜(1989-),女,博士生。E-mail: luoxiaotian512@163.com
** 通讯作者:E-mail: xiaozhonghu@ouc.edu.cn
Q959.117
A
1672-5174(2016)11-082-09
10.16441/j.cnki.hdxb.20160272
罗晓甜, 李俐琼, 马洪钢, 等. 鬃异源棘尾虫的形态学及细胞发生学研究[J]. 中国海洋大学学报(自然科学版), 2016, 46(11): 82-90.
LUO Xiao-Tian, LI Li-Qiong, MA Hong-Gang, et al. Morphology and ontogeny ofTetmemenapustulata(Müller, 1786) Eigner, 1999 from Mohe, China [J]. Periodical of Ocean University of China, 2016, 46(11): 82-90.
Supported by the National Natural Science Foundation of China (31430077); Applied Basic Research Project of Qingdao City (15-12-1-1-jch)
猜你喜欢
今日农业(2022年15期)2022-09-20
湖南电力(2021年1期)2021-04-13
植物研究(2020年6期)2020-03-05
计算机应用(2018年10期)2018-11-22
红土地(2018年7期)2018-09-26
浙江大学学报(农业与生命科学版)(2017年2期)2017-05-19
小朋友·快乐手工(2016年9期)2016-05-14
湖北农业科学(2014年6期)2014-07-02
数学大世界·小学低年级辅导版(2009年8期)2009-07-28
学生天地·小学低年级版(2009年5期)2009-06-29
