
鲢神经肽Y基因的克隆及其在禁食-恢复投喂条件下的表达特征分析
2016-11-24 05:26杨镇宇王忠梁宏伟赵金坤罗相忠邹桂伟
水生生物学报 2016年2期
杨镇宇王 丹,李 忠梁宏伟赵金坤罗相忠邹桂伟
(1. 华中农业大学水产学院, 武汉 430070; 2. 中国水产科学研究院长江水产研究所, 武汉 430223; 3. 中国水产科学研究院淡水渔业研究中心, 无锡 214081)
鲢神经肽Y基因的克隆及其在禁食-恢复投喂条件下的表达特征分析
杨镇宇1,2王 丹2,3李 忠2梁宏伟2赵金坤1,2罗相忠2邹桂伟2
(1. 华中农业大学水产学院, 武汉 430070; 2. 中国水产科学研究院长江水产研究所, 武汉 430223; 3. 中国水产科学研究院淡水渔业研究中心, 无锡 214081)
为深入研究神经肽Y对鲢摄食活动的调节作用, 研究利用同源克隆方法获得鲢NPY基因cDNA全长序列,并检测在禁食-恢复投喂条件下, NPY在脑和肝脏中的表达情况. 结果表明: 鲢NPY基因cDNA全长782 bp, 包括5′端非翻译区68 bp, 3′端非翻译区423 bp, 开放阅读框291 bp, 编码96个氨基酸; 氨基酸相似性比较和系统进化分析结果显示NPY较为保守; NPY在所检测的13个组织中均有表达, 并且在脑垂体中表达量最大; 禁食导致NPY mRNA在脑中的表达量显著上升, 恢复投喂6h后下降到基本水平, 表明NPY对鲢摄食有促进作用; 在肝脏中, 禁食前5天NPY mRNA表达量显著上升, 禁食第7天急剧下降, 恢复投喂4h后下降到基本水平; NPY mRNA在脑和肝脏中的表达具有组织差异性, 其在肝脏中的具体作用机制有待进一步研究. 研究结果为探讨NPY在鲢中的生物学功能和在遗传育种中的作用提供了理论依据。
鲢; NPY基因; 禁食; 恢复投喂; 基因表达
摄食和能量平衡的调节对于动物的生长至关重要, 不适当的摄食和能量代谢会抑制动物生长、延缓青春期、降低动物对寄生虫和疾病的抵抗力[1]。鱼类与其他脊椎动物一样, 通过中枢和外周的分泌因子来调节摄食和能量代谢[2]。神经肽Y,又称神经肽酪氨酸(Neuropeptide Y, NPY), 是由瑞典科学家Tatemoto等[3]于1982年从猪脑中分离出来的一种含36个氨基酸的单链多肽, 是中枢和外周神经系统中最丰富的神经肽。NPY被认为是哺乳动物中最能有效促进食欲的肽之一[4—6]。与哺乳动物相似, 鱼类NPY在脑中的结合区域也主要位于食欲调节区域[7], NPY对鱼类的摄食和能量代谢也起着至关重要的作用[8,9]。Lopez-Patino等[10]发现在金鱼(Carassiu sauratus)中, 侧脑室内注射NPY可显著提高其摄食量, 而加入NPY受体拮抗物可降低这种效应; 禁食能够引起NPY表达量上升。Carpio等[11]通过腹腔注射将重组的NPY蛋白注射到罗非鱼(Oreochromis niloticus)体中, 发现可以促进其摄食量和体重的增加。Peterson等[12]发现, 在斑点叉尾(Ietalurus punetaus)摄食前其脑组织NPY表达量增加, 并且脑组织中NPY表达量与饥饿时间呈正相关。
鲢(Hypophthalmichthys molitrix)是我国传统养殖的“四大家鱼”之一, 以浮游生物为食, 分布于我国各大水系[13]。鲢具有重要的经济价值和生态价值, 近十年来年产量都居淡水鱼类年产量第二[14],是我国普通百姓重要的动物性蛋白来源之一。NPY作为一种重要的摄食和能量代谢调控因子, 研究其对鲢摄食活动的调节作用, 有助为鲢新品种的选育提供辅助信息。本研究利用同源克隆方法获得鲢NPY基因cDNA全长序列, 并检测其在鲢各组织中的表达情况, 分析在饥饿-再投喂条件下在脑和肝中的表达特征, 从分子水平探讨NPY对鲢摄食的调节作用, 以期为进一步研究NPY在鲢中的生物学功能打下基础。
1 材料与方法
1.1 实验材料
禁食-恢复投喂实验 实验鱼由中国水产科学研究院长江水产研究所窑湾试验场提供。选取健康无病、规格一致的鲢330尾[平均体重(13.93 ± 1.35) g], 在养殖车间养殖桶(500 L)内暂养2周, 每天投喂苗种一号粉料(武汉正大有限公司)2次(8:00和16:00), 温度(29.5±1.2)℃。然后将实验鱼随机分为3个实验平行组, 分别在3个养殖桶(500 L)内养殖,每组110尾鱼, 禁食15d, 随后再恢复投喂15d (实验条件同暂养期间), 实验期间均使用曝气自来水, 每天清理粪便和残渣, 隔天换水1/3, 溶氧保持在7 mg/L以上, 自然光照。分别在禁食0(对照组)、1d、2d、3d、5d、7d、10d和15d; 在恢复投喂后1h、2h、4h、6h; 和恢复投喂后1d、3d、5d、7d、10d、15d取样, 为保持取样时间一致, 在每天中午12:00取样。在实验开始前, 15条鱼取各个组织(全脑、脑垂体、心、肝、肾、脾、前肠、中肠、后肠、肌肉、鳃、皮、眼)用于NPY的组织表达分析;在禁食-恢复投喂实验开始后, 每个采样点每个平行组各取5条鱼(3个实验平行组)分别取全脑和肝脏用于检测禁食-恢复投喂实验对NPY mRNA表达量的影响。所取样品均保存于-80℃备用。
实验试剂与仪器 总RNA提取试剂Trizol Reagent购自Invitrogen 公司; 反转录试剂PrimeScriptTMRT Reagen Kit with gDNA Eraser和亚克隆载体pMD®18-T Vector system购自大连宝生物工程有限公司(TaKaRa); 琼脂糖凝胶回收试剂Agarose Gel DNA Fragment Recovery Kit购自北京百泰克生物有限公司(Bioteke); 3′端和5′端的扩增试剂SMART RACE cDNAAmplication Kit购自美国Clontech公司; 荧光定量试剂QuantiFastTM SYBR®Green PCR Kit购自德国Qiagen公司; 大肠杆菌DH5α由本实验室保种。其他试剂均为国产分析纯。
荧光定量PCR仪为Qiagen公司生产的Rotor-Gene 6200 system, 紫外分光光度计为UNICO公司的UV-3802紫外分光光度计, 凝胶成像系统为Syngene公司的G: BOX。
引物 根据GenBank中其他已知物种的NPY基因cDNA序列的保守区设计引物NPY1用于扩增鲢NPY基因中间片段, 设计引物SMA NPY 3和SMA NPY 5用于分别扩增NPY基因的3′端和5′端。RT NPY 1为定量表达的特异性引物; β-actin 1(GenBank登录号: AF301605)为定量表达的内参引物。以上引物均采用Primer 5.0设计, 由武汉擎科生物技术有限公司合成。引物序列见表1。
1.2 方法
总RNA的提取与cDNA合成 称取上述所取鲢组织约100 mg (脑组织用于扩增基因全长), 使用Trizol Reagent (Invitrogen)试剂并根据其使用说明提取总RNA。使用紫外分光光度计测定RNA浓度,并根据A260/A280值判断RNA的质量; 凝胶电泳检测RNA的完整性; 以总RNA为模板, 使用PrimeScriptTM
RT Reagen Kit with gDNA Eraser (TaKaRa)试剂并按照说明书合成cDNA。

表 1 实验所用引物Tab. 1 Primers used in the experiment
3′RACE和5′RACE 按照SMART RACE cDNAAmplication Kit (Clontech) 试剂盒操作要求,采用引物SMA NPY 3、SMA NPY 5进行扩增。PCR反应体系为25 μL, 其中10×Buffer 2.5 μL, 上、下游引物 (10 μmol/L) 各1 μL, dNTPs (10 mmol/L)0.5 μL, 高保真Taq酶0.3 μL, 模板1 μL, 以ddH2O补足体积。反应程序为: 94℃ 5min; 94℃ 30s, 58℃30s, 72℃ 45s, 35个循环; 72℃ 10min。PCR产物经电泳检测、回收、连接、转化后, 挑单克隆菌落送武汉生工生物技术有限公司测序。
序列分析 利用ContigExp软件对获得的鲢NPY cDNA序列进行片段拼接和序列分析, 用DNAStar software 5.0软件对鲢NPY氨基酸序列进行排列比对分析, 用Clustal W 和 MEGA 5.0软件对鲢氨基酸序列进行多序列比较和聚类分析。
鲢NPY mRNA表达的半定量PCR检测 利用引物RT NPY 1和内参引物 β-actin 1检测NPY基因在脑、脑垂体、心脏、肝脏、脾脏、肾脏、眼睛、皮肤、鳃、前肠、中肠、后肠和肌肉等各组织中的表达情况。PCR反应体系为15 μL, 包括10×Buffer 1.5 μL, 上、下游引物 (10 μmol/L) 各0.5 μL, dNTPs (10 mmol/L) 0.3 μL, Taq酶0.2μL, 模板0.8 μL, 以ddH2O补足体积。反应程序: 94℃5min; 94℃ 30s, 58℃ 30s, 72℃ 30s, 27( β-actin)和29(NPY)个循环; 72℃ 10min。对PCR产物进行1%琼脂糖凝胶电泳, 并拍照保存。
实时荧光定量PCR分析 利用引物RT NPY 1和内参引物 β-actin 1检测NPY基因在各组织、以及在饥饿处理条件下的脑和肝脏中的表达情况。反应体系为20 μL: 2×SYBR RealtimePCR Rremixture (Qiagen, Germany)10 μL, 上、下游引物(10 μmol/L)各1 μL, cDNA模板1 μL, 加ddH2O至20 μL。反应条件: 95℃ 10min; 95℃ 15s, 58℃ 15s, 72℃ 20s, 40个循环。使用Rotor-Gene 6200 system仪自带软件建立NPY和内参 β-actin的标准曲线, NPY和 β-actin的扩增效率分别为1.00和0.98, 研究表明在鱼类禁食-恢复投喂条件下 β-actin的表达量是稳定的[15—17]。应用2-ΔΔCt分析方法对数据进行分析, 具体方法参考文献[18], 计算公式如下:
Formula=2-ΔΔCtF=2-[(待测组目的基因平均Ct值)-(待测组内参基因平均Ct值)-(对照组目的基因平均Ct值)-(对照组内参基因平均Ct值)]

2 结果
2.1 鲢NPY基因序列分析
鲢NPY基因cDNA全长782 bp (GenBank 登录号: KJ933391), 包括68 bp的5′端非翻译区、291 bp的最大开放阅读框和423 bp 的3′端非翻译区。该cDNA序列编码96个氨基酸, 在3′端非翻译区有真核细胞加尾信号(AATAAA)和Poly(A)尾巴。
2.2 鲢NPY基因的同源性分析和系统进化树构建
选取GenBank中已知的一些物种的NPY氨基酸序列: 人(Homo sapiens, NP_000896.1)、原鸡(Gallus gallus, AAA48991.1)、牛(Bos Taurus,AAR37328.1)、非洲爪蟾(Xenopus laevis, AAA499-17.1)、小鼠(Musmus culus, EDK98613.1)、斑马鱼(Danio rerio, AAI62071.1)、草鱼(Ctenopharyngodon idella, AGI44276.1)、中华倒刺鲃(Spinibarbus sinensis, ABE73783.1)、江黄颡鱼(Pelteobagrus vachellii, AEM75018.1)、鳗鲡(Anguilla japonica, AFN84517.1)、黄鳝(Monopterus albus,AEX97162.1)、欧洲鲈(Dicentrarchus labrax,CAB64932.1)、齐口裂腹鱼(Schizothorax prenanti,AGF80335.1)、大西洋鲑(Salmo salar, BAH241-01.1)。使用DNAMan softwar软件将这些物种和鲢的NPY氨基酸序列进行比对。结果表明NPY氨基酸序列在分子进化过程中较保守, 与各物种NPY有较高的同源一致性, 其中, 与草鱼、中华倒刺鲃、齐口裂腹鱼相似度最高, 分别为100%、97.9%和97.9%, 与鸟类和哺乳动物的相似度也高达60%以上。
通过邻接法(NJ法)构建的基于鲢NPY氨基酸序列的系统发育树(图 1)进一步可以看出, 鲢NPY与草鱼NPY的亲缘关系最近, 然后与鲤形目和鲶形目鱼的NPY聚在一起; 鳗鲡、大西洋鲑、黄鳝和欧洲鲈的NPY聚为一支; 人、牛、家鼠、原鸡和非洲爪蟾聚为一支。脊椎动物的NPY氨基酸序列相似性比较结果与系统进化分析的结果基本一致。
2.3 鲢NPY基因组织表达谱
分别用半定量和实时荧光定量PCR两种方法检测NPY基因在鲢脑、脑垂体、心脏、肝脏、脾脏、肾脏、眼睛、皮肤、鳃、前肠、中肠、后肠和肌肉等组织中的表达情况(图 2、图 3)。两种方法检测的结果基本一致, NPY在脑垂体、心脏、脾脏、肾脏、鳃和肝脏中大量表达, 在肠道中表达量较低, 在脑、眼睛和皮肤中仅有轻微表达, 在肌肉中几乎不表达。两种方法检测的结果存在一定的差异性, 推测是由于二者检测的灵敏度不一样造成。
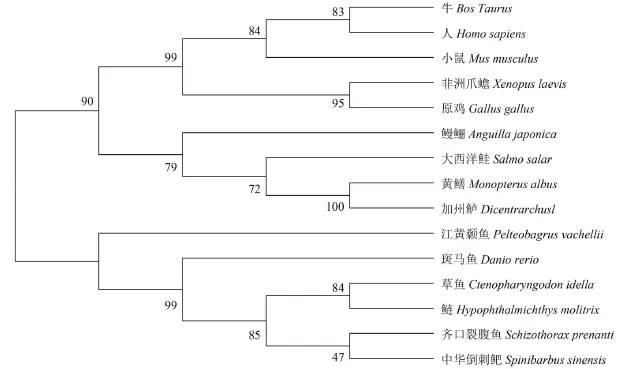
图 1 NPY的系统进化树Fig. 1 The phylogenetic tree of vertebrate NPYs
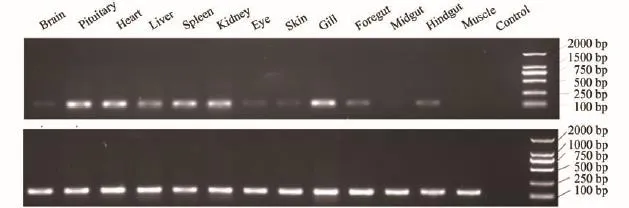
图 2 NPY在不同组织中的表达(RT-PCR检测)Fig. 2 The expression of NPY mRNA in different tissues of the silver carp (with RT-PCR)
2.4 禁食-恢复投喂对鲢脑和肝脏中NPY表达的影响
由图 4可以看出, 禁食1—5d对脑中NPY mRNA的表达没有影响; 禁食第7天, NPY表达量显著上升(P<0.05), 直到禁食第15天上升到最大(P<0.05); 恢复投喂2h后开始, NPY急剧地下降(P<0.05), 直到6h后下降到基本水平。在肝脏中(图 5), 禁食前5d,NPY mRNA表达量显著上升(P<0.05); 禁食第7天,NPY表达量急剧下降(P<0.05), 随后下降趋势变缓(P<0.05); 在恢复投喂后, NPY表达量继续下降(P<0.05), 直到恢复投喂4h后恢复到基本水平。
3 讨论
3.1 鲢NPY基因cDNA序列分析
NPY被证明是脊椎动物进化过程中最保守的神经肽之一[11,19,20]。在本研究中, 氨基酸序列相似性比较显示鲢NPY基因与各物种NPY基因的同源一致性高达60%以上, 与草鱼NPY的同源一致性更是高达100%, 表明NPY基因在长期的分子进化过程中高度保守, 由此可猜测NPY基因在各物种中的功能应该比较保守, 在鱼类中也应具有调节摄食和能量代谢的功能。与氨基酸序列相似性比较结果一致,系统进化分析显示鲢NPY与草鱼NPY亲缘关系最近, 系统进化分析结果与用传统方法所进行的物种亲缘关系分类结果一致。
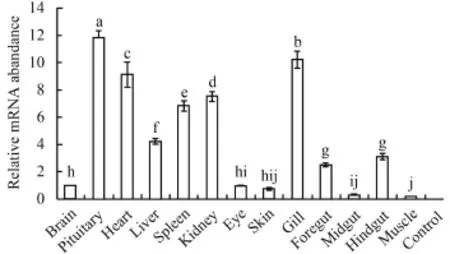
图 3 NPY在不同组织中的表达(荧光定量PCR检测)Fig. 3 The expression of NPY mRNA in different tissues of the silver carp (with real-time quantitative PCR)
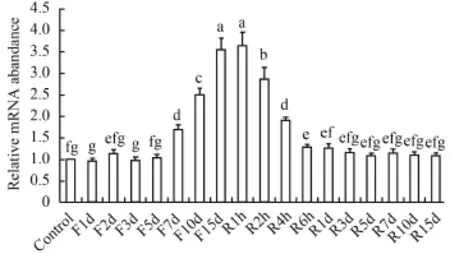
图 4 禁食-恢复投喂条件下, NPY在脑中的表达Fig. 4 The effect of fasting-refeeding on the level of NPY mRNA in the brain of the silver carp
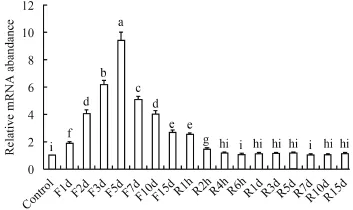
图 5 在禁食-恢复投喂条件下NPY在肝脏中的表达Fig. 5 ffect of fasting-refeeding on NPY mRNA levels in liver of silver carp
3.2 鲢NPY基因组织表达分析
NPY是一种内源性的下丘脑调节肽, 产生于下丘脑的弓状核和脑干中的离散细胞群, 主要在脑中发挥作用, 但是大量研究表明NPY不仅分布在神经系统中, 在非神经组织中同样大量存在[20—23]。在本研究中, NPY在脑垂体、心脏、脾脏、肾脏、鳃和肝脏中大量表达, 在肠道、脑、眼睛、皮肤和肌肉中少量表达, 这与已有文献报道鱼类NPY可以通过神经内分泌系统发挥中枢调节作用, 还可能以旁分泌或自分泌的方式参与鱼类外周组织功能调控[24]的观点一致。研究表明, NPY在南方鲇(Silurus meridionalis)[25]、大西洋鲑[26]、斜带石斑鱼(Epinephelus coioides)[24]、黄颡鱼[27]、大西洋鳕(Gadus morhua)[28]的脑中大量表达, 而在本研究中, NPY在脑垂体中表达量最大, 在脑中只有少量表达。推测其原因可能为: (1)上述研究者在采样过程中没有将全脑与脑垂体分成两个组织分别取样, 而是将二者一起作为脑组织处理, 而本实验采样时则将鲢的全脑和脑垂体作为两个组织分别取样; (2)鱼类的脑垂体分为神经垂体和腺垂体两部分, 神经垂体主要由下丘脑神经分泌细胞的轴突纤维组成, 起着仓库的作用, 贮存由神经纤维传送的下丘脑分泌的部分激素, 例如抗利尿激素和催产素, 当身体需要时才将这些激素释放到血液中[29]。NPY产生于下丘脑, 推测它是这类激素之一, 由下丘脑分泌但是贮存在神经垂体中, NPY在脑垂体中积累就导致NPY在脑垂体中的表达量比在脑中高。
NPY在非神经组织中的表达存在种间差异: 本研究结果显示, NPY在所检测的13种组织中都有表达; Macdonald等[30,31]发现NPY在美洲拟鲽(Pseudopleuronectes americanus)、冬鳐(Raja ocellata)的脑、肠道、肝脏、肾脏、肌肉、心脏和胃中都有表达; 而李振华[32]在大鳍鳠(Mystus macropterus)的心脏、胃、肌肉中均未检测到NPY表达; Cerdá-Reverter等[33]在海鲈(Dicentrarchus labrax)的肝脏、心脏、肾脏、肠中也未检测到NPY表达。
NPY在鲢肠道中表达, 说明与哺乳动物和一些鱼类一样, NPY可能影响鲢胃肠道的收缩和排空作用。Dumont等[34]认为NPY在哺乳动物中扮演脑肠肽的角色; Shahbazi等[35]发现NPY能导致大西洋鳕的血管舒张和肠道收缩; Bjenning等[36,37]发现NPY有抑制软骨鱼类胃收缩和胃排空的作用。
3.3 禁食-恢复投喂对鲢NPY表达的影响
下丘脑弓状核NPY神经元是一个关键的摄食中枢, 主要的外周和中枢中关于能量平衡的信号在这里感应和集成, NPY神经元诱导摄食来响应外周代谢状态[38]。在哺乳动物中, NPY可改变摄食行为,并与脂肪调节密切相关, 在调节能量平衡方面发挥着重要作用[39]。在本研究中, 在禁食-恢复投喂条件下, NPY mRNA的表达量在脑和肝脏中都存在剧烈变化, 表明与哺乳动物一样, NPY对鲢的摄食和能量代谢起着重要的调节作用。在脑中, 禁食导致NPY mRNA表达量上升, 重新投喂之后下降, 表明NPY起着促进鲢摄食的作用。值得注意的是,Narnaware等[40]发现, 经过72h禁食后, 金鱼脑NPY mRNA表达量显著上升, 随后恢复投饵3h, NPY表达量快速下降到正常水平; 曹磊等[27]研究黄颡鱼NPY发现, 禁食96h后, 黄颡鱼脑NPY mRNA表达量显著增加, 重新投喂后3h NPY的表达量即下降到正常水平; 在本研究中, 禁食前5d, NPY mRNA 的表达量一直无明显变化, 禁食第7天, NPY的表达量开始显著上升, 恢复投喂6h后即下降到正常水平; Kehoe等[28]在大西洋鳕中发现, 禁食7d其NPY mRNA表达量都没有明显变化。上述研究表明, NPY系统没有在禁食一开始就呈现出明显的调节作用, 并且在不同种类的鱼中NPY出现调节作用的禁食时间不一致, 推测这可能与各种鱼的耐饥饿能力有关,只有饥饿达到一定程度才能引起NPY系统的响应。
与脑中一样, 在禁食-再投喂条件下, 肝脏中的NPY mRNA水平也发生了剧烈变化, 禁食前5天NPY mRNA表达量显著上升, 随后急剧下降直到重新投喂4h后恢复到基本水平, 表明肝脏中的NPY系统对禁食-再投喂条件产生了响应, 也参与鲢摄食和能量代谢的调节。但是, 肝脏中NPY mRNA的表达情况与脑中的明显不一致, Sucajtys-Szulc等[41]研究慢性限食对大鼠脑和肝脏NPY mRNA的影响时也发现, 长期限食使大鼠脑NPY mRNA的表达量上升, 而肝脏NPY mRNA的表达量却下降。因此猜测脑和肝脏中的NPY参与动物摄食和能量代谢的方式可能不同, 但关于动物肝脏中的NPY的研究报道较少, NPY在动物肝脏中的具体作用并不十分清楚,NPY在鲢肝脏中的作用机制有待我们进一步研究。
[1]Matteri R L. Overview of central targets for appetite regulation [J]. Animal Science, 2001, 79: E148—E158
[2]Xie X J, Deng L, Zhang B, et al. Advances and studies on ecophysiological effects of starvation on fish [J]. Acta Hydrobiologica Sinica, 1998, 22(2): 181—188 [谢小军, 邓利, 张波, 等. 饥饿对鱼类生理生态学影响的研究进展.水生生物学报, 1998, 22(2): 181—188]
[3]Tatemoto K, Carlquist M, Mutt V. Neuropeptide Y-a novel brain peptide with structural similarities to peptide YY and pancreatic polypeptide [J]. Nature, 1982, 296(5858): 659—660
[4]Stanley B G, Kyrkouli S E, Lampert S, et al. Neuropeptide Y chronically injected into the hypothalamus: a powerful neurochemical inducer of hyperphagia and obesity [J]. Peptides, 1986, 7(6): 1189—1192
[5]Woods S C, Figlewicz D P, Madden L, et al. NPY and food intake: discrepancies in the model [J]. Regulatory Peptides, 1998, (75—76): 403—408
[6]Gehlert D R. Role of hypothalamic neuropeptide Y in feeding and obesity [J]. Neuropeptides, 1999, 33(5): 329—338
[7]Liao W Q, Liang X F, Fang L. Research of NPY gene and its appetite regulation mechanism [J]. Water Conservancy Related Fisheries, 2005, 25(4): 35—37 [廖婉琴,梁旭方, 方玲. 鱼类神经肽Y基因及其食欲调控机理研究. 水利渔业, 2005, 25(4): 35—37]
[8]Silverstein J T, Plisetskaya E M. The effects of NPY and insulin on food intake regulation in fish [J]. Integrative and Comparative Biology, 2000, 40(2): 296—308
[9]Wang Q, Tan X G, Sun W, et al. Expression of recombinant flounder NPY protein in vitro and detection of endogenous NPY in flounder [J]. Marine Sciences, 2014,38(4): 15—19 [王倩, 谭训刚, 孙威, 等. 牙鲆NPY的体外表达及体内检测. 海洋科学, 2014, 38(4): 15—19]
[10]López-Patiño M A, Guijarro A I, Isorna E, et al. Neuropeptide Y has a stimulatory action on feeding behavior in goldfish (Carassius auratus) [J]. European Journal of Pharmacology, 1999, 377(2—3): 147—153
[11]Carpio Y, Acosta J, Morales A, et al. Cloning, expression and growth promoting action of Red tilapia (Oreochromis sp.) neuropeptide Y [J]. Peptides, 2006, 27(4): 710—718
[12]Peterson B C, Waldbieser G C, Riley L G, et al. Pre-and postprandial changes in orexigenic and anorexigenic factors in channel catfish (Ictalurus punctatus) [J]. General and Comparative Endocrinology, 2012, 176(2): 231—239
[13]Zhang S M, Deng H, Wang D Q, et al. Population structure and genetic diversity of silver carp and grass carp from populations of Yangtze River system revealed by RAPD [J]. Acta Hydrobiologica Sinica, 2001, 25(4): 324—330 [张四明, 邓怀, 汪登强, 等. 长江水系鲢和草鱼遗传结构及变异性的RAPD研究. 水生生物学报,2001, 25(4): 324—330]
[14]China Fishery Statistical Yearbook [M]. Beijing: Chinese Agriculture Press. 2004—2014 [中国渔业统计年鉴. 北京: 中国农业出版社. 2004—2014]
[15]Huising M O, Geven E J W, Kruiswijk C P, et al. Increased leptin expression in common carp (Cyprinus carpio) after food intake but not after fasting or feeding to satiation [J]. Endocrinology, 2006, 147(12): 5786—5797
[16]Lindblom J, Haitina T, Fredriksson R, et al. Differential regulation of nuclear receptors, neuropeptides and peptide hormones in the hypothalamus and pituitary of food restricted rats [J]. Molecular Brain Research, 2005,133(1): 37—46
[17]Tanic N, Perovic M, Mladenovic A, et al. Effects of aging, dietary restriction and glucocorticoid treatment on housekeeping gene expression in rat cortex and hippocampus — evaluation by real time RT-PCR [J]. Journal of Molecular Neuroscience, 2007, 32(1): 38—46
[18]Livak K J, Schmittgen T D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCTmethod [J]. Methods, 2001, 25(4): 402—408
[19]Mattews J E, Jansen M, Lyerly D, et al. Pharmacological characterization and selectivity of the NPY antagonist GR231118 (1229U91) for different NPY receptors [J]. Regulatory Peptides, 1997, 72(2—3): 113—119
[20]Yu Y, Liang X F, Li S Y, et al. Molecular cloning and evolutional analysis of NPY, UCP2, LPL and HL gene of Largemouth Bass (Micropterus salmoides) [J]. Acta Hydrobiologica Sinica, 2008, 32(6): 900—907 [于燕, 梁旭方, 李诗盈, 等. 大口黑鲈脂代谢相关基因NPY, UCP2,LPL, HL克隆与分子进化分析. 水生生物学报, 2008,32(6): 900—907]
[21]Aboumder R, Elhusseiny A, Cohen Z, et al. Expression of neuropeptide Y receptors mRNA and protein in human brain vessels and cerebromicrovascular cells in culture[J]. Journal of Cerebral Blood Flow and Metabolism,1999, 19(2): 155—163
[22]Heilig M, Widerlöv E. Neuropeptide Y: an overview of central distribution, functional aspects, and possible involvement in neuropsychiatric illnesses [J]. Acta Psychiatrica Scandinavica, 1990, 82(2): 95—114
[23]Doyon C, Gilmour K M, Trudeau V L, et al. Corticotropin-releasing factor and neuropeptide Y mRNA levels are elevated in the preoptic area of socially subordinate rainbow trout [J]. General and Comparative Endocrinology,2003, 133(2): 260—271
[24]Chen R. Cloning, prokaryotic expression and functions of NPY in orange-spotted grouper (Epinephelus coioides)[D]. Thesis for master of Science. Sun Yat-Sen University, Guangzhou. 2006 [陈蓉. 斜带石斑鱼神经肽Y基因的克隆、原核表达与功能研究. 硕士学位论文, 中山大学, 广州. 2006]
[25]Yang Y, Peng H W, Li Y L, et al. Molecular cloning and tissue expression analysis of the NPY gene in Silurus meridionalis [J]. Journal of Henan University (Natural Science), 2013, 43(3): 295—300 [杨雁, 彭焕文, 李艳利, 等.南方鲇 (Silurus meridionalis) 神经肽Y基因cDNA克隆及组织表达分析. 河南大学学报 (自然科学版), 2013,43(3): 295—300]
[26]Murashita K, Kurokawa T, Ebbesson L O E, et al. Characterization, tissue distribution, and regulation of agoutirelated protein (AgRP), cocaine-and amphetamine-regulated transcript (CART) and neuropeptide Y (NPY) in Atlantic salmon (Salmo salar) [J]. General and Comparative Endocrinology, 2009, 162(2): 160—171
[27]Cao L, Liang H W, Li Z, et al. Cloning and expression analysis of NPY gene in yellow catfish (Pelteobagrus fulvidraco) [J]. Journal of Northwest A and F University(Natural Science Edition), 2013, 41(7): 1—7 [曹磊, 梁宏伟, 李忠, 等. 黄颡鱼神经肽Y基因 (NPY) cDNA全序列的克隆及其表达特征分析. 西北农林科技大学学报 (自然科学版), 2013, 41(7): 1—7]
[28]Kehoe A S, Volkoff H. Cloning and characterization of neuropeptide Y (NPY) and cocaine and amphetamine regulated transcript (CART) in Atlantic cod (Gadus morhua) [J]. Comparative Biochemistry and Physiology Part A: Molecular and Integrative Physiology, 2007,146(3): 451—461
[29]Liu Z F. The progress of checking methods on pituitary disease [J]. Introduction of Japanese Medical Science,1995, 16(2): 88—89 [刘志凡. 垂体疾病检查方法的进展. 日本医学介绍, 1995, 16(2): 88—89]
[30]Macdonald E, Volkoff H. Cloning, distribution and effects of season and nutritional status on the expression of neuropeptide Y (NPY), cocaine and amphetamine regulated transcript (CART) and cholecystokinin (CCK) in winter flounder (Pseudopleuronectes americanus) [J]. Hormones and Behavior, 2009, 56(1): 58—65
[31]Macdonald E, Volkoff H. Neuropeptide Y (NPY), cocaine and amphetamine-regulated transcript (CART) and cholecystokinin (CCK) in winter skate (Raja ocellata): cDNA cloning, tissue distribution and mRNA expression responses to fasting [J]. General and Comparative Endocrinology, 2009, 161(2): 252—261
[32]Li Z H. The cDNA cloning and characterization analysis of NPY gene in Mystus macropterus Bleeker [D]. Thesis for master of Science. School of Life Science Southwest University, Chongqing. 2011 [李振华. 大鳍鳠NPY基因cDNA克隆及特征分析. 硕士学位论文, 西南大学生命科学学院, 重庆. 2011]
[33]Cerdá-Reverter J M, Martı́nez-Rodrı́guez G, Zanuy S, et al. Cloning the neuropeptide Y exon 2 from sea bass(Dicentrarchus labrax) [J]. Comparative Biochemistry and Physiology Part B: Biochemistry and Molecular Biology, 1999, 123(2): 181—186
[34]Dumont Y, Martel J C, Fournier A, et al. Neuropeptide Y and neuropeptide Y receptor subtypes in brain and peripheral tissues [J]. Progress in Neurobiology, 1992, 38(2): 125—167
[35]Shahbazi F, Holmgren S, Larhammar D, et al. Neuropeptide Y effects on vasorelaxation and intestinal contraction in the Atlantic cod Gadus morhua [J]. American Journal of Physiology, 2002, 282(5): 1414—1421
[36]Bjenning C, Hazon N, Balasubramaniam A, et al. Distribution and activity of dogfish NPY and Peptide YY in the cardiovascular system of the common dogfish [J]. American Journal of Physiology, 1993, 264(6): 1119—1124
[37]Bjenning C, Holmgren S, Farrell A P. Neuropeptide Y potentiates contractile response to norepinephrine in skate coronary artery [J]. American Journal of Physiology,1993, 265(2): 661—665
[38]Kohno D, Yada T. Arcuate NPY neurons sense and integrate peripheral metabolic signals to control feeding [J]. Neruopeptides, 2012, 46(6): 315—319
[39]Wang J, Wang Y J. Influence of different diet on NPY expression in arcuate nucleus of rats hypothalamus [J]. Journal of Chengde Medical College, 2012, 29(3): 241—243 [王晶, 王英杰. 不同膳食对大鼠下丘脑弓状核NPY表达的影响. 承德医学院学报, 2012, 29(3): 241—243]
[40]Narnaware Y K, Peter R E. Effects of food deprivation and refeeding on neuropeptide Y (NPY) mRNA levels in goldfish [J]. Comparative Biochemistry and Physiology Part B: Biochemistry and Molecular Biology, 2001,129(2—3): 633—637
[41]Sucajtys-Szulc E, Goykea E, Korczynska J, et al. Chronic food restriction differentially affects NPY mRNA level in neurons of the hypothalamus and in neurons that innervate liver [J]. Neuroscience Letters, 2008, 433(3): 174—177
CLONING AND EXPRESSION OF NPY IN THE SILVER CARP(HYPOPHTHALMICHTHYS MOLITRIX) DURING FASTING AND REFEEDING
YANG Zhen-Yu1,2, WANG Dan2,3, LI Zhong2, LIANG Hong-Wei2, ZHAO Jin-Kun1,2,LUO Xiang-Zhong2and ZOU Gui-Wei2
(1. College of Fisheries, Huazhong Agricultural University, Wuhan 430070, China; 2. Yangtze River Fisheries Research Institute,Chinese Academy of Fisheries Sciences, Wuhan 430223, China; 3. Freshwater Fisheries Research Center of Chinese Academy of Fishery Sciences, Wuxi 214081, China)
NPY plays a significant role in regulating food intake and energy metabolism in teleosts. To understand the biological role of NPY in the silver carp (Hypophthalmichthys molitrix), in this study we determined the full-length cDNA sequence of NPY and investigated the effect of fasting and refeeding on the level of NPY mRNA in the brain and the liver. The full cDNA sequence of NPY consisted of 782 bp, including a 291 bp open reading frame that potentially encodes 96 amino acids, a 68 bp 5′-untranslated region and a 423 bp 3′-untranslated region. Our study on the similarity of the amino acid sequence and the phylogenetic analysis of NPY demonstrated that the NPY gene was relatively conservative during the molecular evolution. The mRNA of NPY was ubiquitously expressed in all the tested tissues and especially abundant in the pituitary. In the brain, the expression of NPY mRNA increased significantly during fasting,and then decreased to the basal level at 6h after refeeding, which indicated that NPY could boost the food intake of the silver carp. In the liver, the expression of NPY mRNA increased significantly until 5d after fasting, then decreased sharply on the 7thday, and finally
to the basal level at 4h after refeeding. NPY was expressed in a tissue-specific pattern in both the brain and the liver. However, the function of NPY in the liver needs further study. Our study provided insights into the biological roles of NPY in the silver carp.
Silver carp; NPY gene; Fasting; Refeeding; Gene expression analysis
10.7541/2016.33
Q344+.1
A
1000-3207(2016)02-0235-08
2015-02-05;
2015-05-20
现代农业产业技术体系建设专项资金(CARS-46-01); “十二五”科技支撑计划(2012BAD26B02); 中央级公益性科研院所基本科研业务费专项资金(2015JBFM39); 农业部热带亚热带水产资源利用与养殖重点实验室开放课题; 中央高校基本科研业务费专项基金资助(2013SC14)资助 [Supported by the earmarked fund for China Agriculture Research System (No. CARS-46-01); National Science and Technology Support Program (No. 2012BAD26B02); National Nonprofit Institute Research Grant of Freshwater Fisheries Research Center (No. 2015JBFM39); Key Laboratory of Tropical& Subtropical Fishery Resource Application and Cultivation, Ministry of Agriculture; Fundamental Research Funds for the Central Universities of China (No. 2013SC14)]
杨镇宇(1989—), 女, 四川宜宾人; 硕士研究生; 主要从事鱼类遗传育种研究。E-mail: yzhygood@163.com
邹桂伟, E-mail: zougw@yfi.ac.cn
猜你喜欢
医学综述(2022年5期)2022-04-20
中国药学药品知识仓库(2022年2期)2022-03-23
实用癌症杂志(2021年9期)2021-10-13
海洋科学(2021年6期)2021-08-02
现代临床医学(2021年2期)2021-03-29
医学与法学(2020年3期)2020-09-18
科学(2020年2期)2020-08-24
科学(2020年2期)2020-01-08
黄河黄土黄种人(2018年8期)2018-11-09
生物工程学报(2017年7期)2017-08-01
