
皮肤中基底膜的结构与功能
2016-11-29 11:32张丽丽李贤玉范春晖综述北垣雅人惠美子审校
中国美容医学 2016年10期
张丽丽 李贤玉 穆 荣 范春晖 综述,北垣雅人 高 须 惠美子 审校
[ 资生堂(中国)研究开发中心有限公司 北京 100176 ]
·综述·
皮肤中基底膜的结构与功能
张丽丽 李贤玉 穆 荣 范春晖 综述,北垣雅人 高 须 惠美子 审校
[ 资生堂(中国)研究开发中心有限公司 北京 100176 ]
基底膜是一类细胞外基质,存在于人体中所有的器官组织中,承担着重要的功能。皮肤的基底膜是表皮与真皮间重要的承接结构,主要成分有Ⅳ型和Ⅶ型胶原蛋白、层粘连蛋白、巢蛋白、串珠素等,通过它们交互形成的网状结构,构成了基底膜丰富多样的生物学功能,主要为表皮-真皮的连接功能、信号传导功能及渗透屏障功能等。健康的基底膜发挥多样生物学功能,使表皮-基底膜-真皮三者之间能够顺畅循环,保持皮肤的健康完整性。暴露于阳光下可引起基底膜的改变,可以引发多种肌肤烦恼,例如晒伤、光老化、皮肤癌等。研究发现一些植物提取物可以保护基底膜,如何运用这些活性成分对研究开发功能性化妆品可以提供更多的发展途径。
皮肤;基底膜;胶原蛋白;层粘连蛋白;多样生物学功能;光老化;活性植物提取物
基底膜(Basement Membrane)存在于动物上皮细胞与基质细胞之间,是厚度为50~100nm、一种柔软、坚韧的网膜结构的细胞外基质。皮肤基底膜将表皮基底细胞和位于其下层相连接的组织分隔开,向细胞提供结构支撑,并通过表皮-真皮间细胞因子调节,维持表皮-真皮间机能。基底膜同时还具有一定的渗透屏障作用。随着年龄增长及紫外线照射可引起皮肤基底膜的重叠与断裂,其与表皮、真皮之间的连接、信息传递受到阻碍,可引发多种肌肤烦恼,甚至皮肤病变。因此表皮-基底膜-真皮三者之间的顺畅循环沟通,是构成健康肌肤的重要因素,可以将其称之为“黄金循环”。
引起基底膜变化的因素多种多样,光老化是其中一个重要因素。由UV介导的光老化主要通过引起基底膜中主要成分水平的变化,导致基底膜功能的改变,从而影响表皮和真皮功能,造成三者之间“黄金循环”的不顺畅,引发各种肌肤问题。从化妆品研发的角度出发,针对基底膜的特性,寻找一些适合作用于基底膜的活性功能成分,是开发功能型化妆品的有效探索。
1 皮肤中基底膜的结构与功能
1.1皮肤中基底膜的结构:皮肤中基底膜是真皮与表皮之间的重要连接结构,经电子显微镜观察,由外至内由胞膜层、透明层、致密层和致密下层四层组成。皮肤基底膜的主要成分为Ⅳ型胶原蛋白(type Ⅳ collagen)和Ⅶ型胶原蛋白(type Ⅶ collagen)、层粘连蛋白(laminin)、巢蛋白(nidogen)/内功素(entactin)、串珠素(perlecan)等。透明层中存在层粘连蛋白,蛋白多糖,硫酸肝素等。致密层中存在Ⅳ型胶原蛋白和层粘连蛋白,两者形成网状结构,这种网状结构对于基底膜的稳定性至关重要。XVII型胶原蛋白(BP180,BPAG2)贯通透明层与半桥粒结合于致密层,致密下层中存在VII型胶原蛋白。而巢蛋白/内功素和串珠素在层粘连蛋白和Ⅳ型胶原蛋网状结构上架桥,提升基底膜的稳定性,并影响基底膜的结构完整性(见图1)。
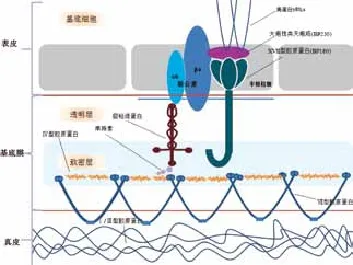
图1 基底膜结构示意图
1.2皮肤基底膜的主要成分
1.2.1Ⅳ型胶原蛋白 (Type Ⅳ collagen):基底膜组成中占50%以上的Ⅳ型胶原蛋白绝大部分都分布于致密层。不同于真皮中大量存在的Ⅰ型、Ⅲ型和Ⅴ型胶原蛋白,Ⅳ型胶原蛋白是一种非纤维状胶原蛋白,具有球状或杆状的外形,和非胶原域(NC domain)[2]。Ⅳ型胶原分子是由三条α链组成的三螺旋结构的大分子肽链构成,至今已发现了6 种同源的α链(α1~α6) , 约含1700 个氨基酸残基, 由高度同源的6 个不同基因(COL4A1~COL4A6) 分别编码[3-4]。由Ⅳ型胶原蛋白分子之间交联形成的连续三维网格具有高度的稳定性,是基底膜的重要支持结构。
1.2.2层粘连蛋白(Laminin):层粘连蛋白是基底膜中含量最丰富的非胶原蛋白,基本分布于透明层。层粘连蛋白家族(~400~800kDa)是由α、β、γ这3条多肽链组成的异三聚体[5]。
皮肤基底膜中发现的为层粘连蛋白1、层粘连蛋白5和层粘连蛋白6。层粘连蛋白5(α3β3γ2,又称为层粘连蛋白332)主要分布于透明层,跨膜半桥粒上的整合素α6β4是层粘连蛋白5的受体。其和Ⅶ型胶原蛋白结合构成锚丝(anchoring filament),穿过透明层、致密下层直接插入真皮的乳头层,固定真皮层;而且还与层粘连蛋白6或层粘连蛋白7形成共价复合物,在基底膜中该复合物(laminin 5~6/7)经巢蛋白可直接与Ⅳ型胶原蛋白反应。因此层粘连蛋白5的存在是稳定表皮-真皮连接稳定性的主要因素[6-7]。
1.2.3巢蛋白(Nidogen)/内功素(Entactin):巢蛋白-1和巢蛋白-2又称内功素(entactin),是一类同源的糖蛋白,占整个基底膜中蛋白的2~3%。它们具有三个球状域和两个相间的棒状域,与细胞外间质的各种成分结合。其中很重要的一个反应就是依次通过层粘连蛋白的LE域和巢蛋白的羰基的球状端,与层粘连蛋白γ亚单元的短臂端结合。如上述,巢蛋白帮助层粘连蛋白与Ⅳ型胶原蛋白结合从而稳定基底膜[8]。
1.2.4串珠素(Perlecan):串珠素是一类硫酸类肝素蛋白多糖,由核心蛋白和多条硫酸肝素(HS)侧链组成,是基底膜中重要的结构性组分。核心蛋白分子量约为400kDa、不同组织细胞来源的串珠素硫酸肝素侧链结构差别较大, 侧链的变化对于其生物学活性具有明显的影响。串珠素的硫酸肝素侧链 带有负电, 通过其吸水性增加组织的容积, 可以结合、释放生长因子, 使生长因子暂时失活,并在必要时恢复其活性, 具有生长因子储存库的作用。并且是细胞粘附分子的重要配体[9]。除细胞外基质外,硫酸肝素侧链也通过稳定半桥粒成分和致密层之间的相互反应,促进半桥粒的形成。因此在皮肤基底膜中对于起固定真皮作用的锚复合物的聚集,尤其是半桥粒和锚原纤维来说,硫酸肝素侧链有着非常重要的作用[10]。
1.2.5Ⅶ型胶原蛋白:Ⅶ型胶原蛋白主要分布于致密下层,锚原纤维是经二硫键结合的Ⅶ型胶原蛋的稳定二聚体。Ⅶ型胶原蛋白通过巢蛋白与层粘连蛋白5~6/7复合物结合,形成胶原蛋白网,Ⅶ型胶原蛋白通过与Ⅳ型胶原蛋白的相互反应与基底膜结合[11]。
1.2.6XVII型胶原蛋白:XVII型胶原蛋白(BP180,BPAG2)是表达于表皮角质形成细胞上的一种跨膜糖蛋白,是表皮锚复合物中一个重要的跨膜成分。它含有一个位于半桥粒斑的N-端球状头部区域与一个插入基底层的C-端胶原尾,该结构可以促进胞外配体和细胞内连接体与细胞骨架的相互反应[12]。XVII型胶原蛋白作为细胞间质的粘合分子,通过配体与基底膜中的重要成分-层粘连蛋白5和Ⅳ型胶原蛋白结合,促进表皮与真皮间的稳固结合。
1.3皮肤基底膜的主要功能
1.3.1真皮与表皮的连接支持作用:基底膜对于表皮和真皮的连接起到承上启下的重要作用。基底膜靠近表皮的一侧,上部基底层的半桥粒(hemidesmosome)是基底层细胞与下方基底膜之间的主要结构[13]。位于半桥粒的整合素α6β4的β4亚基与XVII型胶原蛋白(BP180)位于半桥粒中的N-端球状头部区域相互反应,通过XVII型胶原蛋白和BP230(230kDa大疱性类天疱疮)与表皮中的角蛋白丝(由角蛋白5和角蛋白14构成)连接,而XVII型胶原蛋白的另一个C-端插入致密层中与层粘连蛋白5相结合;与此同时,整合素α6β4与层粘连蛋白5连接,层粘连蛋白5再经巢蛋白或串珠素与Ⅳ型胶原蛋白连接,这样形成的层粘连蛋白5-巢蛋白-Ⅳ型胶原蛋白的中心轴链,将表皮层和基底膜紧密牢固地结合起来[14]。在基底膜靠近真皮的一侧,层粘连蛋白5的β3或γ2亚基与Ⅶ型胶原蛋白结合,构成半圆型锚丝,穿过透明层、致密下层直接插入真皮的乳头层,与Ⅰ型胶原蛋白构成的胶原束紧紧相连,将基底膜和真皮层紧密地固定在一起[11]。
由此可见,基底膜类似一层双面胶,将表皮层和真皮层紧密地结合在一起,而且其所含的层粘连蛋白5和Ⅳ型胶原蛋白分别各自聚集形成的网络相互交汇,对表皮层和真皮层起到了很好的结构支撑作用。
1.3.2信号传导作用:表皮和真皮不是各自独立发挥功能,其正常的动态平衡需要在这两层中来回不间断进行信号传导。总体来说这些信号分子均为小分子,可以无障碍地穿过基底膜。基底膜中的成分选择性地促进或抑制这些信号的传导。在某些情况下,信号分子储存于基底膜中,仅当基底膜受损或被破坏时释放出来。因此通过基底膜-表皮-真皮之间的沟通相当重要[15]。
1.3.3渗透屏障功能:基底膜还具有一定的渗透和屏障功能。表皮中无血管,血液中的营养物质即通过基底膜进入表皮,而表皮的细胞产物又可通过基底膜进入真皮进行代谢。一般情况下,基底膜限制分子量40 000以上的大分子通过,但当其发生损伤时,炎症因子、肿瘤因子及其他大分子物质均可以通过基底膜进入表皮[16]。
2 基底膜与皮肤老化
2.1自然老化:由于年龄增长所致的皮肤老化是客观自然规律不可抗拒的过程,临床特征表现为皮肤出现细纹、松弛、粗糙等。组织学特征表现为表皮变薄、纹理不明显。真皮胶原纤维束变直、交织排列疏松,弹力纤维变粗,弹力纤维崩解成短小的微丝。毛囊数目减少,小汗腺数目减少,大小汗腺内脂褐质沉积增多。皮脂腺增生,真皮乳头层血管约减少30%, 皮下脂肪出现皱缩。
9月3日中国尿素出口价格指数(CNEI)为 1847.26点,环比下跌25.47点;比基期下跌11.74点,跌幅为0.63%。
老化皮肤的病理特征表现为,基底膜中串珠素的硫酸肝素侧链转录水平降低,引起基底层角质形成细胞和真皮中成纤维细胞更新减少,导致表皮角化过程趋于缓慢,真皮中胶原蛋白产生能力降低,作为维持皮肤张力和承受拉力的Ⅰ型胶原蛋白逐渐减少,其构成的交联网变稀疏,Ⅲ型胶原蛋白的表达水平也同时降低[17]。
2.2光老化:日光中的紫外线与皮肤老化有密不可分的关系,称之为光老化。几乎我们能想到的所有肌肤问题,都与光老化有关,紫外线对于皮肤的伤害是累积的。即,每接受一次没有防护的日光照射,就向衰老又迈进了一步。
2.2.1光老化引起的基底膜变化:Amano等人发现,由UV介导的光老化过程中,基底膜中所含成分均有不同程度的变化。例如基底膜主要成分-Ⅳ型胶原蛋白的减少,锚丝主要成分-层粘连蛋白5的减少,串珠素的HS侧链的降解[20]。
有很多研究者对引起这些变化的因素进行了详细的探索和研究。其中Ⅳ型胶原蛋白的减少被认为是与基质金属蛋白酶(以下称为MMPs)相关。根据MMPs酶的构造及功能,通常可分为胶原酶,明胶酶,基质溶解素等。其中明胶酶包含MMP-2 (72kDa)、MMP-9 (92kDa) 等。众所周知这种MMP-2, 9可以分解作为基底膜成分的IV型胶原蛋白、层粘连蛋白、真皮基质成分的弹性蛋白等[18-20]。与之前使用的创伤性检测不同,李贤玉等人利用涂有可与白明胶-SH反应的聚酯纤维胶片,可简单快捷且非创伤性地检测角质层中白明胶酶的含量,调查表皮层中白明胶酶随着紫外线强度变化而产生的变化。同时在人体使用试验中使用该方法判断配合有白明胶酶抑制剂的护肤品对该酶的抑制状况。随着紫外线照射程度的增强和频率的提高,MMP2、MMP9的表达水平增高,导致基底膜中Ⅳ型胶原蛋白含量降低[21]。
层粘连蛋白5是血纤维蛋白溶酶的候选靶点。UVB的照射增强尿激酶型血纤维蛋白溶解原的催化剂(uPA)的合成,提高体内血纤维蛋白溶解原的水平。血纤维蛋白溶酶可以降解层粘连蛋白5的α3和β3支链,直接切断α3链中氨基和羰基端,降低角质形成细胞与层粘连蛋白5之间连接性;同时β3链中氨基端片段被剪切,减弱层粘连蛋白5与Ⅶ型胶原蛋白的亲和性[22-23]。
经UVB照射后皮肤中肝素酶的水平增高,同时角质形成细胞中肝素酶被激活,串珠素的HS侧链被显著降解[24]。Iriyama等人在肌肤等效模型中加入MMP抑制剂CGS27023A和肝素酶抑制剂BIPBIU进行培养后,经免疫组织化学分析和投射电镜观察结果表明,与未添加肝素酶的样本和空白对照相比,添加了肝素酶抑制剂的样本中BP180被极化至基底层一侧,保护了HS侧链[10]。
由上述可知,作为重要的中间结构支撑和物质、信号传导的媒介,基底膜受损后可导致表皮-真皮失去正常的结构连接,表皮无法得到来自真皮提供的营养物质及水分的输入,代谢产物也无法正常排出,其作为皮肤最外层的保护屏障功能被削弱,外界刺激容易进入肌肤内部,进一步影响真皮的状态,导致恶性循环。因此基底膜的健康与否,直接影响表皮与真皮的状态。如果三者之间维持着良好的循环状态,就可以提升整体肌肤的健康完整。这种良好状态,可称之为“黄金循环”状态。
3 保护基底膜的活性成分
研究发现很多植物提取物通过直接或间接作用于基底膜中的主要成分,维持基底膜的结构稳定,达到保护基底膜的作用。一部分植物提取物直接促进生成基底膜中胶原蛋白等主要成分,例如山毛榉芽提取物。该植物提取物可浓度依存性显著促进Ⅳ型和Ⅶ型胶原蛋白的生成,从而使表皮下部致密层连续完整和促进生成半桥粒,与此同时也可促进真皮中Ⅰ型胶原纤维的形成[27]。
还有一部分植物提取物通过抑制基底膜主要成分分解酶的活化,间接保护基底膜。例如无患子果皮提取物,可通过抑制硫酸肝素酶的活性,控制硫酸肝素分解,维持基底膜完整性,对促进基底膜微观结构形成和维持表皮分化非常重要[28]。
4 展望
基底膜是表皮与真皮之间的重要组成部分,承担固定、连接、信息传导及营养传递代谢等诸多功能。国内外已有很多专家学者对基底膜的组成、功能及各组成成分的主要作用机制进行了很多研究分析,但是针对基底膜的信号传导机制尚未十分明确,有待今后进一步的研究。我们希望通过对基底膜组成和功能的详细分析,找到更多适合作用于基底膜的活性功能成分,并将其运用至化妆品中,为功能性化妆品的开发研制提供更多的发展途径。
[1]朱学骏.皮肤基底膜带的研究现状[J].中华皮肤科杂志,2001,34 (4)∶312-313.
[2]Valerie S.Lebleu,Brian Macdonald,Raghu Kalluri.Structure and Function of Basement Membranes[J].Exp Biol Med,2007,232(9)∶1121-1129.
[3]王云峰,丁洁.Ⅳ型胶原分子结构的研究与Alport综合征[J].继续医学教育,2004,13(1)∶71-74.
[4]龚伟,曾彩虹.Goodpasture综合征和Alport综合征的分子基础-Ⅳ型胶原家族[J].肾脏病与透析肾移植杂志,2005,14(6)∶545-549.
[5]Aumailler M,Brucker-Tuderman L,Carter WG,Deutzmann R,et al.A simplified laminin nomenclature[J].Matrix Biol,2005,24(5)∶326-332.
[6]Lawrence S.Chan.Human Skin Basement Membrane in Health and in Autoimmune Diseases[J].Front Biosci,1997,2∶343-352.
[7]Nishiyama T,Amano S,Tsunenaga M,et al.The Importance of Laminin 5 in the Dermal-Epidermal basement membrane[J].J Dermatol Sci,2000,25∶51-59.
[8]Peter D.Yurchenco,Basement Membranes.Cell Scaffolding and Signaling Platform[J].Cold Spring Harb Perspect Biol,2011,3(2)∶1-27. [9]Peter D.Yurchenco,Bruce L.Patton.Developmental and Pathogenic Mechanisms of Basement Membrane Assembly[J].Curr Pharm Des,2009,15(12)∶1277-1294.
[10]S.Iriyama,M.Tsunenaga,S.Amano,Key Role of Heparan Sulfate Chains in Assembly of Anchoring Complex at the Dermal-epidermal Junction[J].Exp Dermatol,2011,20(11)∶953-955.
[11]Patricia Rousselle,Douglas R.Keene,Florence Ruggiero,et al.Laminin 5 Binds the NC-1 Domain of Type Ⅶ Collagen[J].J Cell Biol,1997,138(3)∶719-728.
[12]F.Van den Bergh, Eliason SL, Giudice GJ.Type XVII Collagen (BP180) Can Function as a Cell-matrix Adhesion Molecular via Binding to Laminin 332[J].Matrix Biol,2011,30(2)∶100-108.
[13]Daisuke Tsuruta,Takashi Hashimoto,Kevin J.Hamilld,Hemidesmosomes and focal contact proteins∶Functions and cross-talk in keratinocytes,bullous diseases and wound healing[J].J Dermatol Sci,2011,62(1)∶1-7.
[14]Iana Turcan,Marcel F.Jonkman.Blisitering Disease∶insight from the hemidesmosome and other components of the dermal-epidermal junction[J].Cell Tissue Res,2015,360(3)∶545-569.
[15]Hirai Y,Takebe K,Takashina M,et al.Epimorphin∶a Mesenchymal Protein Essential for Epithelia Morphogenesis[J].Cell,1992,69(3)∶471-481.
[16]Behrens DT,Villone D,Koch M,et al.The Epidermal Basement Membrane Is a Composite of Separate Laminin- or CollagenⅣ-containing Networks Connected by Aggregated Perlecan,but Not by Nidogens[J].J Biol Chem,2012,287(22)∶18700-18709.
[17]Morgan Dos Santos,Anna Michopoulou,Vaerie Andre-Frei.Perlacan Expression Influences the Keratin 15-Positive cell Population Fate in the Epidermis of Aging Skin[J].Aging,2016,8(4)∶751-768.
[18]Takada K,Amano S,Kohno Y.Non-invasive study of gelatinases in sun-exposed and unexposed healthy human skin based on measurements in stratum corneum[J].Arch Dermatol Res,2006,298(5)∶237-242.
[19]Inomata S,Matsunaga Y,Amano S,et al.Possible involvement of gelatinases in basement membrane damage and wrinkle formation in chronically ultraviolet B-exposed hairless mouse[J].J Invest Dermatol,2003,120(1)∶128-134.
[20]Amano S,Ogura Y,Akutsu N,et al.Protective effect of matrix metalloproteinase inhibitors against epidermal basemen membranedamage∶skin equivalents partially mimic photoageing process[J].Br J Dermatol,2005,153(2)∶37-36.
[21]李贤玉,顾明艳,牟田惠子,等.对中国女性皮肤光老化相关酶-明胶酶的表达研究及含有明胶酶抑制剂的化妆品开发[].第十届中国化妆品学术研讨会论文集,2014.
[22]Miralles F.UV irradiation induces the muring urokinase-type plasminogen activator gene via the c-Jun N-terrminal kinase signaling pathway∶requirement of an API enhancer element[J].Mol Cell Biol,1998,18(8)∶4537-4547.
[23]Ogura Y,Matsunaga Y,Nishiyama T,et al.Plasmin induces degradation and dysfunction of laminin 332(laminin 5) and impaired assembly of basement membrane at the dermal-epidermal junction[J].Br J Dermatol,2008,159(1)∶49-60.
[24]Iriyama S,Hiruma T,Tsunenaga M,et al.Influence of heparan sulfate chains in proteoglycan at the dermal-epidermal junction on epidermal homeostasis[J].Exp Dermatol,2001,20(10)∶810-814.
[25]Sarret Y,Woodley DT,Goldberg GS,et al.Constitutive synthesis of a 92-kDA keratinocyte-derived type Ⅳ collagenase is enhanced by type Ⅰ collagen and decreased by type Ⅳ collagen matrices[J].J Invest Dermatol,1992,99(6)∶836-841.
[26]Iriyama S,Matsunaga Y,Takahashi K,et al.Activation of heparanase by ultraviolet B irradiation leads to functional loss of basement membrane at the dermal-epidermal junction in human skin[J].Arch Dermatol Res,2011,303(4)∶253-261.
[27]Ogura Y,Kadoya K,Matsunaga Y,et al.Basement membrane formation is enhanced by type Ⅳ and Ⅶ collagen-increasing factor,beech bud extract and an inhibitor of the urokinase-plasmin system,peppermint extract[J].J Jpn Cosmet Sci Soc,2005,29(1)∶1-8.
[28]Irimaya J,Kaien K,Tsunenaga M,et al.光老化のおける基底膜へパラン硫酸の変化とその役割 (Role of heparan sulfate at epidermal basement membrane in photo-aging )[J].J Soc Cosmet Chem Jpn,2014,48(1)∶35-40.
编辑/张惠娟
The structure and functions of skin basement membrane
ZHANG Li-li,LI Xian-yu,MU Rong,FAN Chun-hui,Masato Kitagaki,Emiko,Takasu [Shiseido (China) Research Center Co.,LTD.,Beijing 100176,China]
Basement membranes (BE) are a kind of extracellular matrices in every tissue of the whole human body and play the most important role as keeping cellular functions and structures. It is the most important that BE in skin functionally connects and separates with epidermal and dermis containing of various proteins,such as type Ⅳ and Ⅶ collagen,laminins,nidogen and perlecan etc.Those interacted with themselves to form networks to support diverse bio functions in BE. And BE connects between epidermis and dermis signaling etc.It will lead the circulation smoothly among epidermis-basement membrane-dermis that healthy BE contributes diverse bio-functions,so that BE could maintain integrity and healthy of skin.Sunlight exposure may predominantly changes BE,such as sunburn, photoaging and photo-related skin cancers. On the other hand, some studies showed that various herbal extracts could protect BE well.It will be a new approach to develop functional cosmetics that how to utilize those active herbal ingredients.
skin;base membranes; collagens; laminins; multi bio-functions; photoaging; functional herbal extracts
R751
A
1008-6455(2016)10-0113-04
2016-06-27
2016-09-25
猜你喜欢
大自然探索(2022年10期)2022-11-27
西部皮革(2022年19期)2022-11-20
西部皮革(2022年21期)2022-11-16
听力学及言语疾病杂志(2022年5期)2022-09-20
中华耳科学杂志(2021年1期)2021-02-01
家庭医药·快乐养生(2019年2期)2019-03-04
汽车与驾驶维修(维修版)(2018年8期)2018-09-21
中华耳科学杂志(2018年1期)2018-04-19
Coco薇(2017年12期)2018-01-03
饮食与健康·下旬刊(2017年3期)2017-03-30
