
水稻生育后期外源赤霉素调控稻草饲用品质的机理研究
2016-12-06 06:55董臣飞顾洪如丁成龙许能祥张文洁
草业学报 2016年11期
董臣飞,顾洪如,丁成龙,许能祥,张文洁
(江苏省农业科学院畜牧研究所,江苏 南京 210014)
水稻生育后期外源赤霉素调控稻草饲用品质的机理研究
董臣飞,顾洪如,丁成龙,许能祥,张文洁
(江苏省农业科学院畜牧研究所,江苏 南京 210014)
本研究利用两优培九(迟熟中籼)和南粳44(早熟晚粳)两个水稻品种,开花后15 d喷施45 g/hm2赤霉素(GA),在处理后第3和14 天采集水稻倒1叶样品,分析其中内源激素[GA,吲哚乙酸(IAA)和脱落酸(ABA)]的含量及与衰老相关指标丙二醛(MDA)含量、超氧化物歧化酶(SOD)和过氧化物歧化酶(POD)活性,并在收获时采集稻草样品分析饲用品质和青贮品质,分析外源GA调控稻草饲用品质的机理。结果表明外施GA后籼稻品种两优培九和粳稻品种南粳44稻草的饲用品质得到显著改善,衰老进程放缓;但是籼稻和粳稻品种间存在显著差异:两优培九GA处理组稻草的单茎绿叶面积增加幅度极显著高于南粳44(P<0.01),稻草中的非结构性碳水化合物(NSC)和粗蛋白(CP)增加幅度也显著高于南粳44(P<0.05),叶片中的MDA含量显著低于对照(P<0.05),SOD和POD活性显著高于对照(P<0.05),且增加幅度高于南粳44,叶片中的GA含量和GA/ABA均显著高于对照(P<0.05),且高于南粳44。这说明水稻生育后期喷施外源GA能显著改善稻草饲用品质,主要通过减少MDA的积累量,提高SOD和POD活性,提高内源GA含量及GA/ABA来实现,但对衰老进程较快的籼稻品种两优培九的效果优于粳稻品种南粳44。
稻草;赤霉素;饲用品质;调控机理
稻草饲料化利用是当前秸秆综合利用研究的热点。前人对稻草饲用的研究多集中在调制方法上[1-2],但效果受到稻草理化性状及机械设备等因素的限制。通过栽培及遗传育种措施改善稻草饲用品质是提高稻草饲用率的简易方法,但目前这方面的研究较少。在改善稻草饲用品质的农艺措施中,施用外源赤霉素(gibberellic acid,GA)是有效方式。经前期研究表明水稻(Oryzasativa)生育后期进行外源GA处理能增加收获时稻草的绿叶面积,提高稻草中的非结构性碳水化合物(nonstructural carbohydrates, NSC)和粗蛋白(crude protein, CP)含量,有效改善稻草饲用品质[3-4],但缺乏深入的机理研究。
激素在植物生长发育和作物产量形成中起着十分重要的调控作用[5-7]。植物激素通过对基因表达的调控影响植物的生长发育过程。GA是调节植物生长发育的五大激素之一,与其他激素相比,具有价格便宜、效果良好、使用方法简单、毒性低等优势,是目前国内外使用最广泛的植物生长调节剂,主要生理功能为:促进植物细胞分裂、茎叶伸长,加速植物生长发育等。GA还能对植物的抗性产生影响。有研究表明外源GA3处理能提高盐胁迫条件下水稻的植株活力[8],外源GA处理后水稻根系[9-10]、叶鞘[11]、液泡壁[12]的相关蛋白表达水平发生了变化。还有研究表明GA对纤维素合成也具有调控作用[13]。但对于GA改善稻草饲用品质的机理尚未见到相关报道。
在前期研究基础上,本研究采用两优培九和南粳44两个不同类型水稻品种,开花后15 d喷施45 g/hm2GA,在处理后不同时间采集水稻叶片样品,分析其中内源激素含量及与衰老相关指标丙二醛(malondialdehyde, MDA)含量、超氧化物歧化酶(superoxide dismutase, SOD)和过氧化物歧化酶(peroxidase, POD)活性,并在收获时采集稻草样品分析饲用品质和青贮品质,分析外源GA调控稻草饲用品质的机理。
1 材料与方法
1.1 试验材料
利用江苏地区生产中推广应用的2个不同类型的优质水稻品种进行试验:两优培九-杂交籼稻,南粳44-早熟晚粳。
1.2 试验设计与田间管理
试验于2015年在江苏省农业科学院水稻试验田进行。5月10日播种,6月13日插秧。随机区组设计,3次重复,每小区10行,每行10株,1穴1株,行株距为30 cm×20 cm。按照常规水稻进行肥水管理。
1.3 测定内容及方法
水稻开花后挑选长势一致的主茎倒1叶挂牌,在水稻开花后第15 天,即9月8日上午喷药,GA喷施浓度为45 g/hm2。喷药后第3 天(9月11日,即开花后第18 天)和第14 天(9月22日,即开花后第29 天)取样。采集主茎倒1叶,用锡箔纸包好放入液氮罐中冷冻保存。分别用于GA,脱落酸(abscisic acid, ABA),吲哚乙酸(3-indole acetic acid, IAA)3种内源激素的测定及丙二醛(MDA)含量,超氧化物歧化酶(SOD)和过氧化物歧化酶(POD)活性的测定。试验所用GA为上海瑞永生物科技有限公司生产的纯度≥90%的赤霉酸(GA3)。
籽粒蜡熟晚期收获。每小区挑选长势一致且没有挂牌的植株10株收割,留茬高度10 cm,挑选其中长势一致的20个分蘖,测定单茎绿叶面积,然后105 ℃杀青15 min,75 ℃烘干至恒重并称重,粉碎过孔径0.38 mm的筛子,将草粉装入密封样品袋备用,测定非结构性碳水化合物 (NSC)、粗蛋白 (CP)、中性洗涤纤维(neutral detergent fiber, NDF)、酸性洗涤纤维(acid detergent fiber, ADF)的含量和干物质体外消化率(invitrodry matter digestibility,IVDMD)。另取300 g左右样品青贮,3次重复,装入40 cm×30 cm的聚乙烯袋内,真空封口,室温贮藏60 d,开袋检测,利用青贮浸提液测定pH、氨态氮(NH3-N)和乳酸(lactic acid, LA)的含量。
NSC的测定参考Yoshida[14]的方法。CP用丹麦产的蛋白分析仪测定(KJELTEC2300, Foss, Denmark)。NDF、ADF用范氏法测定[15]。IVDMD的测定参考胃蛋白酶-纤维素酶两步法[16]。青贮浸提液的pH用Micro-Bench型pH计直接对青贮浸提液进行测定,LA含量采用对羟基联苯法测定[17]。NH3-N含量采用苯酚-次氯酸钠比色法测定[18]。
内源激素测定方法。激素提取:准确称量约 0.5 g 新鲜样品,于液氮中研磨粉碎;向粉末中加入 5 mL 异丙醇/盐酸提取缓冲液,4 ℃振荡 30 min;加入 10 mL 二氯甲烷,4 ℃振荡 30 min;4 ℃,13000 r/min 离心 5 min,取下层有机相;避光,以氮气吹干有机相,以300 μL甲醇(0.1%甲酸)溶解;过 0.45 μm 滤膜,进 HPLC-MS/MS 检测。
液质检测:标准溶液配制,以甲醇(0.1%甲酸)为溶剂配制梯度为 1,5,10,20,50,200 ng/mL 的 IAA、GA3、ABA 标准溶液;液相条件,色谱柱:安捷伦 ZORBAX 300SB-C18 反相色谱柱(4.6×150,3 μm);柱温:30 ℃;流动相:A∶B=(甲醇/0.1%甲酸)∶(水/0.1%甲酸);洗脱梯度:0~2 min,A=20%;2~14 min,A递增至80%;14~15 min,A=80%;15.1 min,A递减至 20%;15.1~20.0 min,A=20%,进样体积:2 μL。
质谱条件:气帘气:103425 Pa;喷雾电压:4500 V;雾化气压力:448175 Pa;辅助气压力:482650 Pa;雾化温度:400 ℃。
MDA及SOD、POD测定方法。MDA含量测定:准确称量约 0.5 g 新鲜植物样品,于预冷研钵中加 5 mL三氯乙酸(trichloroacetic acid, TCA)提取液研磨至匀浆;所得匀浆在3000 r/min下离心10 min;取离心所得上清液 2 mL,加硫代巴比妥酸(thiobarbituric acid, TBA)2 mL,混合后于100 ℃水浴30 min,冷却后离心;以 TCA 取代提取液做对照,分别在450,532和600 nm处测定吸光值。
POD测定:准确称量约 0.5 g 新鲜植物样品,于预冷研钵中加适量磷酸缓冲液研磨至匀浆;所得匀浆在 3000 r/min下离心10 min,上清液转入25 mL 容量瓶中,沉淀用5 mL缓冲液再提取两次,合并提取液,定容至刻度,低温下保存备用;取酶液 0.1 mL,加入到含有 2.9 mL 缓冲液、1 mL 2%过氧化氢、1 mL 0.05 mol/L的愈创木酚的试管中,立即摇匀,并倒入比色皿中;以缓冲液代替酶液做对照,于470 nm 波长下测定吸光度,以后每隔30 s测定一次,共测定 3~4 min。
SOD测定:准确称量约0.5 g 新鲜植物样品,于预冷研钵中加5 mL磷酸缓冲液研磨至匀浆;所得匀浆在 4000 r/min下离心10 min,上清液即为SOD 提取液,低温下保存备用,取酶液 0.1 mL,加入到含有1.5 mL 缓冲液、0.3 mL 130 mmol/L的甲硫氨酸(methionine,Met)溶液、0.3 mL 750 μmol/L的氮蓝四唑(nitro-blue tetrazolium,NBT)溶液、0.3 mL 100 μmol/L的乙二胺四乙酸二钠(edetate disodium,EDTA-Na2)和20 μmol/L 核黄素的试管中,摇匀后于4000 lx日光下反应20 min;以缓冲液代替酶液做2支对照,1支同样品一起光照处理,一支避光处理,反应结束后,以避光对照管做空白,于560 nm 波长下测定吸光度。
1.4 数据分析
用SPSS 11.5软件进行方差分析,用Excel软件进行作图。
2 结果与分析
2.1 内源激素含量测定结果
GA处理后第3 天取样内源激素含量测定结果见表1。两优培九GA处理组的IAA含量为1.90 ng/g,显著高于对照组(P<0.05),而南粳44 GA处理组的IAA含量为3.80 ng/g,显著低于对照组的4.36 ng/g(P<0.05),南粳44不同处理组的IAA含量均高于两优培九;两优培九GA处理组的ABA含量为5.40 ng/g,显著低于对照组的6.05 ng/g(P<0.05),而南粳44 GA处理组的ABA含量为4.49 ng/g,显著高于对照组的3.59 ng/g(P<0.05),南粳44不同处理的ABA含量均显著低于两优培九(P<0.05);两个品种GA处理组的GA含量均极显著高于对照(P<0.01),两优培九GA处理组的GA含量为242.89 ng/g,对照组仅为5.66 ng/g,而南粳44 GA处理组的GA含量为167.62 ng/g,对照组为6.08 ng/g。
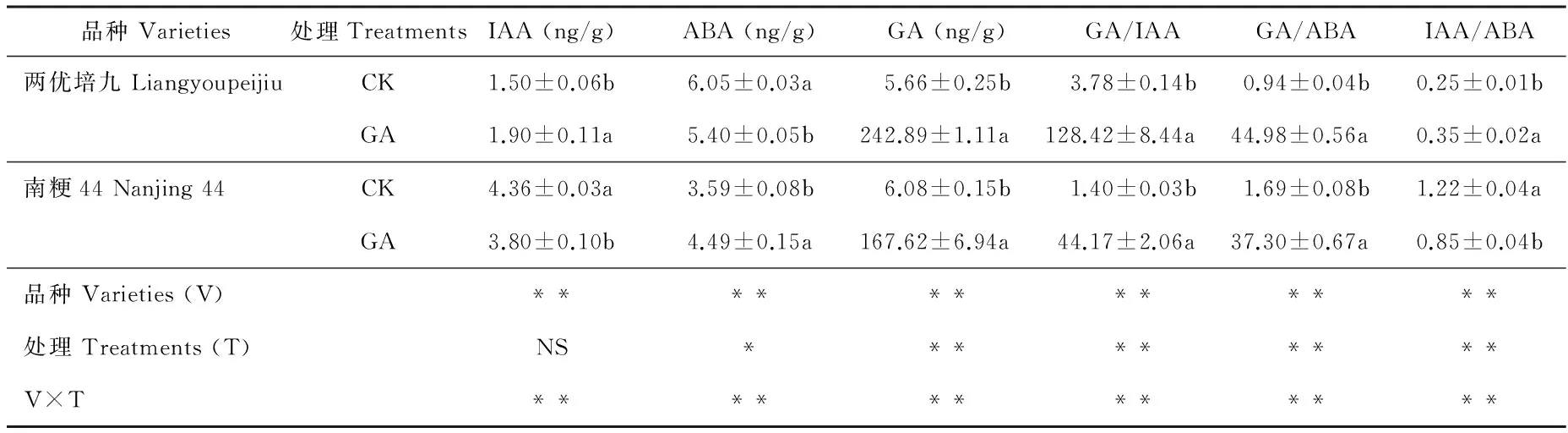
表1 外源GA处理后第3 天水稻叶片内源激素含量
注:* 和**分别代表在P<0.05 和P<0.01水平差异显著。NS表示差异不显著。同一列不同小写字母代表P<0.05水平差异显著。IAA:吲哚乙酸;ABA:脱落酸;GA:赤霉素。下同。
Note: * and ** indicate significant at levelP<0.05 andP<0.01,respectively. NS indicates no significant. Different lowercase letters in the same column mean significantly different atP<0.05 level. IAA: 3-indole acetic acid; ABA: Abscisic acid; GA: Gibberellic acid. The same below.
不同内源激素含量的比值差异显著。两优培九对照组的GA/IAA为3.78,而GA处理组高达128.42,差异极显著(P<0.01),南粳44对照组的GA/IAA为1.40,极显著低于GA组的44.17(P<0.01),两优培九对照组和GA处理组的GA/IAA均显著高于南粳44(P<0.05)。两优培九对照组的GA/ABA为0.94,极显著低于GA处理组(P<0.01),也低于南粳44对照组的1.69,南粳44 GA处理组的GA/ABA为37.30,显著高于其对照组(P<0.05),但低于两优培九GA处理组。两优培九对照组的IAA/ABA为0.25,显著低于GA处理组的0.35(P<0.05),极显著低于南粳44对照组的1.22(P<0.01),南粳44 GA处理组的IAA/ABA为0.85,显著低于其对照组(P<0.05)。
GA处理后第14 天采样内源激素含量测定结果见表2。两优培九GA处理组的IAA含量为2.98 ng/g,显著低于对照组(P<0.05),与第一次取样结果相比IAA含量均显著上升(P<0.05),而南粳44 GA处理组的IAA含量为2.49 ng/g,显著低于对照组的2.89 ng/g(P<0.05),与第一次取样相比IAA含量均有所下降;两优培九GA处理组的ABA含量为8.66 ng/g,显著高于对照组的6.63 ng/g(P<0.05),且较第一次取样结果显著上升(P<0.05),而南粳44 GA处理组的ABA含量为3.80 ng/g,显著高于对照组的3.34 ng/g(P<0.05),且较第一次取样结果显著下降(P<0.05);两个品种GA处理组的GA含量均极显著高于对照(P<0.01),但较第一次取样结果均大幅下降,两优培九GA组为49.33 ng/g,对照组仅为0.98 ng/g,而南粳44 GA处理组的GA含量为42.84 ng/g,对照组为4.62 ng/g。
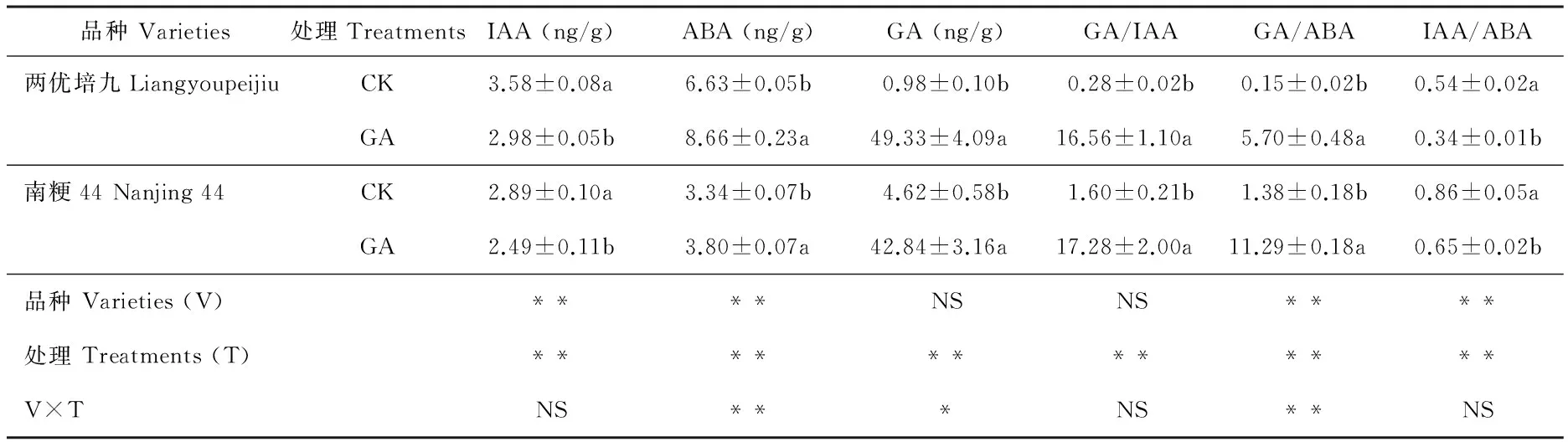
表2 外源GA处理后第14 天水稻叶片内源激素含量
不同内源激素含量的比值差异显著。两优培九对照组的GA/IAA为0.28,极显著低于第一次取样的3.78(P<0.01),而GA处理组为16.56,显著高于对照组(P<0.05),但低于第一次取样的128.42,差异极显著(P<0.01),南粳44对照组的GA/IAA为1.60,高于第一次取样的1.40,但极显著低于GA组的17.28(P<0.01),显著低于第一次取样的44.17(P<0.05),两优培九对照组和GA处理组的GA/IAA均低于南粳44。两优培九对照组的GA/ABA为0.15,极显著低于GA处理组的5.70(P<0.01),也低于南粳44对照组的1.38,南粳44 GA处理组的GA/ABA为11.29,显著高于其对照组(P<0.05),也高于两优培九GA处理组。两优培九对照组的IAA/ABA为0.54,显著高于GA处理组的0.34(P<0.05),南粳44 GA处理组的IAA/ABA为0.65,显著低于其对照组(P<0.05)。
2.2 MDA及SOD和POD测定结果
GA处理后第3 天取样的MDA含量、SOD和POD活性测定结果见表3。两优培九GA处理组的MDA含量为14.86 μmol/g,显著低于对照组(P<0.05),而南粳44 GA处理组的MDA含量为33.56 μmol/g,显著低于对照组的38.94 μmol/g(P<0.05),且南粳44不同处理组的MDA含量均显著高于两优培九(P<0.05);两优培九GA处理组的SOD含量为166.64 U/g,显著高于对照组的125.70 U/g(P<0.05),而南粳44 GA处理组的SOD含量为157.97 U/g,显著低于对照组的173.83 U/g(P<0.05);两优培九GA处理组的POD含量为724.23 U/(g·min),极显著高于对照组的393.93 U/(g·min)(P<0.01),而南粳44 GA处理组的POD含量为776.66 U/(g·min),极显著低于对照组的820.24 U/(g·min)(P<0.01)。南粳44不同处理组的POD含量均显著高于两优培九(P<0.05)。
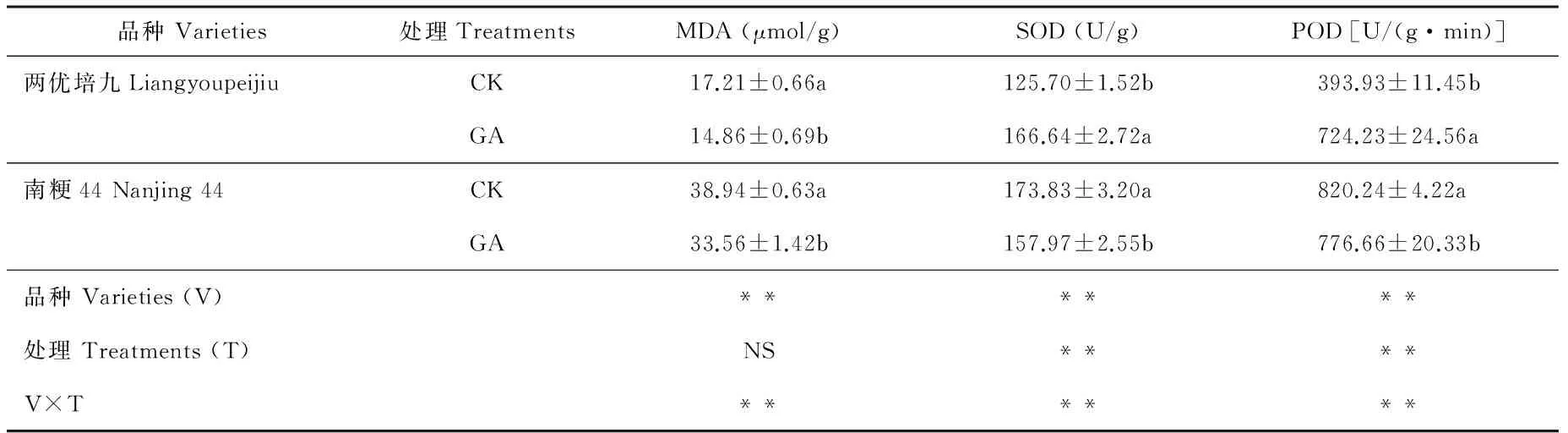
表3 外源GA处理后第3 天水稻叶片MDA含量及SOD和POD活性
注:MDA:丙二醛;SOD:超氧化物歧化酶;POD:过氧化物歧化酶。下同。
Note: MDA: Malondialdehyde; SOD: Superoxide dismutase; POD: Peroxidase. The same below.
GA处理后第14 天取样的MDA含量、SOD和POD活性结果见表4。两优培九GA处理组的MDA含量为2.21 μmol/g,显著低于对照组的3.46 μmol/g(P<0.05),南粳44 GA处理组的MDA含量为2.70 μmol/g,显著低于对照组的4.24 μmol/g(P<0.05),两优培九和南粳44的MDA含量均显著低于第一次取样结果(P<0.05);两优培九GA处理组的SOD含量为180.32 U/g,显著高于对照组的173.19 U/g(P<0.05),而南粳44 GA处理组的SOD含量为170.20 U/g,显著低于对照组的197.43 U/g(P<0.05),南粳44和两优培九不同处理的SOD含量均显著高于第一次取样结果(P<0.05);两优培九GA处理组的POD含量为2845.66 U/(g·min),极显著高于对照组的1853.62 U/(g·min)(P<0.01),而南粳44 GA处理组的POD含量为2261.25 U/(g·min),极显著高于对照组的2144.47 U/(g·min)(P<0.01)。两优培九和南粳44不同处理的POD含量均极显著高于第一次取样结果(P<0.01)。
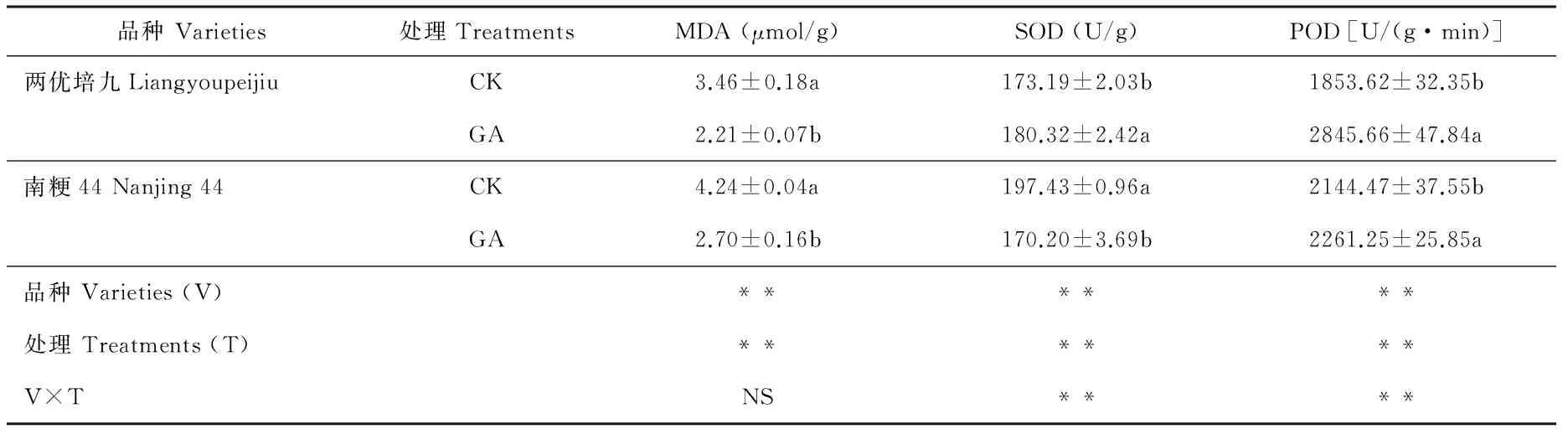
表4 外源GA处理后第14 天水稻叶片MDA含量及SOD和POD活性
2.3 不同处理的稻草饲用品质
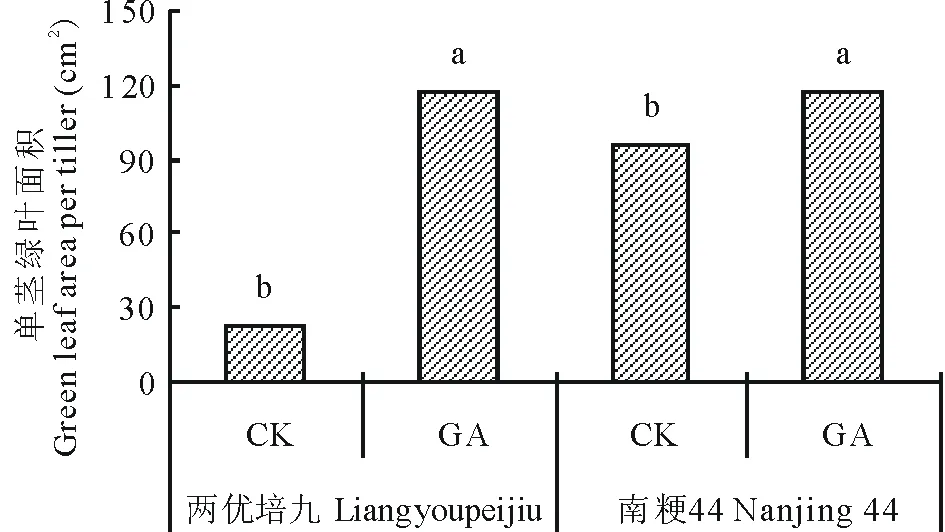
图1 GA处理后两优培九和南粳44的稻草绿叶面积Fig.1 The green leaf area of Liangyoupeijiu and Nanjing 44 after GA application不同小写字母代表P<0.05水平差异显著。Different lowercase letters mean significantly different at P<0.05 level.
GA处理后两优培九和南粳44收获时的单茎绿叶面积见图1。两优培九GA处理组的绿叶面积为117.01 cm2,对照组为22.89 cm2,增幅为411.18%,而南粳44单茎绿叶面积GA处理组为117.06 cm2,对照组为95.41 cm2,增幅为22.69%,极显著低于两优培九(P<0.01)。
外源GA处理后两优培九和南粳44稻草饲用品质性状见表5。两优培九GA处理组稻草的NSC含量极显著高于对照(P<0.01),CP含量高于对照,NDF和ADF差异不显著(P>0.05),IVDMD显著高于对照(P<0.05);而南粳44虽然对照组和GA处理组的稻草饲用品质各性状均优于两优培九,如NSC含量、CP含量均高于两优培九,而NDF和ADF含量则低于两优培九,但GA处理后的效果则与两优培九不同:南粳44 GA组的NSC含量较对照组上升了13.56%,CP含量上升了18.27%,而两优培九则分别上升了91.04%和24.65%。另外,两优培九GA处理组的NDF和ADF含量差异不显著(P>0.05),而南粳44 GA处理组的NDF和ADF含量显著高于对照组(P<0.05)。
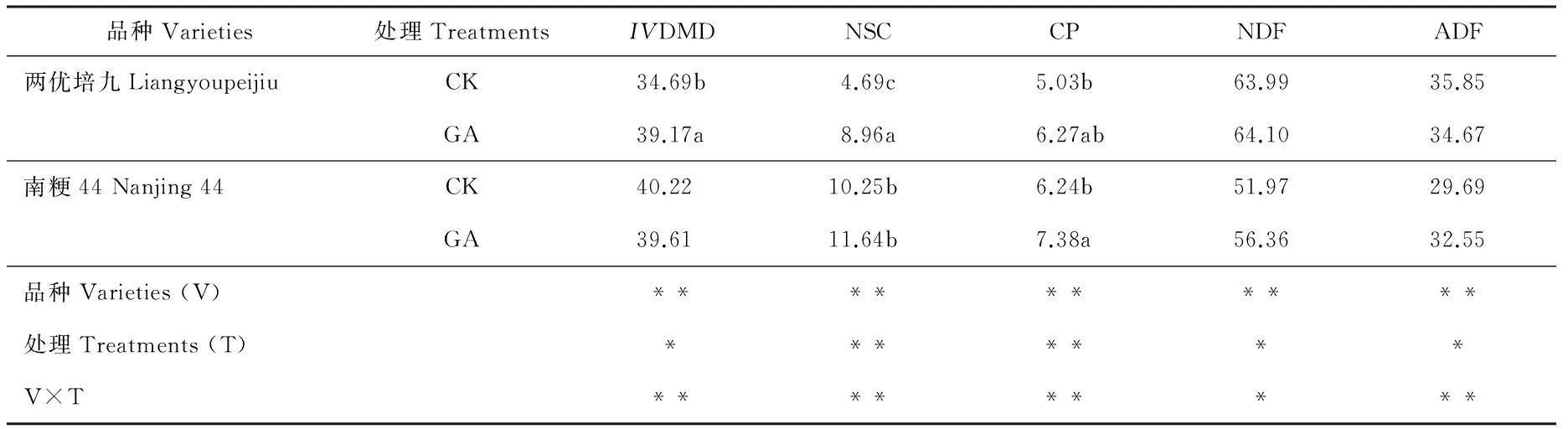
表5 GA处理后的稻草饲用品质性状
IVDMD:干物质体外消化率Invitrodry matter digestibility;NSC:非结构性碳水化合物Nonstructural carbohydrates;CP:粗蛋白 Crude protein;NDF:中性洗涤纤维Neutral detergent fiber;ADF:酸性洗涤纤维Acid detergent fiber。

表6 GA处理后的稻草青贮品质性状Table6 ThefermentationqualityofricestrawafterGAapplication品种Varieties处理TreatmentspHLA(mg/g)NH3-N(mg/g)两优培九LiangyoupeijiuCK4.745.34b9.64aGA4.655.97a8.51b南粳44Nanjing44CK4.3611.864.32GA4.2011.874.29品种Varieties(V)*****处理Treatments(T)NS****V×T***** LA:乳酸Lacticacid.
不同GA 处理后稻草青贮品质见表6。两优培九对照组与GA 处理组的LA 和NH3-N 含量差异显著(P<0.05),GA处理组的LA含量显著高于对照组(P<0.05),NH3-N含量低于对照组,pH差异不显著(P>0.05)。南粳44GA处理组与对照组的青贮品质性状差异均不显著(P>0.05),但所有指标显著优于两优培九(P<0.05),LA含量显著高于两优培九(P<0.05),而NH3-N 含量显著低于两优培九(P<0.05),pH显著低于两优培九(P<0.05)。
3 讨论
3.1 外源GA处理对延缓植株衰老和改善稻草饲用品质的效果
经前期研究表明水稻生育后期稻草中的NSC含量呈现先下降后回升的趋势,CP含量则持续下降[19],若能有效延缓叶片衰老,则有利于促进NSC在稻草中的积累,同时因减缓叶片衰老及叶绿素的分解,可提高稻草CP含量。本研究结果表明外源GA处理后籼稻品种两优培九和粳稻品种南粳44的衰老进程相关指标及稻草饲用品质的变化存在差异:南粳44 GA组稻草中的NSC含量较对照组上升了13.56%,CP含量上升了18.27%,而两优培九则分别上升了91.04%和24.65%(表5)。两优培九GA处理组的绿叶面积增幅为411.18%,而南粳44单茎绿叶面积增幅为22.69%,极显著低于两优培九(P<0.01)(图1)。另外,本研究结果还表明外源GA处理后能有效减少叶片中MDA的积累,提高水稻叶片的SOD和POD活性(表3~4)。郭郁频等[20]研究表明经适宜浓度的GA3处理后,可缓解苜蓿(Medicagosativa)幼苗叶绿素的降解,可降低相对电导率,减少MDA的积累,保持较高的SOD和POD活性,本研究与此结果一致。本研究中两优培九GA处理组的SOD和POD增加幅度均高于南粳44(表3~4),而南粳44对照组在开花后第18天和第29天的SOD和POD活性均高于两优培九对照组(表3~4),这说明南粳44的衰老进程比两优培九缓慢。综合上述结果,说明GA处理对衰老进程慢的粳稻品种南粳44延缓衰老、改善稻草饲用品质的效果不如对衰老进程较快的籼稻品种两优培九显著。
脂质过氧化产物MDA含量是脂质过氧化程度的重要指标之一。它可引起生物大分子的变性或降解及生物合成的降低,破坏细胞结构[21]。刘凯[22]研究表明GA能有效抑制水稻旗叶MDA含量的上升,能有效降低旗叶脂质过氧化的速度,减少MDA对细胞结构的破坏,对延缓细胞的衰亡有一定作用,本研究与此结果一致。SOD、POD是清除细胞活性氧等生物自由基的主要保护酶。SOD是一种诱导酶,能使氧自由基歧化为氧分子和过氧化氢;POD不但能清除过氧化氢,还能使脂质的过氧化物转变为正常的脂肪酸,从而阻止脂质过氧化物的积累引起的细胞中毒[23]。POD是植物膜脂过氧化防御体系中最重要的一种保护酶,其活性的大小反映了植物对逆境引起的膜脂过氧化的防御能力[24]。本研究表明GA处理提高了水稻倒1叶的POD活性值,这对水稻植株清除体内膜脂过氧化物、减小膜伤害的能力是有益的。
3.2 外源GA处理对植株内源激素含量的影响
外源GA处理后,水稻叶片中的内源激素含量发生了显著变化,但籼稻和粳稻品种间存在显著差异。GA处理后虽然内源GA含量及GA/IAA和GA/ABA均高于对照组,但外源GA处理后第14 天的值均较GA处理后第3 天的值出现不同程度的下降(表1~2)。两优培九对照组的GA含量和GA/ABA在两次取样时均显著低于南粳44对照组(P<0.05),ABA含量高于南粳44,两优培九GA处理组的GA含量和GA/ABA在两次取样时均极显著高于对照(P<0.01),改变幅度高于南粳44(表1~2)。
植物激素间可以起相互协同作用或起相互拮抗作用,正是促进和抑制物质之间的相互平衡最终决定植物的生长发育[7,25]。外源GA处理后,水稻内源GA、ABA的含量发生了变化,GA/ABA也随之改变,从而对水稻的生理进程产生影响。这说明延缓植株衰老从而改善稻草饲用品质的机理是因为喷施GA后改变了水稻叶片内源激素的含量及GA/ABA,从而对SOD和POD活性产生调控,高的GA/ABA有利于提高植株抗氧化抗衰老相关酶的活性,进而延缓植株衰老,改善稻草饲用品质。
本研究结果还表明南粳44 GA处理组的NDF和ADF含量显著高于对照组(表5)。外施GA后南粳44稻草中的NDF和ADF含量上升,有可能与GA促进纤维素的合成有关。最近Huang等[13]研究表明GA对纤维素合成的调控作用,通过对水稻GA相关突变体的细胞壁成分测定和基因表达分析,发现GA合成缺陷导致茎秆机械组织厚壁细胞的细胞壁变薄、纤维素含量下降;而GA信号转导抑制子突变和外施GA均能上调纤维素合酶基因的表达,并增加纤维素含量,这表明GA可促进纤维素合成。
4 结论
外施GA后籼稻品种两优培九和粳稻品种南粳44稻草的饲用品质得到显著改善,衰老进程放缓;但是籼稻和粳稻品种间存在显著差异:两优培九GA处理组稻草的单茎绿叶面积增加幅度极显著高于粳稻品种南粳44,稻草中的NSC和CP增加幅度也显著高于南粳44,与之相对应的是叶片中的MDA含量显著低于对照组,SOD和POD活性显著高于对照,且增加幅度高于南粳44,叶片中的GA含量和GA/ABA均显著高于对照组,且高于南粳44。这说明水稻生育后期喷施外源GA能显著改善稻草饲用品质,主要通过减少MDA的积累量,提高SOD和POD活性,提高内源GA含量及GA/ABA来实现,但对衰老进程较快的籼稻品种两优培九的效果优于粳稻品种南粳44。
[1] Li J, Shen Y X, Cai Y M. Improvement of fermentation quality of rice straw silage by application of a bacterial inoculant and glucose. Asian-Australian Journal of Animal Science, 2010, 23(7): 901-906.
[2] Ma Y Y, Li Y F, Cheng Y F,etal. Effects of different chemical treatments on fermentation characteristics of rice strawinvitro. Acta Prataculturae Sinica, 2014, 23(3): 350-355.
[3] Dong C F, Gu H R, Ding C L,etal. Effects of gibberellic acid application after anthesis on the feeding value of double-purpose rice (OryzasativaL.) straw at harvest. Field Crops Research, 2012, 131: 75-80.
[4] Dong C F, Gu H R, Xu N X,etal. Effects of gibberellic acid on the nonstructural carbohydrates content in rice (OryzasativaL) straw harvested at different times. Acta Prataculturae Sinica, 2015, 24(8): 53-64.
[5] Kende H, Zeevaart J A D. The five “classical” plant hormones. Plant Cell, 1997, 9: 1197-1210.
[6] Brenner M L, Cheikh N. The Role of Hormones in Photosynthate Partitioning and Seed Filling[M]//Davies P J. Plant Hormones, Physiology, Biochemistry and Molecular Biology. The Netherlands: Kluwer Academic Publishers, 1995: 649-670.
[7] Davies P J. Introduction[M]//Davies P J. Plant Hormones, Biosynthesis, Signal Transduction, Action. The Netherlands: Kluwer Academic Publishers, 2004: 1-35.
[8] Wen F, Zhang Z, Bai T,etal. Proteomics reveals the effects of gibberellic acid (GA3) on salt-stressed rice (OryzasativaL.) shoots. Plant Science, 2010, 178: 170-175.
[9] Konishi H, Maeshima M, Komatsu S. Characterization of vacuolar membrane proteins changed in rice root treated with gibberellin. Journal of Proteome Research, 2005, 4: 1775-1780.
[10] Komatsu S, Konishi H. Proteome analysis of rice root proteins regulated by gibberellin. Genomics Proteomics Bioinformatics, 2005, 3: 132-142.
[11] Shen S, Sharma A, Komatsu S. Characterization of proteins responsive to gibberellin in the leaf-sheath of rice (OryzasativaL.) seedling using proteome analysis. Biological Pharmaceutical Bulletin, 2003, 26: 129-136.
[12] Komatsu S, Zang X, Tanaka N. Comparison of two proteomics techniques used to identify proteins regulated by gibberellin in rice. Journal of Proteome Research, 2006, 5: 270-276.
[13] Huang D, Wang S, Zhang B,etal. A gibberellin-mediated DELLA-NAC signaling cascade regulates cellulose synthesis in rice. The Plant Cell, 2015, 27: 1681-1696.
[14] Yoshida S. Laboratory Manual for Physiological Studies of Rice[M]. Philippines: Los Baios, IRRI, 1976: 43.
[15] Van Soest P J, Robertson J B, Lewis B A. Methods for dietary fiber, neutral detergent fiber and nonstarch polysaccharides in relation to animal nutrition. Journal of Dairy Science, 1991, 74: 3583-3597.
[16] Goto I, Minson D J. Prediction of the dry matter digestibility of tropical grasses using a pepsin-cellulase essay. Animal Feed Science and Technology, 1977, 2(3): 247-253.
[17] Barker S B, Summerson W H. The colorimetric determination of lactic acid in biological material. Journal of Biology Chemistry, 1941, 138: 535-554.
[18] Weather M W. Phenol-hypochlorite reaction for determinations of ammonia. Annual of Chemistry, 1967, 39: 971-974.
[19] Dong C F, Liu X B, Qu H,etal. Dynamical partition of photosynthates in tillers of rice (OryzasativaL.) during late growth period and its correlation with feeding value of rice straw at harvest. Field Crops Research, 2011, 123(3): 273-280.
[20] Guo Y P, Ren Y X, Liu G H,etal. Effects of calcium (CaCl2), GA3and complex liquid on the physiological characteristics of alfalfa seedlings under drought stress. Acta Prataculturae Sinica, 2015, 24(7): 89-96.
[21] Chen X B, Liao A J, Luo Z M. Physiological properties of the roots and leaves of a high yield rice line at the late growth stage. Life Science Research, 1999, 3(3): 250-255.
[22] Liu K. Regulation of Abscisic Acid and Ethylene to Grain Filling in Rice and Wheat and Its Physiological Mechanism[D]. Yangzhou: Yangzhou University, 2008.
[23] Wang A G, Luo G H, Shao C B. Plant oxygen metabolism and cell damage induced by reactive oxygen species. Collected Papers of China Academy of Sciences, Southern China Institute of Botany, 1989, 5: 11-23.
[24] Chen S Y. Injury of membrane lipid peroxidation to plant cell. Plant Physiology Communications, 1991, 27(2): 84-90.
[25] Davies P J. Introduction[M]//Davies P J. Plant Hormones, Physiology, Biochemistry and Molecular Biology. The Netherlands: Kluwer Academic Publishers, 1995: 1-12.
[2] 马艳艳, 李袁飞, 成艳芬, 等. 不同化学处理对稻草体外发酵动态变化的影响. 草业学报, 2014, 23(3): 350-355.
[4] 董臣飞, 顾洪如, 许能祥, 等. 赤霉素对不同收获时间的稻草中非结构性碳水化合物含量的影响. 草业学报, 2015, 24(8): 53-64.
[20] 郭郁频, 任永霞, 刘贵河, 等. 外源钙和赤霉素对干旱胁迫下苜蓿幼苗生理特性的影响. 草业学报, 2015, 24(7): 89-96.
[21] 陈信波, 廖爱君, 罗泽民. 大穗型水稻生育后期叶片和根系生理的特性. 生命科学研究, 1999, 3(3): 250-255.
[22] 刘凯. 脱落酸和乙烯对水稻与小麦籽粒灌浆的调控作用及其机理[D]. 扬州: 扬州大学, 2008.
[23] 王爱国, 罗广华, 邵从本. 活性氧引起的植物氧代谢和细胞伤害. 中国科学院华南植物研究所集刊, 1989, 5: 11-23.
[24] 陈少峪. 膜脂过氧化对植物细胞的伤害. 植物生理学通讯, 1991, 27(2): 84-90.
Effects of gibberellic acid on forage quality of rice (Oryzasativa) straw
DONG Chen-Fei, GU Hong-Ru, DING Cheng-Long, XU Neng-Xiang, ZHANG Wen-Jie
InstituteofAnimalScience,JiangsuAcademyofAgriculturalSciences,Nanjing210014,China
In this study, two different rice (Oryzasativa) varieties, Liangyoupeijiu (late maturing indica) and Nanjing 44 (early maturing japonica), were used to investigate the mechanisms of gibberellic acid (GA) regulation of the forage value of rice straw during late growth. GA was applied 15 days after anthesis at a rate of 45 g/ha. The endogenous plant hormones [such as GA, 3-indole acetic acid (IAA), and abscisic acid (ABA)], superoxide dismutase (SOD) and peroxidase (POD) activities, and malondialdehyde (MDA) content in the first top leaf were assessed on the 3rd d and 14th d after GA application. Forage quality and fermentation quality of rice straw were also assessed after rice harvest. The results indicated that the forage value Liangyoupeijiu and Nanjing 44 were significantly improved by GA application as a result of reduced senescence. However, there were significant cultivar differences; the green leaf area per tiller of Liangyoupeijiu was significantly higher than that of Nanjing 44 (P<0.01); nonstructural carbohydrates (NSC) and crude protein (CP) content in the straw of Liangyoupeijiu were significantly higher than those of Nanjing 44 (P<0.05). The MDA content of GA treated Liangyoupeijiu was significantly lower than that of the control (P<0.05), while the SOD and POD activities of GA treated plants were significantly higher than those of the control (P<0.05). The GA content and the GA∶ABA ratio of GA treated plants of Liangyoupeiju were significantly higher than those of the control (P<0.05) and Nanjing 44. It was concluded that application of GA during late growth could significantly improve the forage quality of rice straw. The effects of GA were greater in Liangyoupeijiu than in Nanjing 44.
rice straw; gibberellic acid; feeding quality; regulation mechanism
10.11686/cyxb2016014
http://cyxb.lzu.edu.cn
2016-01-11;改回日期:2016-03-25
国家自然科学基金(31302030),江苏省农业科技自主创新资金项目[CX(14)5036]和国家牧草产业技术体系项目(CARS-35-31)资助。
董臣飞(1981-),女,山东青岛人,副研究员,博士。E-mail: cfdong1981@126.com
董臣飞,顾洪如,丁成龙,许能祥,张文洁. 水稻生育后期外源赤霉素调控稻草饲用品质的机理研究. 草业学报, 2016, 25(11): 94-102.
DONG Chen-Fei, GU Hong-Ru, DING Cheng-Long, XU Neng-Xiang, ZHANG Wen-Jie. Effects of gibberellic acid on forage quality of rice (Oryzasativa) straw. Acta Prataculturae Sinica, 2016, 25(11): 94-102.
猜你喜欢
环球时报(2023-01-16)2023-01-16
舰船科学技术(2022年11期)2022-07-15
西藏农业科技(2019年3期)2019-11-04
现代园艺(2018年3期)2018-02-10
上海农业学报(2017年3期)2017-04-10
动物营养学报(2016年7期)2016-08-03
华东理工大学学报(自然科学版)(2015年4期)2015-12-01
动物营养学报(2015年10期)2015-12-01
小说月刊(2014年10期)2014-11-18
云南中医学院学报(2014年3期)2014-07-31
