
北京低山区油松林和元宝枫林凋落物酶活性研究
2016-12-19 21:43赵恒毅耿玉清周红娟张海兰赵广亮
中南林业科技大学学报 2016年6期
赵恒毅 ,耿玉清 ,杨 英 ,周红娟 ,张海兰 ,王 玲 ,赵广亮
(1.北京林业大学 林学院,北京 100083;2.北京市八达岭林场,北京102112)
北京低山区油松林和元宝枫林凋落物酶活性研究
赵恒毅1,耿玉清1,杨 英1,周红娟1,张海兰1,王 玲2,赵广亮2
(1.北京林业大学 林学院,北京 100083;2.北京市八达岭林场,北京102112)
森林凋落物中的酶直接催化凋落物中有机化合物的分解。为探讨不同林分凋落物中酶活性的差异,以北京低山区油松林、元宝枫林以及油松元宝枫混交林为对象,比较不同林分未分解层(L层)和半分解层(F层)凋落物的化学性质以及与碳氮转化相关酶活性的差异,并分析了凋落物化学性质对酶活性的影响。结果表明,与油松林相比,元宝枫林凋落物中L层的α-糖苷酶、β-糖苷酶、几丁质酶、酸性磷酸酶和多酚氧化酶分别高103.66%、158.17%、222.70%、32.18%、242.14%,F层的几丁质酶、亮氨酸氨基肽酶和多酚氧化酶分别高86.10%、63.10%、845.28%;而混交林凋落物中L层的α-糖苷酶、几丁质酶、酸性磷酸酶和多酚氧化酶比油松林凋落物分别高58.81%、58.28%、89.54%、210.64%,F层的几丁质酶、氨基酸氨基肽酶和多酚氧化酶分别高43.98%、46.43%、280.85%。因此,针叶树和阔叶树混交有利于针叶凋落物酶活性的提高,从而促进凋落物的分解。
油松林;元宝枫林;混交林;凋落物层;酶活性
凋落物是森林生态系统的重要组成部分,是土壤物质转化的基础[1];其通过分解作用将养分归还生态系统[2-3],在平衡和维持森林生态系统功能上起到重要作用[4]。凋落物分解包括一系列化学和生物反应,其中酶几乎参与了分解的整个过程。酶活性的提高能加速破坏凋落物结构组成[5],促进有机物质的分解[6]。由于凋落物质量和分解速率的不同[7],不同林分凋落物的酶活性有显著的差异。Kourtev等[8]研究发现不同物种凋落物的β-糖苷酶、纤维素酶和氨基肽酶活性有显著不同。Fioretto等在研究凋落物分解过程中发现纤维素酶和木聚糖酶的活性与微生物的呼吸作用和凋落物类型有密切关系[9]。Dornbush认为,由于不同林分凋落物中理化性质和生物学性质的差异,所以不同凋落物中β-糖苷酶和酸性磷酸酶活性也会大不相同[10]。邵怡若等研究发现采石场废弃地生态恢复中由于植被凋落物的不同,导致土壤凋落物层的脲酶、磷酸酶和过氧化氢酶有显著差异[11]。还有学者认为凋落物酶在不同林分中具有一定的规律,杉木林凋落物中的淀粉酶、蛋白酶、蔗糖酶和磷酸酶活性均显著低于栎树林,而杉木林中的过氧化氢酶和脲酶显著高于栎树林[12]。另有研究表明,杉木-阔叶树混交林凋落物比杉木凋落物含有更高的微生物量,所以脲酶、蛋白酶、过氧化氢酶和脱氢酶的活性显著高于杉木凋落物[13]。因此,不同林分凋落物的酶活性有很大差异,但不同种类酶活性在不同林分间的规律尚未有一致结论。
油松是北京低山区人工林的主要树种,但大面积油松纯林的连栽,可导致土壤肥力退化,生物多样性降低,生态系统稳定性下降[14-15]。而油松林和元宝枫林的混交,可增加凋落物的生物多样性,提高凋落物的分解速率,加快生态系统的能量转化和物质循环[16]。前人已对油松林、元宝枫林以及混交林的结构与功能[17]、物种多样性[18]和凋落物持水特性[19]等进行了深入研究,但关于凋落物酶活性的研究却鲜有报道。为了解不同林分凋落物酶活性的差异,以北京低山区油松林和元宝枫林以及油松-元宝枫混交林的凋落物为研究对象,对不同凋落物层次中与碳氮相关的5种水解酶(α-糖苷酶、β-糖苷酶、几丁质酶、酸性磷酸酶和亮氨酸氨基肽酶)和1种氧化还原酶(多酚氧化酶)的酶活性进行分析,研究结果可为了解混交林凋落物分解机理提供依据,也可为北京低山区森林经营提供参考。
1 材料与方法
1.1 供试材料
试验地位于北京市八达岭林场,地处北纬40°17′,东经 115°55′,属燕山山脉的一部分,平均海拔780 m。该区属温带半湿润大陆性气候,年均气温10.8℃,年均降水量454 mm。现存植被主要为人工林,主要树种有油松Pinus tabulaeformis、元 宝枫Acer truncatum、刺 槐Robinia pseudoacacia、侧柏Platycladus orientalis和黄栌Cotinus coggygria等。土壤类型主要有褐土和棕壤。
2015年5月,在研究地区海拔500~600 m的地段选取坡度、坡向及郁闭度比较一致的油松林、元宝枫林和混交林3个样地(样地基本情况见表1)。在每个样地内随机设置3个20 cm×20 cm的样方,分别收集未分解层(L层)和半分解层(F层)的凋落物,把收集的凋落物清除杂质然后带回实验室。一部分凋落物风干后用超微细粉碎机粉碎,用于化学性质的测定;另一部分新鲜样品保存在0~4℃下,用于凋落物酶活性的测定。
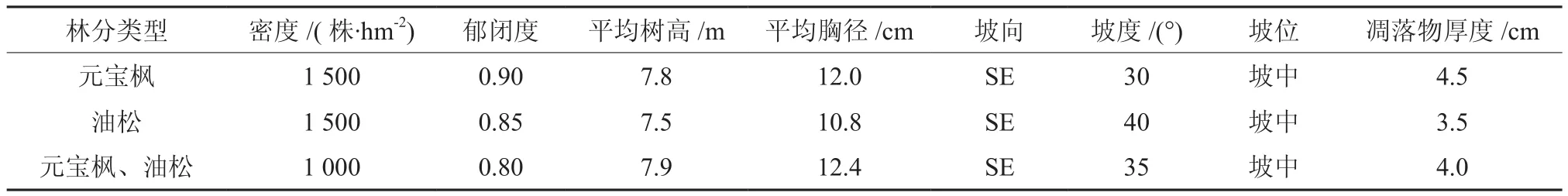
表1 样地基本情况Table 1 The general situation of experimental plots
1.2 分析指标及方法
凋落物pH值采用梅特勒-托利多酸度计(FE20K)测定,浸提液m(叶)∶V(水)比为1∶20;凋落物有机碳采用硫酸重铬酸钾氧化-容量法测得;凋落物全氮测定采用硫酸过氧化氢消煮、碱化后蒸馏定氮的方法;凋落物可溶性有机碳和可溶性全氮的测定:采用0.5 mol/L 纯水浸提(m(叶) ∶m(液) =1∶20),在180次/min的速度下间歇振荡1 h后静置15 min,混合液经过滤后离心15 min,再用0.45 μm滤膜过滤,滤液用德国Multi N/C3100分析仪直接测定[20];水溶性有机氮含量采用水溶性全氮减无机氮含量的方法计算得出[19]。
本研究共涉及6种凋落物酶的分析。水解酶活性测定均采用对硝基苯酚(pNP)[21-22]法,采用pH值5.0的醋酸缓冲液,分别以对硝基苯-α-D-吡喃葡萄糖苷、对硝基苯-β-D-吡喃葡萄糖苷、对硝基苯酚乙酰基氨基葡萄糖苷、对硝基苯磷酸二钠盐为底物测定α-葡萄糖苷酶、β-葡萄糖苷酶、几丁质酶、酸性磷酸酶活性;亮氨酸氨基肽酶活性是采用pH值8.0三羟甲基氨基甲烷缓冲液(Tris),以L-亮氨酸-4-硝基苯胺为底物来测定。氧化还原酶活性采用Sinsabaugh[21]法测定,采用pH5.0的醋酸缓冲液并以左旋多巴(DOPA)为底物测定多酚氧化酶,其中DOPA的吸光系数用酪氨酸酶进行校正[22]。以上6种凋落物酶测定的同时做无土空白试验和无底物对照试验。
2.3 数据处理
实验数据经Excel 2013整理后,用SPSS 22.0进行差异显著性分析和相关性分析。其中,凋落物化学性质和酶活性的差异显著性分析采用Oneway ANOVA法;凋落物化学性质与酶活性的相关性分析采用Pearson法。
3 结果与分析
3.1 不同林分凋落物化学性质的差异
表2数据表明不同林分中不同凋落物层次化学性质有显著差异。在L层中,油松林凋落物的pH值显著高于混交林,但与元宝枫林差异不显著;而油松林凋落物有机碳含量则显著高于混交林,二者又显著高于元宝枫林;对于全氮含量而言,则表现为元宝枫林凋落物显著高于油松林和混交林,但油松林和混交林之间差异不显著;不同林分间凋落物可溶性有机碳和可溶性有机碳差异显著,且均表现为油松林中的含量最高,元宝枫林最低;油松林凋落物中可溶性全氮含量显著高于元宝枫林,但与混交林的差异性未达到显著水平。

表2 不同林分凋落物化学性质的差异†Table 2 The differences of chemical properties in litter of the different forests
在F层中,凋落物pH值在元宝枫林中最高,其显著高于混交林,且混交林显著高于油松林;不同林分间凋落物有机碳含量达到显著水平,且油松林最高;凋落物全氮含量在元宝枫林中最高,其与油松林之间差异显著,但与混交林之间差异不显著;混交林可溶性有机碳含量显著高于元宝枫林,但油松林与前二者差异均不显著;对于可溶性有机碳和可溶性全氮而言,油松林凋落物显著高于元宝枫林和混交林,但后二者差异不显著。
3.2 不同林分凋落物酶活性的差异
表3数据表明不同林分中不同凋落物层次酶活性有显著差异。在L层中,元宝枫林凋落物α-糖苷酶活性显著高于油松林,但混交林与它们的差异均不显著;对于β-糖苷酶活性而言,元宝枫林凋落物最高,显著高于油松林和混交林;三种林分凋落物的几丁质酶和多酚氧化酶活性差异显著,表现为元宝枫林>混交林>油松林;混交林凋落物中酸性磷酸酶活性显著高于元宝枫林,且元宝枫林显著高于油松林;对于亮氨酸氨基肽酶而言,三种林分凋落物之间差异性均达到显著水平,油松林高于元宝枫林,元宝枫林高于混交林。
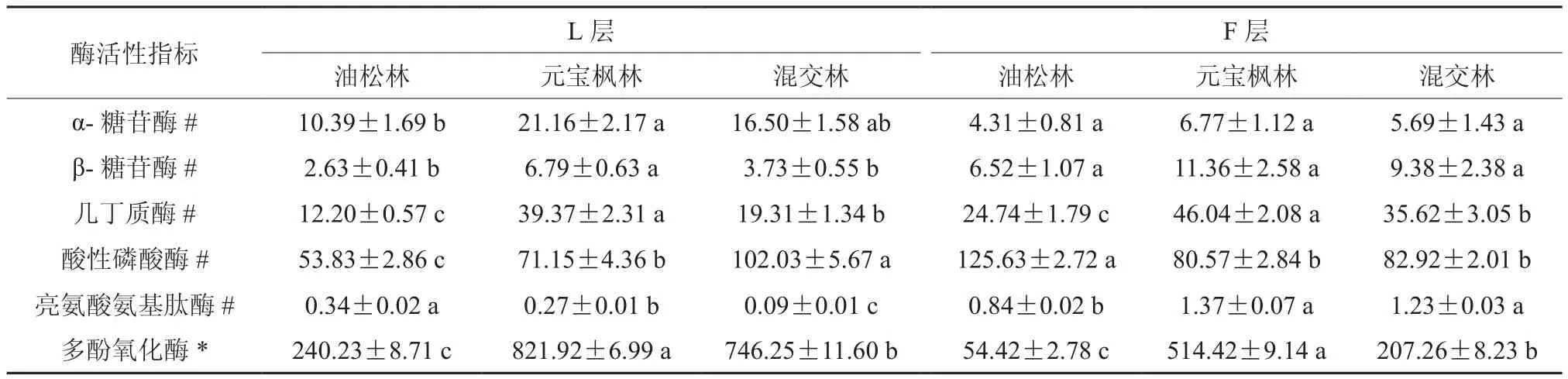
表3 不同林分凋落物酶活性的差异†Table 3 The differences of enzyme activities in the litter of the different forests
在F层中,油松林凋落物中酸性磷酸酶活性最高,显著高于元宝枫林和混交林,但后两者差异不显著;对于亮氨酸氨基肽酶活性而言,元宝枫林和混交林差异不显著,但均显著高于油松林;三种林分凋落物的几丁质酶和多酚氧化酶活性之间的有显著差异,而α-糖苷酶和β-糖苷酶活性的差异不显著。
3.3 凋落物化学性质与酶活性的关系
凋落物的化学性质可显著影响凋落物分解过程中的酶活性。由表4看出,凋落物pH与α-糖苷酶活性呈极显著正相关(P<0.01),但与酸性磷酸酶和亮氨酸氨基肽酶活性呈极显著性负相关,与β-糖苷酶、几丁质酶和多酚氧化酶活性的相关性不显著;凋落物有机碳含量与β-糖苷酶、酸性磷酸酶和亮氨酸氨基肽酶活性呈显著正相关(P<0.05),但与α-糖苷酶和多酚氧化酶活性呈极显著性负相关,与几丁质酶活性无相关性;全氮含量与α-糖苷酶和多酚氧化酶活性呈极显著正相关,与酸性磷酸酶和亮氨酸氨基肽酶活性呈显著负相关,与β-糖苷酶、几丁质酶和酸性磷酸酶活性不相关;可溶性有机碳含量与α-糖苷酶活性呈极显著正相关,与β-糖苷酶、酸性磷酸酶和亮氨酸氨基肽酶活性呈极显著负相关,与几丁质酶活性呈显著负相关,但与多酚氧化酶活性无相关性;可溶性有机碳含量与α-糖苷酶活性呈显著正相关,与β-糖苷酶、酸性磷酸酶和亮氨酸氨基肽酶活性呈极显著负相关,与几丁质酶活性呈显著负相关,但与多酚氧化酶活性相关性不显著;可溶性全氮含量仅与多酚氧化酶活性呈显著负相关,而与其他的酶活性均无相关性。
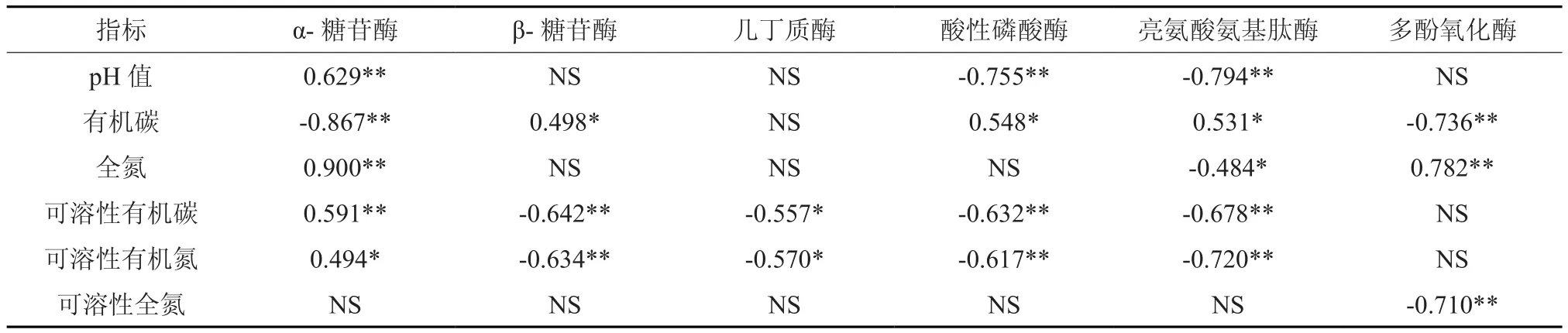
表4 凋落物化学性质与酶活性相关性分析†Table 4 The relationship between chemical properties and enzyme activities in litter layer
4 讨 论
4.1 不同林分对凋落物酶活性的影响
不同林分凋落物由于组成成分不同,所以酶活性有很大差异[24-25]。阔叶凋落物中的C/N较低,且含有大量的糖、氨基酸和脂肪酸等易分解的化合物,有利于微生物的活动;但针叶凋落物由于其含水量少且C/N高等因素,不利于微生物的生长[26],而且有较高含量的单宁,其与蛋白质能络合成丹宁-蛋白质复合物,导致酶活性降低,甚至完全失活[27]。从理论上推断,阔叶凋落物酶活性高于针叶凋落物。Waring在研究不同林分凋落物分解与酶活性的关系时发现,β-糖苷酶和氨基肽酶的活性在阔叶林中显著高于针叶林[5]。还有一些研究也指出,β-糖苷酶和几丁质酶活性的最高值都在常绿阔叶林,且显著高于针叶纯林[26,28]。但有学者却认为部分酶在不同林分中的活性并没有阔叶林高于针叶林的规律,比如多酚氧化酶和酸性磷酸酶[12]。本研究发现不同酶种类在不同林分类型间的差异有别,α-糖苷酶、β-糖苷酶、几丁质酶和多酚氧化酶在元宝枫林凋落物中的活性高于油松林,这与前人研究基本一致[26,28];但元宝枫L层酸性磷酸酶和F层的亮氨酸氨基肽酶活性均高于油松林,这与Waring[5]和戴凌[29]的一些研究结果相似,可能是由于油松林凋落物中的pH和微生物营养源等条件比元宝枫林更适合分泌酸性磷酸酶和亮氨酸氨基肽酶的微生物生长[30]。综合看来,元宝枫林凋落物的酶活性较油松林高。
针阔叶混交后,阔叶树凋落物能提供较高的N素,降低了C/N,从而有利于微生物的活动。所以,混交林凋落物酶活性可能会高于针叶纯林[5]。有学者认为混交林凋落物中的糖苷酶和多酚氧化酶活性要显著高于针叶林[28]。但也有研究发现在凋落物分解中期时,槲栎-马尾松混交林凋落物的酸性磷酸酶酶活性显著低于槲栎和马尾松纯林[30]。所以,混交林凋落物酶的活性是否高于针叶林与酶的种类有关。本研究认为,混交林凋落物中的α-糖苷酶、β-糖苷酶、几丁质酶和多酚氧化酶活性较油松林有所提高,这与Hu[31]和黄宇[13]等人的研究结果相似;但本研究还发现,混交林凋落物中L层的亮氨酸氨基肽酶和F层的酸性磷酸酶活性比油松林低,这可能是混交林凋落物在分解过程中影响了底物的有效性[30],降低了酶活性。总之,凋落物酶活性在不同林分中有明显差异,其差异程度与酶的种类和凋落物层次有密切关系。
4.2 不同凋落物层次对酶活性的影响
凋落物酶在不同的凋落物层次的规律有很大差异[32]。凋落物层次对酶活性的影响,主要是由于凋落物层质量[33]和凋落物分解程度[34]的变化致使微生物生物量发生改变,从而使其产生的酶种类和活性也发生变化。Andersson等[32]认为随着凋落物分解的进行,微生物活性和酶活性有显著变化,但F层中的几丁质酶始终高于L层。有学者在研究不同发育阶段日本落叶松人工林时发现,凋落物层糖苷酶、几丁质酶和酸性磷酸酶活性均表现为F层>L层[35]。还有人在研究凋落物酶活性季节动态时发现阔叶林L层中β-糖苷酶活性高于F层[26]。但也有学者发现了与他人不同的规律,针叶林F层凋落物中的β-糖苷酶活性在3月、5月和7月时显著高于L层[27]。所以,不同凋落物层次中各种酶活性的变化有很大差异。本研究发现,L层的α-糖苷酶和多酚氧化酶显著高于F层,这可能是因为F层中可利用的底物数量比L层少,也可能是在凋落物随着分解的进行产生了难分解物质,影响了产生α-糖苷酶和多酚氧化酶微生物的活性[35]。F层β-糖苷酶、几丁质酶、酸性磷酸酶和亮氨酸氨基肽酶活性均高于L层,这可能是由于L层产生酶的微生物活性较低,而随着L层凋落物不断分解,在转入到F层的过程中,产生该酶的微生物活性不断升高,同时提高了酶活性[36]。所以,不同凋落物层次的凋落物中的酶活性有很大的不同[30]。
5 结 论
凋落物的分解是由多种酶共同作用来完成的,林分种类可直接影响凋落物化学性质以及酶活性,其影响程度与凋落物层次以及酶的种类有关。通过对三种林分不同凋落物层次中酶活性的研究发现:(1)凋落物L层中的α-糖苷酶和多酚氧化酶显著高于F层,而F层中的β-糖苷酶、几丁质酶、酸性磷酸酶和亮氨酸氨基肽酶活性均高于L层;(2)元宝枫林凋落物中的α-糖苷酶、β-糖苷酶、几丁质酶和多酚氧化酶活性均高于油松林,但F层的酸性磷酸酶和L层的亮氨酸氨基肽酶活性却低于油松林;(3)元宝枫和油松混交后,α-糖苷酶、β-糖苷酶、几丁质酶和多酚氧化酶活性较油松林均有显著提高,但酸性磷酸酶和亮氨酸氨基肽酶活性却由于凋落物层次的差异呈现出不同程度的提高或下降。据此可知,针阔林混交可提高凋落物酶的活性,加快凋落物的分解。但由于条件限制,本研究在实验过程中并未考虑相关环境因素如水、热和光照条件以及微生物数量和种类等对凋落物酶活性所引起的差异。在未来研究中需要对引起差异的因素全面考虑,并同步监测凋落物分解过程中酶活性的动态变化,以便更深入地说明凋落物分解过程中酶活性的变化以及酶活性对凋落物分解的作用。
[1] 李 云, 周建斌, 董燕捷, 等. 黄土高原不同植物凋落物的分解特性[J]. 应用生态学报, 2012, 23(12):3309-3316.
[2] 陈晓丽, 王根绪, 杨燕, 等. 山地森林表层土壤酶活性对短期增温及凋落物分解的响应[J]. 生态学报, 2015, 35(21): 7071-7079.
[3] 林 波, 刘 庆, 吴 彦 , 等.森林凋落物研究进展[J]. 生态学杂志, 2004, 23(1):60-64.
[4] Ge X, Zeng L, Xiao W, et al. Effect of litter substrate quality and soil nutrients on forest litter decomposition: A review[J].ActaEcologicaSinica, 2013, 33(2):102-108.
[5] Waring B G. Exploring relationships between enzyme activities and leaf litter decomposition in a wet tropical forest[J]. Soil Biology & Biochemistry, 2013, 64(9):89-95.
[6] 葛晓改, 肖文发, 曾立雄, 等. 三峡库区马尾松林土壤-凋落物层酶活性对凋落物分解的影响[J]. 生态学报, 2014,34(9):2228-2237.
[7] Criquet S, Ferre E, Farnet A M, et al. Annual dynamics of phosphatase activities in an evergreen oak litter: influence of biotic and abiotic factors[J]. Soil Biology & Biochemistry, 2004,36(7):1111–1118.
[8] Kourtev P S, Ehrenfeld J G, Huang W Z. Enzyme activities during litter decomposition of two exotic and two native plant species in hardwood forests of New Jersey[J]. Soil Biology &Biochemistry, 2002, 34(9):1207–1218.
[9] Fioretto A, Papa S, Sorrentino G, et al. Decomposition of Cistusincanus leaf litter in a Mediterranean maquis ecosystem:mass loss, microbial enzyme activities and nutrient changes[J].Soil Biology & Biochemistry, 2001, 33(3):311–321.
[10] Dornbush M E. Grasses, litter, and their interaction affect microbial biomass and soil enzyme activity[J]. Soil Biology &Biochemistry, 2007, 39:2241–2249.
[11] 邵怡若, 许建新, 薛 立,等. 黄石采石场废弃地生态恢复过程中的土壤变化[J]. 中南林业科技大学学报,2014,34(4):82-89.
[12] 肖慈英, 阮宏华, 屠六邦. 宁镇山区不同森林土壤生物学特性的研究[J]. 应用生态学报, 2002, 13(9):1077-1081.
[13] 黄 宇, 汪思龙, 冯宗炜, 等. 不同人工林生态系统林地土壤质量评价[J]. 应用生态学报, 2004, 15(12):2199-2205.
[14] 杨承栋, 孙启武, 焦如珍, 等. 大青山一二代马尾松土壤性质变化与地力衰退关系的研究[J].土壤学报,2003,40(2):267-273.
[15] 唐以杰, 方展强, 何 清, 等. 无瓣海桑与乡土红树植物混交对林地大型底栖动物的影响[J].生态学报, 2015, 35(22):7355-7366.
[16] 杨曾奖, 曾 杰, 徐大平, 等. 森林枯枝落叶分解及其影响因素[J]. 生态环境, 2007, 16(2):649-654.
[17] 鲁绍伟, 刘凤芹, 余新晓, 等. 华北土石山区油松-元宝枫混交林的结构与功能[J]. 东北林业大学学报, 2007, 35(9):20-23.
[18] 邓宏兼, 李卫忠, 曹 铸, 等. 基于不同取样尺度的油松针阔混交林物种多样性[J]. 浙江农林大学学报, 2015, 32(1):67-75.
[19] 樊登星, 余新晓, 岳永杰, 等. 北京西山不同林分枯落物层持水特性研究[J]. 北京林业大学学报, 2008, 30(Z2):177-181.
[20] Chen C R, Xu Z H, Keay P, et al. Total soluble nitrogen in forest soils as determined by persulfate oxidation and by high temperature catalytic oxidation[J]. Soil Research, 2005,43(4):515-523.
[21] Sinsabaugh R L, Klug M J, Collins H P, et al. Characterizing soil microbial communities[C]// Robertson G P, Coleman D C, Bledsoe C S, et al. Standard Soil Methods for Long-term Ecological Research. New York: Oxford University Press, 1999:329-338.
[22] Verchot L V, Borelli T. Application of para-nitrophenol (pNP)enzyme assays in degraded tropical soils[J]. Soil Biology and Biochemistry, 2005, 37(4): 625-633.
[23] Allison S D, Vitousek P M. Extracellular Enzyme Activities and Carbon Chemistry as Drivers of Tropical Plant Litter Decomposition[J]. Biotropica, 2004, 36(3):285-296.
[24] Hättenschwiler S, Tiunov AV, Scheu S. Biodiversity and litter decomposition in terrestrial ecosystems[J]. Annual Reviews,2005, 36(36):191-218.
[25] Tian L, Shi W. Short-term effects of plant litter on the dynamics, amount, and stoichiometry of soil enzyme activity in agroecosystems[J]. European Journal of Soil Biology, 2014, 65:23-29.
[26] 张 鹏, 田兴军, 何兴兵, 等. 亚热带森林凋落物层土壤酶活性的季节动态[J]. 生态环境, 2007, 16(3):1024-1029.
[27] Kraus T E C, Dahlgren R A, Zasoski R J. Tannins in nutrient dynamics of forest ecosystems - a review[J]. Plant & Soil, 2003,256(1):41-66.
[28] 齐泽民, 王开运, 宋光煜, 等. 川西亚高山箭竹群落枯枝落叶层生物化学特性[J]. 生态学报, 2004, 24(6):1230-1236.
[29] 戴 凌, 黄志宏, 文 丽. 长沙市不同森林类型土壤养分含量与土壤酶活性[J]. 中南林业科技大学学报, 2014, 34(6):100-105.
[30] 宋 影, 辜夕容, 严海元, 等. 中亚热带马尾松林凋落物分解过程中的微生物与酶活性动态[J]. 环境科学, 2014, 35(3):1151-1158.
[31] Hu Y L, Wang S L, Zeng D H. Effects of single Chinese fi r and mixed leaf litters on soil chemical, microbial properties and soil enzyme activities[J]. Plant & Soil, 2006, 282:379-386.
[32] Andersson M, AnneliseKjøller, Struwe S. Microbial enzyme activities in leaf litter, humus and mineral soil layers of European forests[J]. Soil Biology & Biochemistry, 2004,36(10):1527-1537.
[33] 罗 佳, 周小玲, 田育新, 等. 武陵山区小流域不同年龄结构杜仲人工林生物量研究[J]. 经济林研究,2015,33(3):99-102.
[34] 汪思龙. 森林残落物生态学[M]. 北京: 科学出版社, 2010.
[35] 牛小云, 孙晓梅, 陈东升, 等. 日本落叶松人工林枯落物土壤酶活性[J]. 林业科学, 2015, 51(4): 16-25.
[36] 张 兵, 苏淑钗, 陈 凤, 等. 菌渣覆盖对榛子园土壤酶活性及理化性质的影响[J]. 经济林研究, 2015, 33(1):33-38.
Enzyme activities in litter of Pinus tabulaeformis and Acer truncatum forests in lower mountain area, Beijing
ZHAO Heng-yi1, GENG Yu-qing1, YANG Ying1, ZHOU Hong-juan1, ZHANG Hai-lan1, WANG Ling2, ZHAO Guang-liang2
(1. College of Forestry, Beijing Forestry University, Beijing 100083, China; 2. Beijing Badaling Forest Farm, Beijing 102112, China)
Enzyme in the forest litter can directly catalyze compounds decomposition in the litter.This objective of this paper was to investigate the variation of enzyme activity in litter of the different forests.Three forest plots included Acer truncatum forest,Pinus tabulaeformis forest and mixed forest were established.The chemical properties and the enzyme activities (α-glycosidase,β-glycosidase, glucosaminidase, acid phosphatase, leucine aminopeptidase and polyphenol oxidase)involved in the transformations of carbon and nitrogen of litter were compared in litter layer and fragmentation layer of the different forests, and the correlation of litter chemical properties and the enzyme activities was analyzed.The results showed that the activities of α-glycosidase, β-glycosidase,glucosaminidase, acid phosphatase and polyphenol oxidase in litter layer of Acer truncatum forest was higher than Pinus tabulaeformis forest by 103.66%, 158.17%, 222.70%, 32.18%, 242.14%, respectively. Activities of glucosaminidase, leucine aminopeptidase and pdyphenol oxidase in fragmentation layer were increased by 86.1%, 63.1%, 845.28%; the activities of α-glycosidase, glucosaminidase,acid phosphatase and polyphenol oxidase in litter layer of mixed forest litter was higher than Pinus tabulaeformis forest by 58.81%,58.28%, 89.54%, 210.64%, the activities of chitinase, leucine aminopeptidase and polyphenol oxidase in fragmentation layer was higher by 43.98%, 46.43%, 280.85%. In sum, coniferous and broadleaf mixed may stimulate the enzyme activity, thus accelerate the litter decomposition.
Pinus tabulaeformis forest; Acer truncatum forest; mixed forest; litter layer; enzyme activity
S791.254
A
1673-923X(2016)06-0023-06
10.14067/j.cnki.1673-923x.2016.06.005
2015-09-21
北京市财政项目(PXM2015-154309-000010-00174287-FCG);;““十十二二五五””国国家家科科技技支支撑撑计计划划项项目目(2012BAC16B03)
赵恒毅,硕士研究生
耿玉清,副教授,硕士研究生导师;E-mail:gengyuqing@bjfu.edu.cn
赵恒毅,耿玉清,杨 英,等. 北京低山区油松林和元宝枫林凋落物酶活性研究[J].中南林业科技大学学报,2016, 36(6):23-28.
[本文编校:吴 彬]
猜你喜欢
国外畜牧学(猪与禽)(2022年3期)2023-01-06
中国林业产业(2021年5期)2021-12-14
昆明医科大学学报(2020年12期)2021-01-26
食品与生物技术学报(2021年4期)2021-01-17
民主(2019年4期)2019-08-15
吉林农业(2019年9期)2019-01-06
天然产物研究与开发(2018年8期)2018-09-10
中国医药生物技术(2017年2期)2017-04-15
农村农业农民·B版(2016年12期)2017-01-11
家庭用药(2016年9期)2016-12-03
