
铬污染对延胡索生长和生理特性的影响
2017-02-05 19:48余顺慧方荣美唐洁谭俊陆亚
江苏农业科学 2016年10期
余顺慧++方荣美++唐洁++谭俊++陆亚萍++罗杨++孙嘉
doi:10.15889/j.issn.1002-1302.2016.10.064
摘要:为了阐明Cr6+污染对延胡索的毒害机理,探索重金属对延胡索生长发育的影响。采用土培试验,研究不同浓度的Cr6+(0、50、75、100、150、200、250、300 mg/kg)对延胡索生长、抗氧化酶活性和Cr6+积累等的影响。结果表明,延胡索在Cr6+污染下生长受抑制,生物量下降;Cr6+污染能明显影响延胡索抗氧化酶活性,即超氧化物歧化酶(SOD)活性随Cr6+浓度的升高而逐渐降低,过氧化物酶(POD)的活性随Cr6+浓度的升高而逐渐增加,过氧化氢酶(CAT)活性均随Cr6+浓度的升高而先增后减;Cr6+污染能明显影响延胡索叶绿素含量、可溶性糖含量、丙二醛(MDA)含量等。叶绿素含量与可溶性糖的含量随着Cr6+浓度的升高而先增后减,而MDA含量逐渐增加。随着Cr6+污染浓度的升高,延胡索对Cr6+的积累在地下块茎逐渐增多,在Cr6+浓度为150 mg/kg时Cr6+含量增加速度最大,生产上应该注意土壤中的Cr6+含量。延胡索栽培中应尽量选择Cr6+污染较少的土地进行。
关键词:铬污染;延胡索;生长;生理特性;栽培模式
中图分类号: S567.21+9.01文献标志码: A文章编号:1002-1302(2016)10-0236-04
收稿日期:2015-08-12
基金项目:重庆市应用开发计划(编号:cstc2014yykfA110024)。
作者简介:余顺慧(1964—),女,重庆万州人,教授,主要从事植物生态学研究。Tel:(023)58105874;E-mail:ysh_dch@163.com。延胡索别称元胡、玄胡索、玄胡,为罂粟科紫堇属一年生草本植物,以其地下块茎入药,为常用中药,具有行气、活血、止痛的作用。铬广泛存在于环境中,在自然状态下,铬离子有Cr+3和Cr6+ 2种形态,其中Cr6+则是一种毒性较大的致畸、致突变离子[1]。Cr6+容易被植物吸收,能在植物体残留对植物产生毒害[2-5]。而农作物受到铬毒害后,不但严重影响农产品的产量和质量,更为严重的是通过食物链影响人畜健康[6]。随着现代工业的发展,铬在化工行业中的广泛应用,导致环境中的铬总量显著增加,致使土壤、水体和生物遭到不同程度的污染。目前对延胡索的研究主要集中在测定生物碱含量方面,但是有关重金属对延胡索的产量和品质的影响鲜有报道。笔者所在实验室用重金属铜、铅等胁迫狗牙根以研究其耐性机理,并取得了一定的成绩[7-10]。本试验以延胡索为研究对象,设置不同浓度Cr6+处理,测定不同处理延胡索生长发育状况及抗氧化酶活性等,为了提高延胡索的产量,优化其质量,提高经济效益,为栽培模式提供实践指导和理论依据,通过对延胡索进行不同铬浓度的处理,探讨重金属对其生长的影响。
1材料与方法
1.1材料
供试材料采于重庆三峡学院百安坝校区实验基地。
1.2方法
1.2.1试验设计本试验在重庆三峡学院百安坝校区实验基地内完成。试验材料为2014年10月由浙江省东阳市农业局提供的延胡索块茎,从中挑选出大小基本一致的地下块茎105块,移栽至深25 cm、内径30 cm的花盆中,每盆装土壤 8 kg,进行常规管理,让其自然生长。
采用7个处理,1个对照,3组平行,每盆装小粉土,按含铬0、50、75、100、150、200、250、300 mg/kg分别加入K2Cr2O7混和均匀,重复3次,种植延胡索5块于每盆中。对照(CK)不含铬,定期检查生长情况。持续处理4个月时间,试验过程中除了重金属浓度不同以外,保持其余环境条件完全一致。
供试土壤肥力基础为:有机质含量89%,全氮含量023%,水解氮含量12.58 mg/kg,速效钾含量50.34 mg/kg,速效磷含量11.34 mg/kg,含铅13.4 mg/kg。
1.2.2测定指标120 d后延胡索出现伤害症状,选取各植株上部第3或4张叶用于测定叶绿素含量、超氧化物歧化酶(SOD)活性、过氧化物酶(POD)活性、过氧化氢(CAT)活性、丙二醛(MDA)活性和可溶性糖含量。
生物量的测定:在延胡索出现伤害症状后,收获,取其地下部分洗净称鲜质量,烘箱中80 ℃烘干至恒质量称干质量。
叶绿素含量采用丙酮-乙醇分光光度法[11]测定,3个平行,计算平均值。
叶绿素的提取:鲜质量叶片剪成细丝状后准确称取0.2 g放入加有25 mL混合提取液(丙酮、乙醇体积比为1 ∶1)的洁净试管中,封口后将试管置于暗处直至叶片颜色完全变白。
使用江苏南京建成生物工程研究所购买的SOD、POD试剂盒测定SOD、POD活性。CAT活性测定采用分光光度法[11];丙二醛(MDA)含量测定采用硫代巴比妥酸法[12];可溶性糖含量测定采用蒽酮法[13];微波消解样品测定其重金属铬含量。
收获后,延胡索地下部分分别用自来水、去离子水冲洗3次,80 ℃烘干,粉碎后,精确称取延胡索各0.200 0 g,加入酸液(浓HNO3、浓H2SO4体积比为4 ∶1),同时做空白组;用MARS240微波消解系统消解以上溶液后,冷却。过滤(滤液在25 mL容量瓶中定容),保存在塑料瓶中,样品中Cr6+含量(mg/kg干质量)用AA-6300原子吸收分光光度计测定。
1.2.3统计分析数据处理和分析采用Sigmapiot 10.0软件;采用单因素方差分析方法(one-way ANOVA)进行平均值间的比较。
2结果与分析
2.1铬污染对延胡索生物量的影响
如表1所示,随着Cr6+处理浓度升高,延胡索块茎的生物量呈现出递减趋势。不同Cr6+处理条件下生物量差异不显著(P>0.05)。这表明延胡索对Cr6+有一定的耐性。研究表明,生物量在Cr6+处理浓度为200 mg/kg时减少幅度突然增加,鲜质量由 150 mg/kg 的28.40%增加到200 mg/kg的54.47%,干质量由150 mg/kg的30.20%增加到200 mg/kg的65.77%(均和对照相比),说明Cr6+污染浓度200 mg/kg对延胡索生长和产量有明显的影响。在生产上要注意土壤中Cr6+含量。
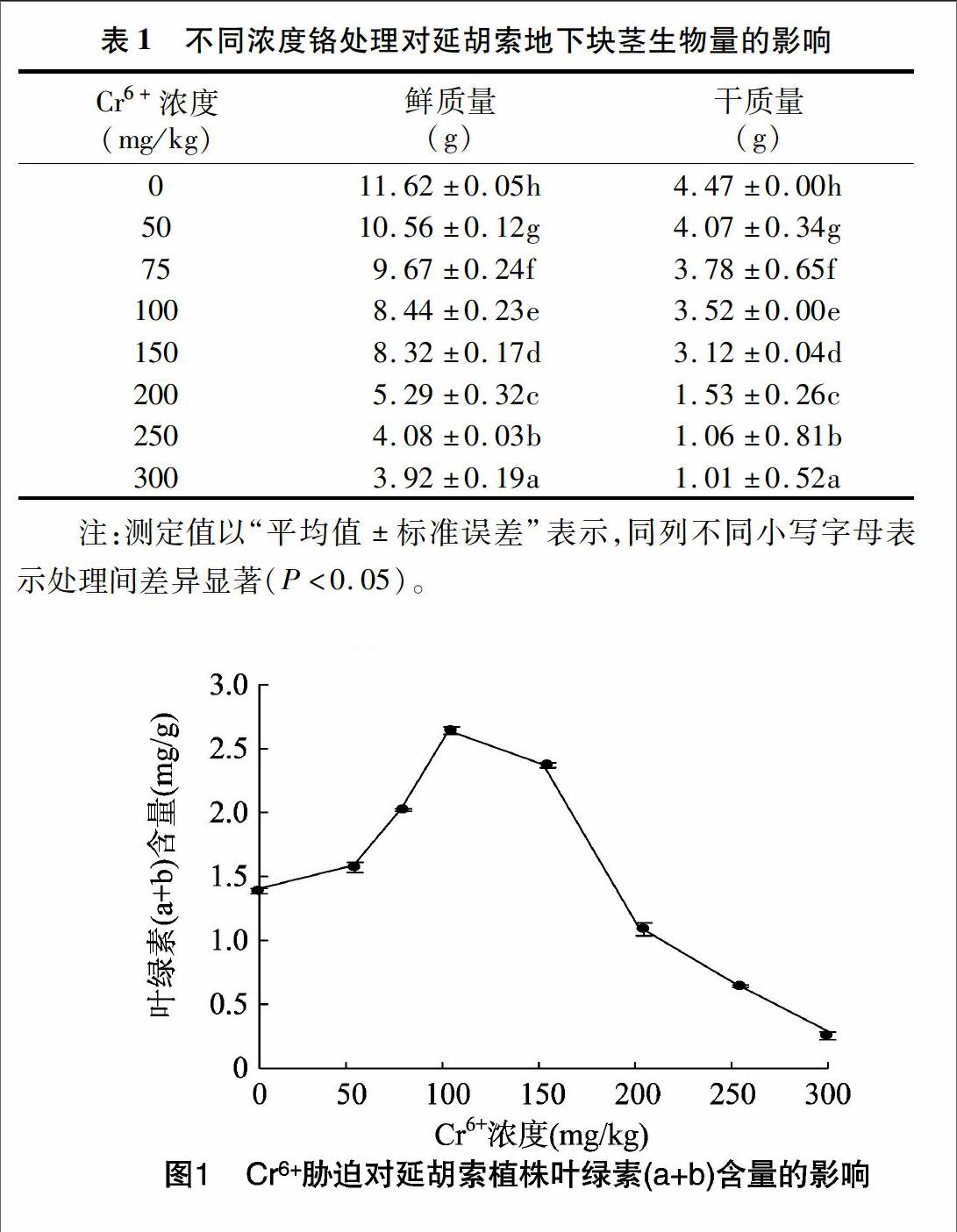
2.2Cr6+污染对延胡索叶绿素含量的影响
植物进行光合作用必需的色素为叶绿素,一定程度上其含量高低反映了光合作用水平,叶绿素含量低,光合作用弱。由图1可见:延胡索经Cr6+处理后,与对照相比,在Cr6+浓度≤100 mg/kg时,叶绿素(a+b)含量随Cr6+浓度的逐渐降低而减少;随着Cr6+处理浓度的升高,各处理之间的差异越明显,叶绿素含量在Cr6+浓度>150mg/kg时明显下降,说明对延胡索叶片的伤害作用随着Cr6+浓度升高而增大。
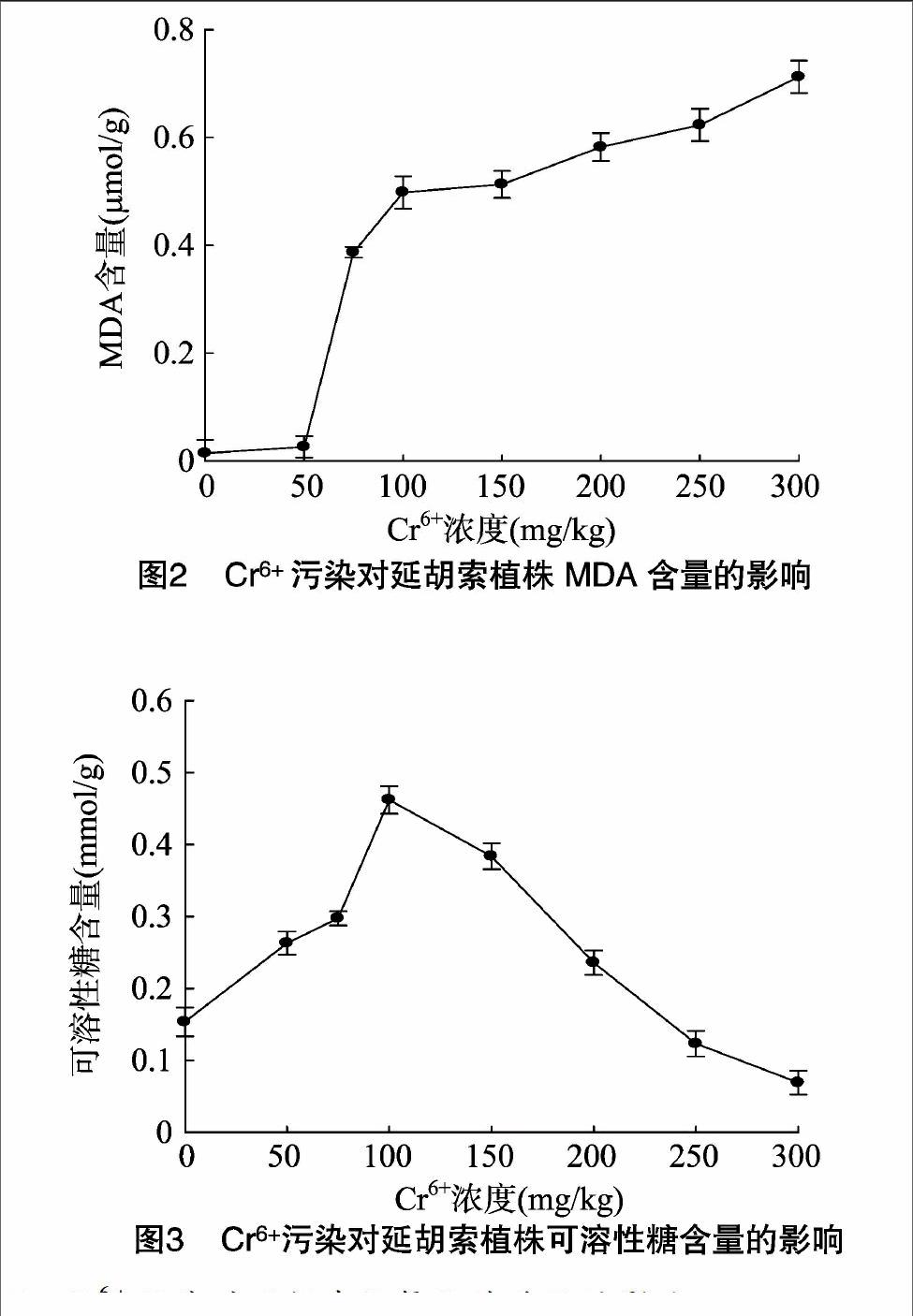
2.3Cr6+污染对延胡索MDA含量的影响
MDA是生物体内自由基作用于脂质发生过氧化反应所产生的氧化终产物,它会引起蛋白质、核酸等生物大分子的交联聚合,且具有细胞毒性,是植物膜系统受伤害的重要指标,其含量可反映膜脂过氧化的程度[14]。从图2可以看出,随着Cr6+浓度的不断升高,MDA含量总体呈上升趋势,在 300 mg/kg 时最大,与对照相比,增加了97.97%,表明了重金属胁迫大大提高了延胡索的膜脂过氧化水平,MDA含量在植物体内迅速聚集,表示植物膜系统受伤害越来越严重,植物的抗逆性减弱。
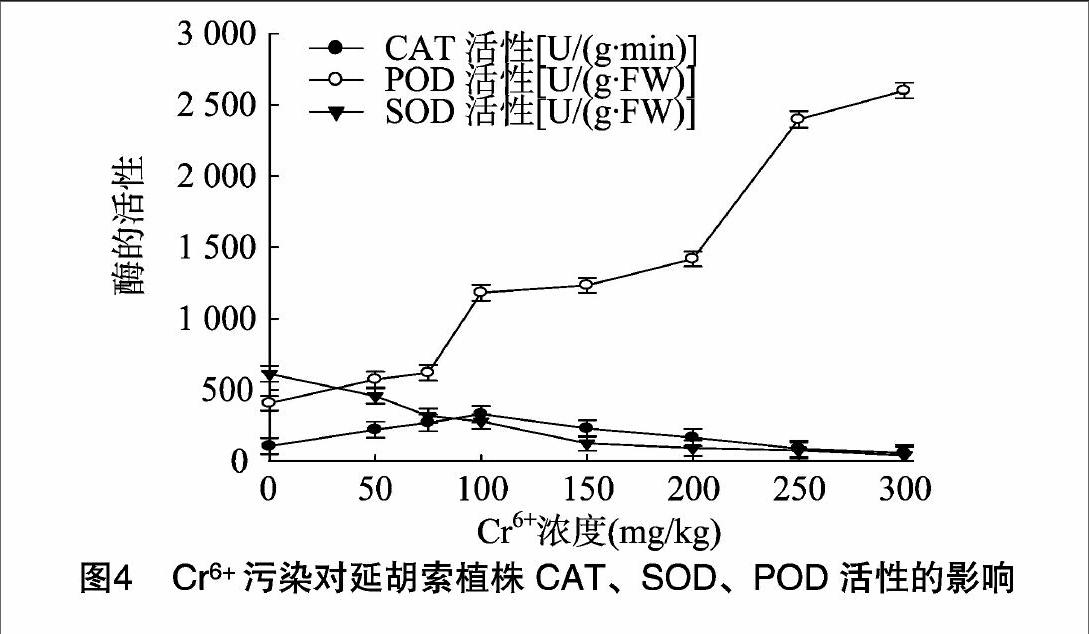
2.4Cr6+污染对延胡索可溶性糖含量的影响
可溶性糖具有调节渗透胁迫的作用,积累脯氨酸需要碳水化合物,脯氨酸的合成需要氧化还原能力,由碳水化合物通过氧化磷酸化作用提供,是植物代谢的基础物质。由图3可以看出,延胡索叶片细胞内可溶性糖含量随着Cr6+浓度的升高总体呈先升后下降趋势。延胡索叶片细胞内可溶性糖含量在Cr6+浓度为100 mg/kg时最大,比对照增加64.76%;随着Cr6+浓度的进一步升高,延胡索叶片细胞内可溶性糖含量逐渐降低,到300 mg/kg时最小,比对照减少了55.19%。这说明在Cr6+胁迫浓度较低时,Cr6+对其合成有一定的促进作用,高浓度Cr6+处理时,可溶性糖合成被抑制,对植物的生长发育有重大影响。
2.5Cr6+污染对延胡索抗氧化酶系统的影响
2.5.1Cr6+污染对延胡索SOD活性的影响植物体内存在抗氧化酶系统,由SOD、POD、CAT三者组成,若要有效清除植物细胞内的自由基和过氧化物,则需要它们三者的相互协调作用。清除细胞内的活性氧自由基由SOD酶完成。在逆境条件下,如重金属胁迫时,SOD酶能及时有效地清除植物细胞内产生的大量活性氧自由基,保护植物不被重金属胁迫。从图4中可以看出,SOD酶活性随着Cr6+浓度的升高呈下降的趋势,在Cr6+浓度为300 mg/kg时SOD活性最低,与对照相比,降低了93.10%。可能是在铬胁迫下植株SOD活性受到抑制,铬对延胡索叶片SOD活性毒害作用较大所致。
2.5.2Cr6+污染对延胡索POD活性的影响在植物叶片中,POD酶和SOD酶同等重要,其功能是将SOD酶的歧化产物双氧水分解成水。经研究发现,严重的逆境也可能增强POD的活性,因为逆境诱使植物产生更多的过氧化物。植物组织受到损害和破坏的程度越高,其POD活性越高。由图4可知,延胡索叶片的POD活性随着Cr6+浓度升高逐渐上升,在300 mg/kg时最大,与对照相比,增加了84.38%,这说明在重金属Cr6+胁迫作用下,随Cr6+浓度升高,植物叶片内的过氧化氢迅速积累,导致植株叶片内POD活性大幅度增加。
2.5.3Cr6+污染对延胡索CAT活性的影响CAT与SOD共同作用将H2O2分解为O2和H2O,催化能力很强。由图4可知,Cr6+浓度在小于100 mg/kg时延胡索叶片的CAT活性逐渐上升,在100 mg/kg时酶的活性最高,比对照增加了 68.09%,之后CAT活性下降,在Cr6+浓度为300 mg/kg时最低,减小了45.71%(与对照相比)。究其原因,低浓度Cr6+促进植株催化过氧化氢的生成量出现增加,CAT活性出现升高,CAT活性在Cr6+浓度>100 mg/kg后开始逐渐下降。在Cr6+浓度为300 mg/kg时酶的活性最低,此时Cr6+胁迫对植株表现出抑制作用,植物CAT活性降低。
2.6Cr6+污染对延胡索重金属含量的影响
由图5可知,随着Cr6+浓度的升高,延胡索地下部分Cr6+积累量增加,各处理间差异明显,并且浓度越大,延胡索地下块茎Cr6+增长的幅度也越大,积累的量越大。在Cr6+浓度为150 mg/kg时,Cr6+含量增加速度最大,达到 87.23 mg/kg,比Cr6+浓度为100 mg/kg时增加了91.95%;Cr6+浓度为300 mg/kg时,Cr6+积累量为93.04 mg/kg,比Cr6+浓度为250 mg/kg增加了3.62%。
3讨论与结论
3.1Cr6+ 污染对延胡索生长的影响
Cr6+污染影响延胡索的生长。本研究发现,Cr6+污染降低延胡索的生物量和产量。随着Cr6+处理浓度的升高,延胡索块茎的生物量呈现出递减趋势,Cr6+污染浓度200 mg/kg对延胡索生长和产量有明显的影响。鲜质量和干质量减少幅度突然增加,鲜质量由Cr6+污染浓度150 mg/kg时的 28.40% 增加到200 mg/kg时的54.47%,干质量由 150 mg/kg 的30.20%增加到200 mg/kg的65.77%(均和对照相比)。
3.2Cr6+污染对延胡索生理特性的影响
延胡索叶绿素含量能被Cr6+污染明显影响。绿色植物进行光合作用的主要色素对植物的生长发育有着重要作用,其含量在一定程度上能反映植物同化物质的能力。许多试验结果表明,绿色植物的光合作用在重金属胁迫下都表现为抑制作用,且抑制效果与胁迫的程度成正相关,另外光合抑制也与植物种类和发育时期以及重金属的种类有关[15]。本研究发现,Cr6+浓度小于100 mg/kg时,叶绿素(a+b)含量随Cr6+浓度逐渐降低而减少。下降原因可能是Cr6+进入延胡索叶片细胞内,使叶绿体酶活性失调,加速叶绿素分解,同时由于Cr6+局部积累较多,与叶绿体中蛋白质上-SH结合等使叶绿素蛋白中心离子组成发生变化而失和。如果重金属Cr6+浓度够高,植物不能进行光合作用得到足够的养分,会出现生长缓慢,甚至会停止生长,其生物量降低。MDA是一种高活性的膜脂过氧化产物,其含量的多少能反映组织细胞膜损伤程度,衰老和逆境引起组织中MDA含量的增加量与细胞膜损伤程度呈正相关,而叶绿素含量与MDA含量呈显著负相关[16]。本研究也获得了同样的结论,并且叶绿素含量与MDA含量也呈现明显的负相关.由于MDA为高活性的膜脂过氧化物,通过交联脂类、核酸、糖类及蛋白质可影响质膜和叶绿体片层膜的结构和功能,影响膜的流动性及其与酶的结合力,从而影响位于片层上的叶绿素含量[17]。重金属Cr6+对可溶性糖含量有影响,可溶性糖含量随着重金属Cr6+浓度的升高而先升后降,可溶性糖是小分子物质,具有调节渗透胁迫的作用,可溶性糖含量的减少表明渗透调节胁迫系统紊乱,氧化还原增强。
植物抗氧化酶系统由SOD、POD和CAT共同构成,活性氧的生成和代谢受其活性直接影响[18]。高活性抗氧化酶有利于提高植物对各种生物及非生物胁迫的耐受性[19]。本研究表明,低浓度Cr6+处理,对延胡索的POD活性和CAT活性均有增加作用,而高浓度Cr6+处理则表现抑制作用,导致CAT活性降低,而POD酶活性一直增加。可能低浓度的Cr6+对CAT活性有一定的刺激作用,这说明CAT对低浓度Cr6+有耐受作用,能在一定范围内催化有毒物质的分解,但是超过一定的范围,依然会破坏CAT的活性。POD酶活性一直上升,说明在重金属Cr6+胁迫作用下,随Cr6+浓度的升高,植物叶片内的过氧化氢迅速积累,导致植株叶片内POD活性大幅度增加。但SOD活性随着Cr6+浓度的升高逐渐降低,低于对照处理。这说明Cr6+对植株叶片SOD活性毒害作用较大,植株SOD活性受到抑制明显。这一结果暗示保护酶系统在高浓度Cr6+胁迫下对活性氧的清除能力大大削弱,自由基在细胞内的积累已不能阻止,使膜发生膜脂过氧化,其过氧化产物MDA产生并积累,膜透性增加,导致膜功能受损,从而引起细胞的衰老和死亡,对植物产生抑制效应甚至毒害作用。
本研究结果表明,延胡索的生长对Cr6+污染有明显的影响,延胡索在种植时要注意土壤中Cr6+含量,延胡索栽培中应尽量选择Cr6+污染较少的土地进行。
参考文献:
[1]Gu G W,Zhang H W. Trace elements and malignant tumor[M]. Shanghai:Shanghai Science and Technology Press,1993:199-205.
[2]Sharma D C,Sharma C P,Tripathi R D. Phytotoxic lesions of chromium in maize[J]. Chemosphere,2003,51(1):63-68.
[3]Choudhury S,Panda S K. Toxic effects,oxidative stress and ultrastructural changes in moss Taxithelium nepalense (Schwaegr.) Broth. under chromium and lead phytotoxicity[J]. Water Air and Soil Pollution,2005,167(1/2/3/4):73-90.
[4]Pandey V,Dixit V,Shyam R. Antioxidative responses in relation to growth of mustard (Brassica juncea cv. Pusa Jaikisan) plants exposed to hexavalent chromium[J]. Chemosphere,2005,61(1):40-47.
[5]Sinha S,Gupta A K,Bhatt K. Uptake and translocation of metals in fenugreek grown on soil amended with tannery sludge:involvement of antioxidants[J]. Ecotoxicology and Environmental Safety,2007,67(2):267-277.
[6]郑爱珍. 重金属Cr6+污染对辣椒幼苗生理生化特性的影响[J]. 农业环境科学学报,2007,26(4):1343-1346.
[7]李廷真,付川,余顺慧,等. Cu2+胁迫下三峡库区消落带的狗牙根生长状况及其铜富积效应[J]. 贵州农业科学,2013,41(7):184-186.
[8]余顺慧,来守军,王绍恒,等. 铜胁迫对三峡库区消落带适生植物狗牙根生理特性和铜积累的影响[J]. 西南大学学报:自然科学版,2013,38(11):58-63.
[9]余顺慧,黄怡民,潘杰,等. 铜胁迫对2种三峡库区消落带适生植物生长及铜积累的影响[J]. 西南农业学报,2014,27(3):1196-1201.
[10]潘杰,来守军,黄怡民,等. 铅胁迫对2种三峡库区消落带适生植物生长及铅积累的影响[J]. 江苏农业科学,2014,42(6):332-335.
[11]郝建军. 植物生理学试验技术[M]. 沈阳:辽宁科学技术出版社,2004.
[12]赵世杰,许长成,邹琦,等. 植物组织中丙二醛测定方法的改进[J]. 植物生理学通讯,1994,30(3):207-210.
[13]李合生. 植物生理生化试验原理和技术[M]. 北京:高等教育出版社,2000.
[14]李志刚,梅利民,万雪芹,等. 低浓度重金属Cr6+对紫狼尾草生理生化特性的影响[J]. 广西农业科学,2009,40(5):478-482.
[15]江行玉,赵可夫. 植物重金属伤害及其抗性机理[J]. 应用与环境生物学报,2001,7(1):92-99.
[16]严明理,冯涛,向言词,等. 铀尾沙对油菜幼苗生长和生理特征的影响[J]. 生态学报,2009,29(8):4215-4222.
[17]徐振柱,于振文,董庆裕,等. 水分胁迫对冬小麦旗叶细胞质膜及叶肉细胞超微结构的影响[J]. 作物学报,1997,23(3):370-375.
[18]韩金龙,李慧,蔺经,等. 核黄素对盐胁迫下杜梨叶片抗氧化系统的影响[J]. 江苏农业学报,2015,31(4):893-898.
[19]Vannini C,Iriti M,Bracale M A,et al. The ectopic expression of the rice Osmyb4 gene in Arabidopsis increases tolerance to abiotic,environmental and biotic stresses[J]. Physiological and Molecular Plant Pathology,2006,69(1/2/3):26-42.陈璐,苏家乐,李畅,等. 贮藏时间、温度及PEG胁迫对云南杜鹃种子萌发的影响[J]. 江苏农业科学,2016,44(10):240-242.
猜你喜欢
天然产物研究与开发(2018年11期)2018-11-30
中成药(2017年9期)2017-12-19
山东农业科学(2016年12期)2017-01-21
现代园艺(2016年9期)2016-07-16
西南农业学报(2016年5期)2016-05-17
现代园艺(2016年4期)2016-03-24
现代园艺(2016年2期)2016-03-15
河北医药(2013年3期)2013-10-10
