
大菱鲆胚胎生长发育的研究
2017-02-05 15:27佟雪红鲍成满汤新慧杨小兰
江苏农业科学 2016年10期
佟雪红++鲍成满++汤新慧++杨小兰++陈倩

doi:10.15889/j.issn.1002-1302.2016.10.092
摘要:观察了大菱鲆胚胎的发育过程,研究了其发育特征和胚体的生长特征。结果表明,大菱鲆受精卵在水温为14.4 ℃、盐度为3.1%~3.3%条件下,历经103 h胚体孵化出膜。胚体长度在卵裂期、囊胚期、原肠期、器官形成期经历了“缩小-膨大-再缩小-再膨大”的变化过程,并在卵裂期达到最低值1.015 mm,胚体大小的变化跟不同发育期的细胞运动以及器官发育密切相关。胚体中油球直径从受精卵至卵裂期逐渐增加并在卵裂期达到最高值 0.186 mm,之后逐渐减小。胚体质量在卵裂期时最低,为0.509 mg,之后快速增长并在原肠期达到最高值0.576 mg,此后急剧降低,在孵化时胚体质量为0.525 mg。
关键词:大菱鲆;胚胎发育;生长;油球;体质量
中图分类号: S961.2文献标志码: A文章编号:1002-1302(2016)10-0318-03
收稿日期:2015-08-29
基金项目:国家自然科学基金(编号:31340007);国家大学生创新创业训练计划项目;江苏省高校自然科学基础研究项目(编号:12KJB240001);江苏省滩涂生物资源与环境保护重点建设实验室开放课题(编号:JLCBE12003);盐城师范学院教授、博士基金(编号:6207110006)。
作者简介:佟雪红(1979—),女,江苏盐城人,博士,讲师,研究方向为鱼类生理学。E-mail:txh113005@163.com。大菱鲆(Scophthalmus maximus)属鲆科(Bothidae)菱鲆属(Scophthalmus),在中国又称“多宝鱼”,自然分布于东北大西洋沿岸。它具有适应低水温生活、生长速度快、肉质好、养殖和市场潜力大等优点,是名贵的低温经济鱼类,现已成为我国北方沿海重要的养殖品种[1]。
关于大菱鲆早期发育变化规律的研究已有一些报道。雷霁霖等对大菱鲆胚胎和仔鱼、稚鱼发育进行了详细阐述[2];李庆华等采用营养强化和控光控温方法对大菱鲆亲鱼性腺发育及卵子质量的影响进行了研究[3];彭墨等用亚麻籽油替代鱼油对大菱鲆幼鱼生长、脂肪酸组成及脂肪沉积的影响进行了探讨[4];苗淑彦等探究了饲料中不同种类的碳水化合物对大菱鲆幼鱼生长、成活、饲料利用、体组成和血液生理生化指标等的影响[5];Al-Maghazachi等对大菱鲆仔鱼、稚鱼的发育进行了分期研究[6];van Bussel等研究了硝酸盐对大菱鲆幼鱼的生长生理的影响[7]。上述研究主要围绕大菱鲆早期发育的形态学和消化生理学而展开,而关于大菱鲆胚胎发育阶段的生长特性的研究未见相关报道。本试验在观察记录了大菱鲆胚胎发育时序的基础上,进一步测定了胚体的长度、油球的直径和胚体的体质量,分析了胚体的生长特性,以期为完善大菱鲆的人工育苗技术提供基础资料。
1材料与方法
1.1材料
试验于2014年4月在山东莱州大菱鲆育苗场进行。试验用雌、雄亲鱼均取自亲鱼驯养池,将亲鱼人工催产后进行受精,将品质较好的浮性卵移入孵化网箱进行孵化培育。
1.2方法
1.2.1胚胎发育的观察胚胎孵化过程中水温为(14.4±05) ℃,盐度为3.1%~3.3%,pH值为7.9~8.3。在不同发育期连续采样,并用奥林巴斯显微镜(BX41)观察胚胎的发程、记录发育特征,直至孵化出膜。
1.2.2胚胎生长的测定每个发育期随机取样70粒受精卵,其中30粒用于测定胚胎的长度和油球的直径,采用与显微镜配套的电子目镜进行测量;另外40粒受精卵在吸干水分后测定其质量,用电子天平进行测量。
2结果与分析
2.1大菱鲆胚胎发育时序及形态特征
由表1可知,受精后2 h 40 min进入卵裂期,9 h 40 min时胚体出现卵黄合胞体;10 h 40 min进入囊胚期;17 h 10 min进入原肠期,36 h原口关闭前期,胚盘下包80%,体节、视囊原基、克氏囊出现,39 h 10 min原口关闭,胚体成型,尾芽形成;47 h 50 min 胚体克氏囊明显,听囊出现、色素形成,51 h 30 min胚体色素增多,晶体原基形成,克氏囊开始退化,86 h胚体心跳出现,角膜形成,尾部鳍褶出现;103 h时胚体头部破膜而出。
2.2大菱鲆胚胎大小变化
大菱鲆胚胎生长过程中,其长度整体呈增长趋势。但不同发育期胚体长度经历“缩小—膨大—再缩小—再膨大”的变化过程。由图1可知,胚体长度从受精卵到卵裂期呈缩小趋势并在卵裂期达到最低值(1.015±0.121) mm,之后保持膨大趋势直至囊胚期,囊胚期至原肠期再次出现缩小趋势并在原肠期时达到(1.044±0.241) mm,之后保持膨大状态直至孵化。
2.3大菱鲆胚体油球变化
如图2所示,油球直径从受精卵至卵裂期逐渐增加并在卵裂期达到最高点(0.186±0.011) mm,之后逐渐缩短。油球直径在卵裂期至原肠期间减少的幅度[(0.186±0.011)~表1大菱鲆胚胎发育过程及主要特征
发育时期发育阶段受精后时间胚体发育及主要特征受精卵受精0 h质量较好的受精卵无色透明,漂在水面上胚盘出现2 h原生质在动物极形成胚盘卵裂期2细胞2 h 40 min胚盘出现2个均等的分裂球4细胞3 h 10 min胚盘出现4个均等的分裂球8细胞3 h 40 min胚盘出现大小不均等的8个分裂球16细胞4 h 40 min胚盘出现16个分裂球,呈双层分布,里层4个细胞,外层12个细胞32细胞5 h 40 min胚盘出现不均等、不规则的32个分裂球64细胞6 h 50 min胚盘出现不规则的64个分裂球,最外侧分裂球界限模糊连成一体128细胞7 h 30 min分裂球增为128个,细胞体积小、不易分辨多细胞9 h 40 min胚盘中细胞数量越来越多,边界变得模糊不清,中央部分的细胞连成一体,出现与营养物质吸收有关的卵黄合胞体囊胚期高囊期10 h 40 min动物极的胚盘高度较高,卵黄合胞体层分化为内、外两部分低囊期11 h 50 min胚盘中细胞沿卵黄囊迁移,高度变低原肠期原肠早期17 h 10 min胚盘中分裂细胞沿卵黄迁移,开始下包;边缘细胞会内卷并向上运动,在囊胚层外周形成胚环。部分分裂细胞流向胚环的某部并加厚形成胚盾原肠早期24 h胚盘下包1/5,胚盾继续加厚,并形成胚体原基原肠中期29 h 30 min胚盘下包1/2;胚盾继续延长,胚体形态明显原肠中期32 h 10 min胚盘下包3/4;胚体的头突出现并逐渐增大;表皮、神经褶出现原肠中期36 h胚盘下包4/5;胚环逐渐缩小为胚孔,而未被完全包被的卵黄被称为原口;体节、视囊原基、克氏囊出现原肠晚期39 h 10 min胚盘下包完全,原口封闭,胚体成型;脊索原基形成、尾芽形成器官形成期听囊形成期47 h 50 min胚体克氏囊明显,听囊出现、色素形成晶体形成期51 h 30 min胚体色素增多,消化道、晶体原基形成,克氏囊开始退化体节形成期56 h 50 min胚体克氏囊消失,体节分节,肌节逐步形成肌节分化期60 h 40 min胚体绕卵黄2/3,背部色素增多,胚胎肌节分节明显,头部晶体体积增大心脏分化期65 h胚体绕卵黄3/4,心脏进一步发育并分化鳍褶形成期72 h 30 min胚体绕卵黄4/5,透明的鳍褶在尾部生成并向头部延伸鳍原基形成期79 h 35 min胚体扭动加快,脉络膜裂形成,脊索中空化,腹部出现鳍原基心脏跳动期86 h胚体心跳出现,角膜形成,尾部鳍褶出现肌肉效应期95 h 40 min胚体绕卵黄9/10,扭动剧烈,红色色素明显增多,尾部出现黑色色素孵化期破膜期103 h胚体剧烈扭动并孵化出膜,仔鱼背鳍和臀鳍上出现绒毛状色素丛,鱼体呈深棕色并携带卵黄囊
(0.184±0021) mm]小于原肠期至孵化期间减少的幅度[(0.184±0021)~(0.180±0.009) mm]。
结合图1和图2的数据,胚体长度与油球直径的比值在不同发育期差异较大。比值在卵裂期时最低,为5.46,其后该比值整体呈现升高状态并且在器官形成期和孵化期分别达到5.88和5.98,较受精卵时期增幅分别为4.57%和6.26%。
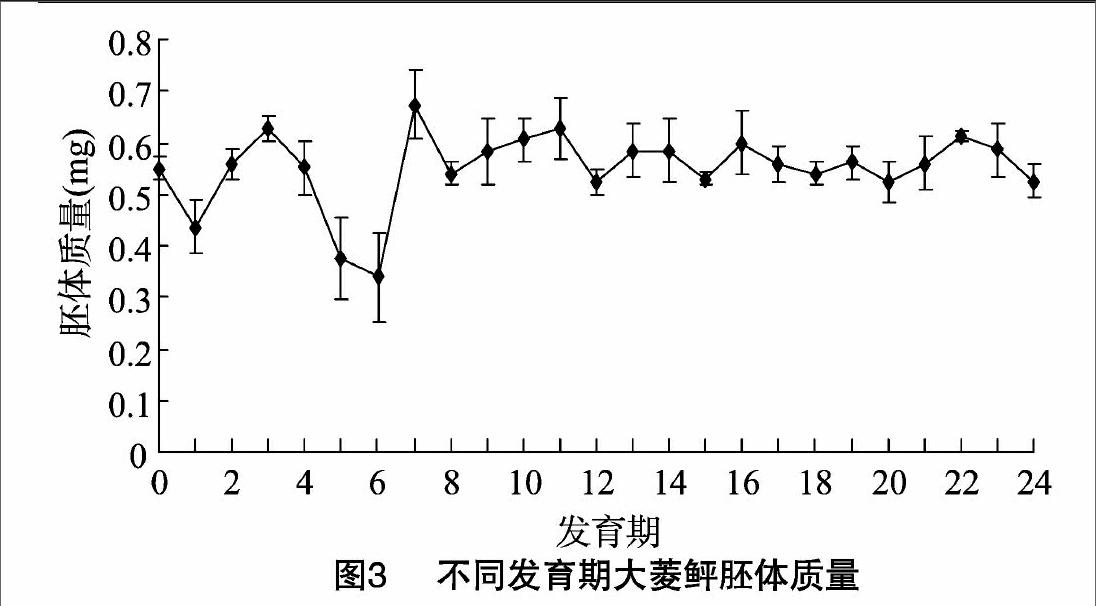
2.4大菱鲆胚胎体质量变化
胚体质量在卵裂期最低,为(0.509±0073) mg;之后快
速增长并在原肠期达到最高值(0.576±0.068) mg;此后急剧降低,在孵化时胚体质量为(0.525±0106) mg,较原肠期降幅为8.85%(图3)。
3讨论
3.1大菱鲆胚胎发育特征
大菱鲆受精卵呈圆形,浮性卵。不同发育期历时时间差异较大,其中卵裂期7 h,囊胚期1 h 10 min,原肠期22 h,器官形成期47 h 50 min,即胚体器官原基形成以及功能强化过程
经历的时间最长,这与其他生物早期发育过程规律一致[8-9]。胚体的这种发育机制为完善机体生理功能、提高胚后阶段仔稚鱼的成活率提供了保障。
温度是影响鱼类胚胎发育速度的重要因子,不同温度条件下胚体出膜的时间差异较大。雷霁霖等研究表明,在 13 ℃ 条件下,大菱鲆胚胎约经116 h完成孵化[2]。而本试验中,水温为14.4 ℃,胚胎历时103 h孵化出膜。上述数据说明,胚胎发育跟水温密切相关,在一定范围内,水温越高,发育速度越快;水温越低,发育速度越慢。类似的结果也出现在半滑舌鳎[10]、对虾[11]、黄鲷[12]、扇贝[13]、乌贼[14]等生物的胚胎发育过程中。但孵化时水温也不能过高,水温过高会降低孵化率[15]。此外,有报道称,受精卵的分裂速度受细胞质影响[16],由此推测,可能是水温的变化首先影响到细胞质的理化性质,进而影响到胚胎的卵裂速度和孵化成功率。另外,大菱鲆胚胎发育速度不同还可能与试验对象分布在不同地理环境导致的种群差异以及胚体内DNA含量、卵径、卵黄成分等方面的差异有关[17]。
3.2大菱鲆胚胎生长特征
近年来,关于大菱鲆胚胎发育的研究已有报道[1-2,18],但是关于胚体发育期胚体生长的研究鲜有报道。本试验精确测定了胚胎的大小和体质量,结果表明,大菱鲆胚胎生长过程中,胚体长度经历“缩小—膨大—再缩小—再膨大”的变化过程。有研究报道表明,卵膜的收缩和膨胀跟胚体的细胞运动以及器官发育密切相关[19]。在本试验中,胚体大小跟发育时期密切相关,在细胞数量急剧增多的卵裂期和变化剧烈的原肠期,胚体长度均出现缩小的现象,而在其他时期表现为增大的趋势,原因可能是在发育的最后阶段,胚体通过吸收大量水分来增加自身体积,进而增大胚体内的压力,以便帮助胚体冲破胚膜,顺利孵出。
大菱鲆的卵子属于端黄卵,有1个油球。油球中营养丰富,含有脂类和脂肪酸等营养物质,是大菱鲆胚胎发育必需的营养和能量来源。不同发育时期,大菱鲆胚体中油球直径差异较大,卵裂期之前油球直径逐渐增大,卵裂期之后逐渐降低。此外,胚体长度与油球直径的比值也呈现了类似的变化趋势:该比值在卵裂期时最低,为5.46,其后该比值整体呈现升高状态并且在器官形成期和孵化期分别达到5.88和 5.98。卵裂期之后,油球直径持续下降的趋势说明,原肠期和器官形成期,尤其是器官形成期,胚体对油球中脂类和脂肪酸等营养物质的利用比率逐渐增加,这与上述时期细胞活动和组织分化剧烈导致的耗能量增大有关[20]。
胚胎的体质量跟发育期也联系密切,在卵裂期时体质量达到最低值,可能是与胚体的迅速生长和旺盛的新长代谢导致的内源营养消耗量升高有关;在原肠期时,胚体质量达到最高峰,但原肠期之后,胚体质量一直呈下降趋势直至孵化出膜,具体的原因尚有待于进一步深入研究分析。
参考文献:
[1]雷霁霖,刘新富. 大菱鲆引进养殖的初步研究[J]. 现代渔业信息,1995,10(11):1-3.
[2]雷霁霖,马爱军,刘新富,等. 大菱鲆(Scophthalmus maximus L.)胚胎及仔稚幼鱼发育研究[J]. 海洋与湖沼,2003,34(1):9-18.
[3]李庆华,孙建,李仰真,等. 营养强化和控光控温对大菱鲆亲鱼性腺发育及卵子质量的影响[J]. 南方农业学报,2013,44(6):1030-1035.
[4]彭墨,徐玮,麦康森,等. 亚麻籽油替代鱼油对大菱鲆幼鱼生长、脂肪酸组成及脂肪沉积的影响[J]. 水产学报,2014,38(8):1131-1139.
[5]苗淑彦,苗惠君,聂琴,等. 饲料中不同种类的碳水化合物对大菱鲆生长性能和代谢反应的影响[J]. 水产学报,2013,37(6):910-919.
[6]Al-Maghazachi S J,Gibson R. The developmental stages of larval turbot,Scophthalmus maximus (L.)[J]. Journal of Experimental Marine Biology and Ecology,1984,82(1):35-51.
[7]van Bussel C G J,Schroeder J P,Wuertz S,et al. The chronic effect of nitrate on production performance and health status of juvenile turbot (Psetta maxima)[J]. Aquaculture,2012,326-329:163-167.
[8]胡振禧,黄洪贵,吴妹英,等. 福建地区斑鳜胚胎与仔鱼早期发育的研究[J]. 上海海洋大学学报,2014,23(2):193-199.
[9]殷名称. 鱼类早期生活史研究与其进展[J]. 水产学报,1991,15(4):348-358.
[10]张鑫磊,陈四清,刘寿堂,等. 温度、盐度对半滑舌鳎胚胎发育的影响[J]. 海洋科学进展,2006,24(3):342-348.
[11]杨世平,王成桂,黄海立,等. 环境温度和盐度对墨吉明对虾(Fenneropenaeus merguiensis)胚胎发育的影响[J]. 海洋与湖沼,2014,45(4):817-822.
[12]夏连军,施兆鸿,王建钢,等. 温度对黄鲷胚胎发育的影响[J]. 上海水产大学学报,2006,15(2):163-168.
[13]陈来钊,王子臣. 温度对海湾扇贝与虾夷扇贝及其杂交受精,胚胎和早期幼虫发育的影响[J]. 大连水产学院学报,1994,9(4):1-9.
[14]陈道海,王雁,梁汉青,等. 虎斑乌贼(Sepia pharaonis)胚胎发育及孵化历期观察[J]. 海洋与湖沼,2012,43(2):394-400.
[15]高养春,李雅娟,姜志强,等. 泥鳅杂交三倍体和二倍体胚胎发育的比较研究[J]. 大连海洋大学学报,2014,29(2):103-108.
[16]夏仕玲,余来宁,吴作文. 鱼类卵细胞质对胚胎发育速度的调控[J]. 淡水渔业,1990(3):17-18.
[17]蔡明夷,刘贤德,陈庆凯,等. 温度对大黄鱼受精卵卵裂间隔时间的影响[J]. 集美大学学报:自然科学版,2010,15(3):161-164.
[18]Devauchelle N,Alexander J C. Spawning of turbot (Scophthalmus maximus) in capativity[J]. Aquaculture,1988,69(1/2):159-184.
[19]蒋霞敏,唐锋,罗江,等. 拟目乌贼的胚胎发育[J]. 水产学报,2013,37(5):711-718.
[20]谢菁,区又君,李佳儿,等. 七带石斑鱼胚体和卵黄囊期仔鱼的发育[J]. 海洋通报,2009,28(2):41-49.
猜你喜欢
黑龙江水产(2022年6期)2022-12-03
食品工业(2021年9期)2021-04-16
小读者(2021年2期)2021-03-29
疯狂英语·新悦读(2019年11期)2019-12-18
华人时刊(2019年13期)2019-11-17
中国水产(2019年3期)2019-03-25
NBA特刊(2018年21期)2018-11-24
海洋科学(2017年10期)2017-02-09
浙江农业学报(2016年7期)2016-06-15
中国海洋大学学报(自然科学版)(2014年9期)2014-02-28
