
饲料蛋白质和脂肪水平对亚东鲑亲鱼生长性能、消化酶活性和血清指标的影响
2017-02-28 05:04王常安国孙王炳谦徐奇友
动物营养学报 2017年2期
王常安 户 国孙 鹏 谷 伟 王炳谦徐奇友
(中国水产科学研究院黑龙江水产研究所,哈尔滨 150070)
饲料蛋白质和脂肪水平对亚东鲑亲鱼生长性能、消化酶活性和血清指标的影响
王常安 户 国∗孙 鹏 谷 伟 王炳谦∗徐奇友
(中国水产科学研究院黑龙江水产研究所,哈尔滨 150070)
本试验旨在研究饲料蛋白质和脂肪水平对亚东鲑亲鱼生长性能、性腺指数、消化酶活性和血清指标的影响。试验采用5×2两因素随机设计,设5个蛋白质水平(36%、39%、42%、45%、48%)和2个脂肪水平(9%、18%),共配制10种实用试验饲料。每种试验饲料投喂3个重复,每个重复19尾鱼。初重为(462.53±45.40)g亚东鲑在室内水族箱中流水饲养,饲养周期为77 d。结果表明:同一脂肪水平下,随饲料蛋白质水平的升高,增重率有所升高,而饲料系数逐渐降低,至蛋白质水平为48%时又有所升高。饲料蛋白质和脂肪水平对雌性亲鱼性腺指数无显著影响(P>0.05),雄性亲鱼性腺指数随蛋白质水平的升高而降低,但至蛋白质水平为48%时又有所回升。同一脂肪水平下,随着饲料蛋白质水平的升高,胃、肠道和幽门盲囊的脂肪酶活性逐渐增加,蛋白质水平为48%时显著高于蛋白质水平为36%时(P<0.05);而胃、肠道和幽门盲囊的蛋白酶活性虽有所增加,但各组间无显著差异(P>0.05)。同一蛋白质水平下,18%脂肪水平组胃、肠道和幽门盲囊的蛋白酶和脂肪酶活性均略高于9%脂肪水平组。同一脂肪水平下,血清氨含量随饲料蛋白质水平的升高而逐渐增加。饲料蛋白质和脂肪水平对血清超氧化物歧化酶、溶菌酶活性以及葡萄糖、甘油三酯、总蛋白、白蛋白和丙二醛含量均未产生显著影响(P>0.05)。由此得出,亚东鲑亲鱼对饲料蛋白质(36%~48%)和脂肪(9%~18%)水平有一定的适应性反应;以增重率为指标进行折线模型分析,得出饲料脂肪水平为9%和18%时,亚东鲑亲鱼对蛋白质需要量分别为43.11%和45.69%。
亚东鲑亲鱼;蛋白质;脂肪;生长;消化酶;血清指标
亲鱼营养的研究对亲鱼饲料的配制、优质苗种的培育等具有重要的实践意义。目前,亲鱼营养的知识十分有限,主要是由于研究的难度较大,需要较多的养殖设施,且投喂试验需要花费大量的时间和经费[1-2]。亲鱼的营养与早期发育阶段有所不同,且亲鱼的营养水平直接影响其子代的营养状况。目前已经确定一些营养素如必需脂肪酸对维持亲鱼的生长和繁殖性能十分重要,但过量或不平衡会严重影响其繁殖性能。然而,关于大多数营养素如微量元素和维生素等对亲鱼影响的资料还比较缺乏。
褐鳟(Salmo trutta)是鲑科鱼类中重要的经济品种之一,其肉质鲜美、营养丰富、经济价值较高,国内仅在西藏亚东地区有分布,称为亚东鲑(Salmo trutta fario),系与褐鳟同物异名种[3]。国外对褐鳟的研究报道较多,主要集中在生物学等方面[4-5]。目前,国内对亚东鲑的养殖生物学研究资料较少,主要集中于分子生物学、繁殖生物学、养殖方式等方面[6-9],尚无关于其营养学方面的研究,亦未有专门的配合饲料,而用其他鱼类的饲料投喂往往导致亚东鲑生长性能低下。为此,很有必要研究亚东鲑的高效饲料,以提高其生长性能,改善其品质。饲料中适宜的蛋白质和脂肪水平是鱼类健康生长和发育的前提条件,也是配合饲料中需优先确定的2个重要参数,因此,本研究探讨饲料蛋白质和脂肪水平对亚东鲑亲鱼生长性能、性腺指数、消化酶活性、血清生化指标和非特异性免疫力的影响,旨在为研制亚东鲑亲鱼配合饲料提供理论依据。
1 材料与方法
1.1 试验动物
初重为(462.53±45.40)g的亚东鲑由中国水产科学研究院黑龙江水产研究所渤海冷水鱼试验站提供。
1.2 生长试验
本试验采用5×2两因素完全随机设计,设5个蛋白质水平(36%、39%、42%、45%、48%)和2个脂肪水平(9%、18%),共配制10种实用试验饲料(P36/L9、P39/L9、P42/L9、P45/L9、P48/L9、P36/L18、P39/L18、P42/L18、P45/L18、P48/L18),每种试验饲料投喂3个重复,每重复19尾鱼。试验饲料组成及营养水平见表1。试验鱼以重复为单位在室内玻璃钢水族箱(直径90 cm,水深45 cm)中流水饲养,以商品饲料驯化14 d后进行养殖试验。试验开始前,试验鱼饥饿24 h后用苯氧乙醇(0.5 mL/L)麻醉后称量体重和体长。试验水为涌泉水,水温11.5~17.5℃,pH 7.2~7.5,溶氧浓度7.8~10.0 mg/L,氨氮浓度<0.2 mg/L。光周期为15 h明:9 h暗。日投喂2次,投饲率为体重的1.0%,以饱食而没有剩料为原则,每次投喂结束40 min后收集残饵,饲养期为77 d。
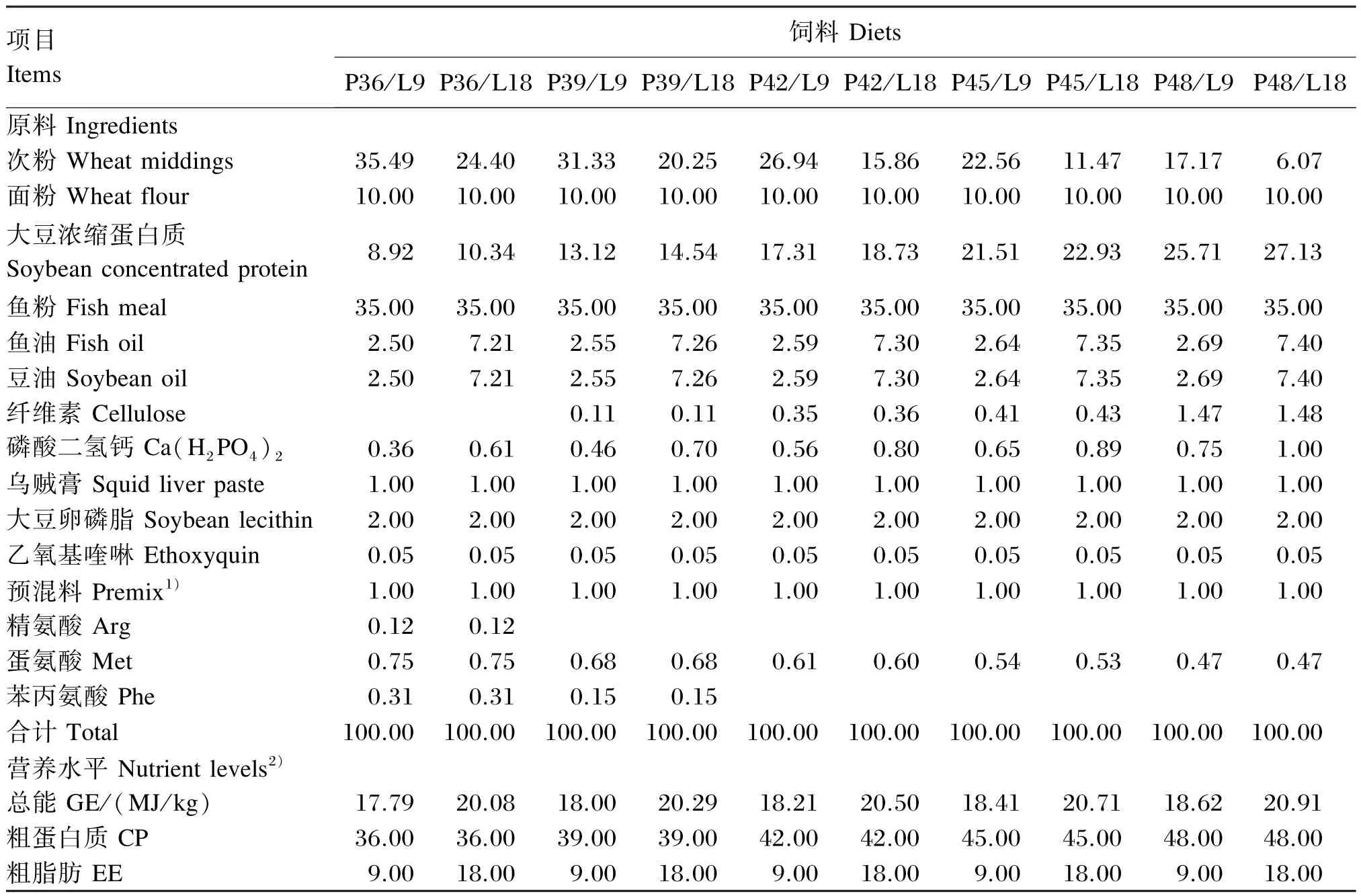
表1 试验饲料组成及营养水平(风干基础)Table 1 Composition and nutrient levels of experimental diets(air-dry basis) %
1.3 指标测定和方法
1.3.1 生长性能和性腺指数
饲养期结束后,试验鱼饥饿24 h,统计养殖周期内试验鱼死亡数和每个水族箱试验鱼饲料摄入量,称量每个水族箱试验鱼总重。每个水族箱随机取6尾鱼,解剖鱼体,分离性腺,鉴定雌雄,并称重,取其整个内脏团并称重。根据以下公式计算试验鱼的生长性能指标和性腺指数:

式中:W0为初重(g);Wt为终重(g);Nf为终末尾数;Ni为初始尾数;Wg为性腺重(g);Wq为去内脏体重(g)。
1.3.2 消化酶活性
每个水族箱随机取4尾鱼,于冰盘上分别取胃、肠道和幽门盲囊,0.86%预冷生理盐水洗净,滤纸吸干,称重。胃、肠道和幽门盲囊分别按1∶9的重量体积比加入 0.86%预冷生理盐水,以 FJ-200CL高速组织匀浆机匀浆(15 000 r/min,3 min)稀释,4℃条件下4 000 r/min离心10 min,取上清液放入1.5 mL离心管中,-80℃冷冻保存备测消化酶活性。蛋白酶活性采用福林-酚法测定,淀粉酶活性采用淀粉-碘比色法测定,脂肪酶活性采用聚乙烯醇橄榄油乳化液水解法测定,组织蛋白质含量采用考马斯亮蓝法测定[10]。蛋白酶活性的定义:在 37℃时,每分钟水解酪素产生1 μg酪氨酸为1个酶活性单位。脂肪酶活性的定义:在37℃时,脂肪酶水解脂肪每分钟产生1 μg分子脂肪酸所消耗的酶量为1个酶活性单位。淀粉酶活性的定义:在 37℃ 时,在 30 min内,100 mL酶液中的淀粉酶能完全水解10 mg淀粉时为1个酶活性单位。酶活性单位用每克组织蛋白质的酶活性单位(U/g prot)表示。
1.3.3 血清指标
每个水族箱随机取4尾鱼,尾柄采血,4℃冰箱中静置30 min,3 000 r/min离心15 min,取血清置于1.5 mL离心管中,-80℃冷冻保存。血清总蛋白(total protein,TP)、白蛋白(albumin,ALB)、甘油三酯(glycerinate,TG)和葡萄糖(glucose,GLU)含量采用美国贝克曼库尔特有限公司生产的试剂盒以全自动生化分析仪(贝克曼ProCX4,美国)测定,其中TP和ALB含量采用化学法测定,球蛋白(globulin,GLB)含量为二者的差值,TG和GLU含量采用酶法测定。血清氨(ammonia,AMM)、丙二醛(malondialdehyde,MDA)含量及超氧化物歧化酶(superoxide dismutase,SOD)、溶菌酶(lysozyme,LZM)活性采用南京建成生物工程研究所生产的试剂盒测定。酶活性单位定义:在37℃恒温条件下,每毫升反应液中,每分钟抑制邻苯酚自氧化率达50%的酶量定义为1个超氧化物歧化酶活性单位;在37℃恒温条件下,pH为6.2时,在波长 530 nm处,每分钟引起吸光度下降0.001为1个溶菌酶活性单位。
1.4 数据处理和统计分析
结果以平均值±标准差(mean±SD)表示。数据采用SPSS 19.0进行双因素方差分析(two-way ANOVA),之后采用Duncan氏法进行多重比较,显著性水平为P<0.05。以增重率为指标进行折线模型分析。
2 结 果
2.1 饲料蛋白质和脂肪水平对亚东鲑亲鱼生长性能和性腺指数的影响
由表2可知,饲料蛋白质和脂肪水平对亚东鲑亲鱼的成活率未产生显著影响(P>0.05)。饲料脂肪水平为9%时,增重率随饲料蛋白质水平的升高而升高;而饲料系数则随饲料蛋白质水平的升高先降低,至蛋白质水平为48%时又有所升高。饲料脂肪水平为18%,增重率随饲料蛋白质水平的升高先升高,至蛋白质水平为48%时又有所降低;而饲料系数则随饲料蛋白质水平的升高逐渐降低。饲料蛋白质水平为36%~42%时,18%脂肪水平组的增重率高于9%脂肪水平组,饲料系数与之相反。饲料蛋白质水平为45%~48%时,9%脂肪水平组的增重率高于18%脂肪水平组,饲料系数与之相反。饲料蛋白质和脂肪水平对亚东鲑雌性亲鱼的性腺指数未产生显著影响(P>0.05)。饲料脂肪水平为9%时,雄性亲鱼的性腺指数随饲料蛋白质水平的升高整体上呈下降趋势,仅在蛋白质水平为42%时有小幅回升;饲料脂肪水平为18%时,雄性亲鱼的性腺指数随饲料蛋白质水平的升高先降低,至蛋白质水平为48%时又有所回升。饲料蛋白质水平为36%~42%时,18%脂肪水平组雄性亲鱼的性腺指数高于9%脂肪水平组。饲料蛋白质水平为45%~48%时,9%脂肪水平组雄性亲鱼的性腺指数高于18%脂肪水平组。饲料蛋白质和脂肪水平对增重率、饲料系数和成活率均无显著的交互作用(P>0.05),对雄性亲鱼的性腺指数具有显著的交互作用(P<0.05)。以增重率为指标,进行折线模型回归分析可知:饲料脂肪水平为9%时,其蛋白质需要量为43.11%(Y=2.970 03X-75.683,X≤43.11;Y=-0.512 3X-66.835 7,X≥43.11;R2=0.686 2);饲料脂肪水平为18%时,其蛋白质需要量为45.69%(Y=2.746 9X-78.634 7,X≤45.69;Y=-0.526 9X+71.830 3,X≥45.69;R2=0.778 7)。
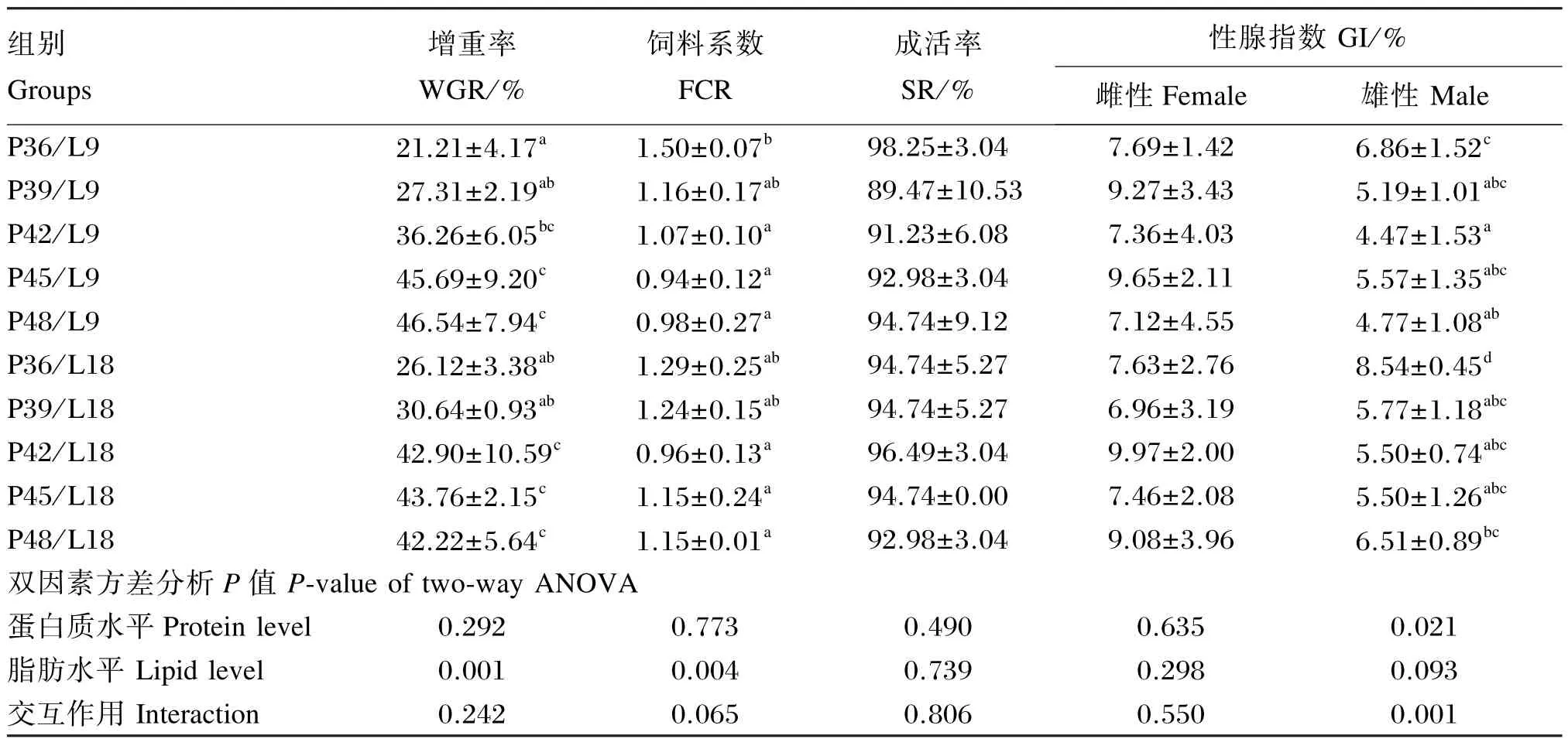
表2 饲料脂肪和蛋白质水平对亚东鲑亲鱼生长性能和性腺指数的影响Table 2 Effects of dietary protein and lipid levels on growth performance and GI ofSalmo trutta fariobroodstock
2.2 饲料蛋白质和脂肪水平对亚东鲑亲鱼消化酶活性的影响
由表3可知,饲料脂肪水平为9%或18%时,随饲料蛋白质水平的升高,胃、肠道和幽门盲囊的脂肪酶活性均呈增加趋势,P48/L18组显著高于P36/L9、P36/L18组(P<0.05);而胃、肠道和幽门盲囊的蛋白酶活性虽有所增加,但各组间无显著差异(P>0.05)。同一蛋白质水平下,18%脂肪水平组胃、肠道和幽门盲囊的蛋白酶和脂肪酶活性均略高于9%脂肪水平组。饲料蛋白质和脂肪水平对亚东鲑亲鱼的胃、肠道和幽门盲囊淀粉酶活性未产生显著影响(P>0.05)。饲料蛋白质和脂肪水平对消化道不同部位各消化酶活性均无显著的交互作用(P>0.05)。
2.3 饲料蛋白质和脂肪水平对亚东鲑亲鱼血清生化指标的影响
由表4可知,饲料脂肪水平为9%时,随饲料蛋白质水平的升高,血清氨含量逐渐增加,在蛋白质水平为48%时有最大值,显著高于蛋白质水平为36%时(P<0.05);饲料脂肪水平为18%时,随饲料蛋白质水平的升高,血清氨含量逐渐增加,蛋白质水平为48%时有最大值,显著高于蛋白质水平为36%和39%时(P<0.05)。饲料脂肪水平为9%时,血清球蛋白含量在蛋白质水平为48%时达最小值,显著低于蛋白质水平为 45%时(P<0.05),其余各组间差异不显著(P>0.05);饲料脂肪水平为18%时,血清球蛋白含量在蛋白质水平为39%时达最小值,显著低于蛋白质水平为42%时(P<0.05),其余各组间差异不显著(P>0.05)。饲料蛋白质和脂肪水平对亚东鲑亲鱼血清葡萄糖、甘油三酯、总蛋白和白蛋白含量未产生显著影响(P>0.05)。饲料蛋白质和脂肪水平对血清氨、葡萄糖、甘油三酯、总蛋白、白蛋白和球蛋白含量均无显著的交互作用(P>0.05)。
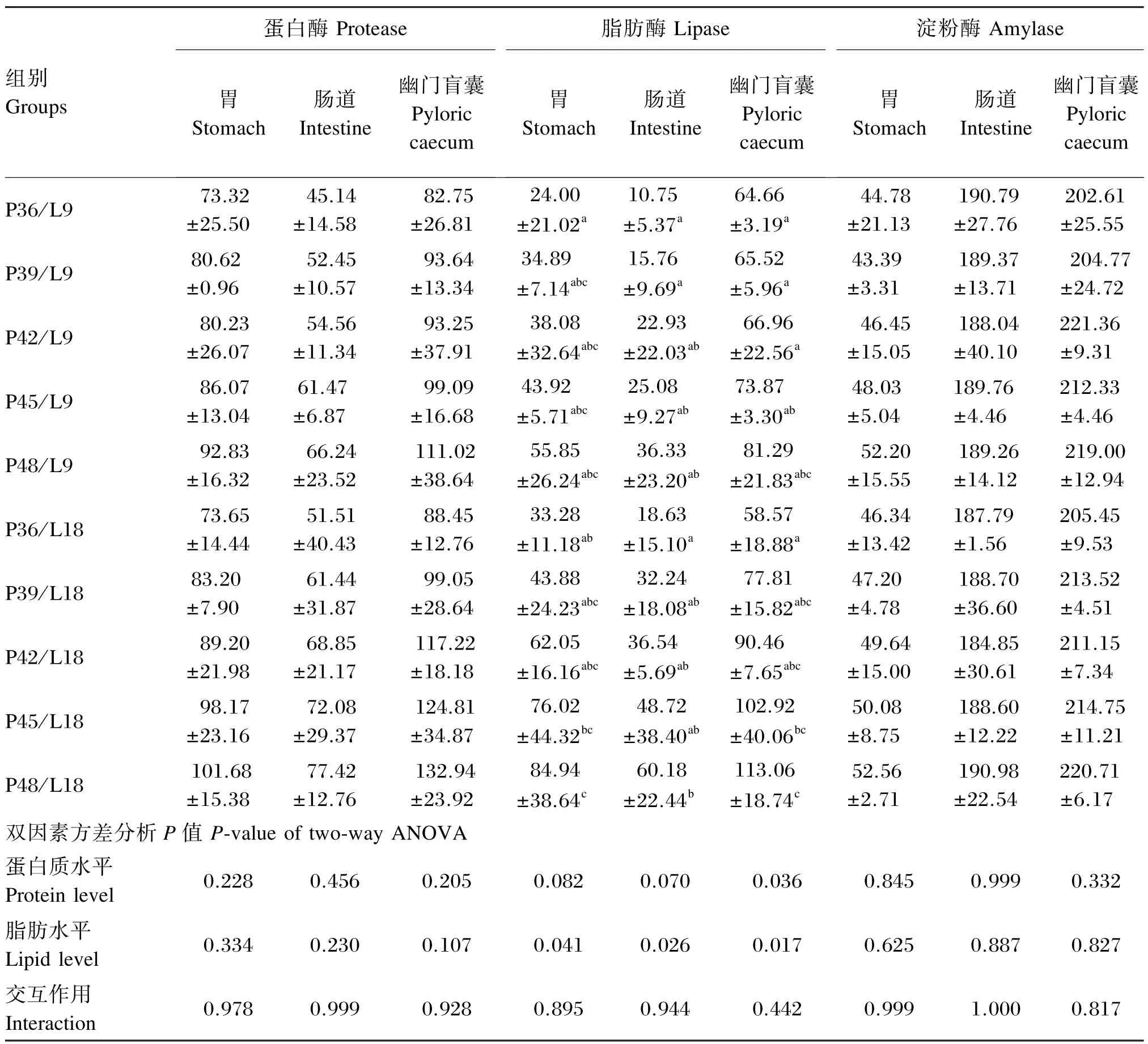
表3 饲料蛋白质和脂肪水平对亚东鲑亲鱼消化酶活性的影响Table 3 Effects of dietary protein and lipid levels on digestive enzyme activities ofSalmo trutta fariobroodstock U/g prot
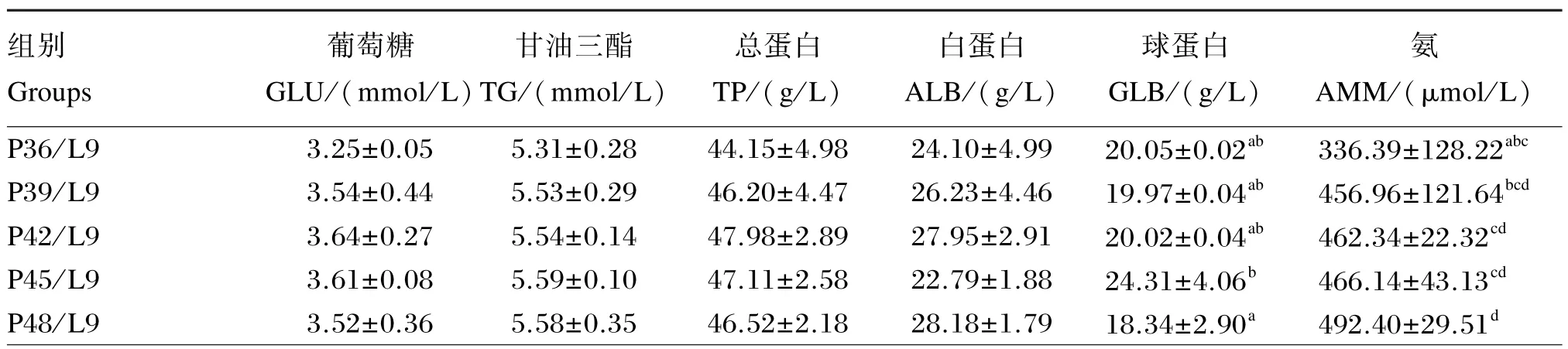
表4 饲料脂肪和蛋白质水平对亚东鲑亲鱼血清生化指标的影响Table 4 Effects of dietary protein and lipid levels on serum biochemical indices ofSalmo trutta fariobroodstock
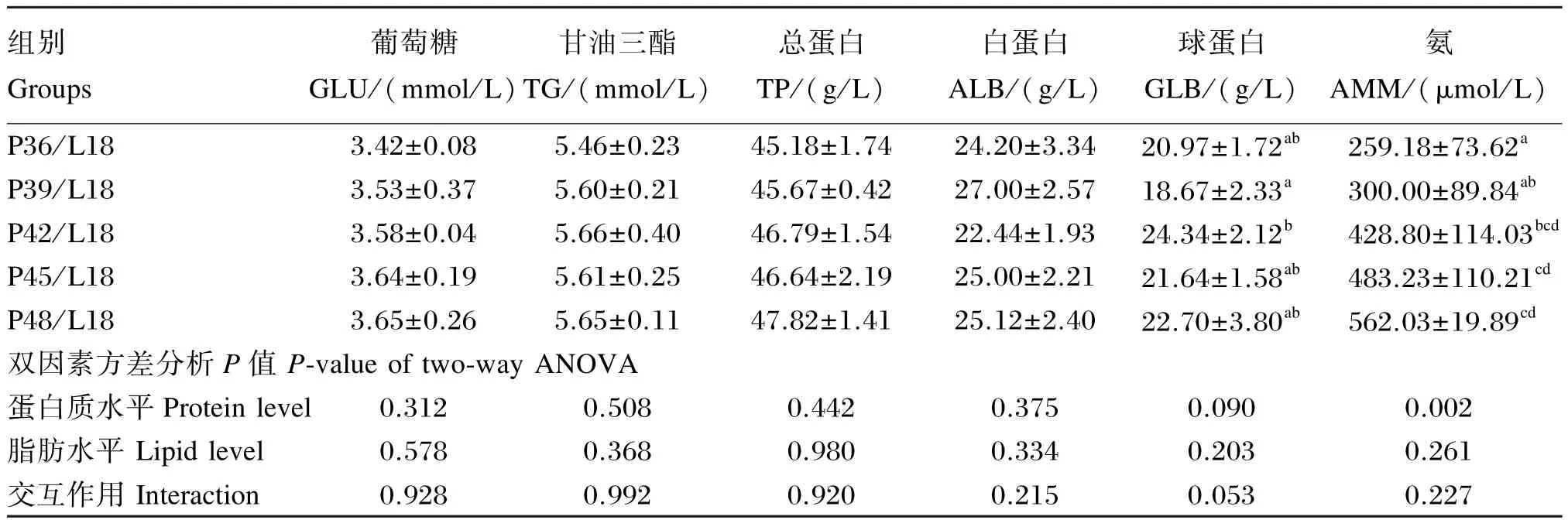
续表4
2.4 饲料蛋白质和脂肪水平对亚东鲑亲鱼血清非特异性免疫和抗氧化指标的影响
由表5可知,饲料蛋白质和脂肪水平对血清超氧化物歧化酶、溶菌酶活性和丙二醛含量的影响均不显著(P>0.05),但以低蛋白质(36%)和低脂肪水平(9%)组(P36/L9组)的血清超氧化物歧化酶和溶菌酶活性最低,丙二醛含量最高。饲料蛋白质和脂肪水平对血清超氧化物歧化酶、溶菌酶活性和丙二醛含量均无显著的交互作用(P>0.05)。
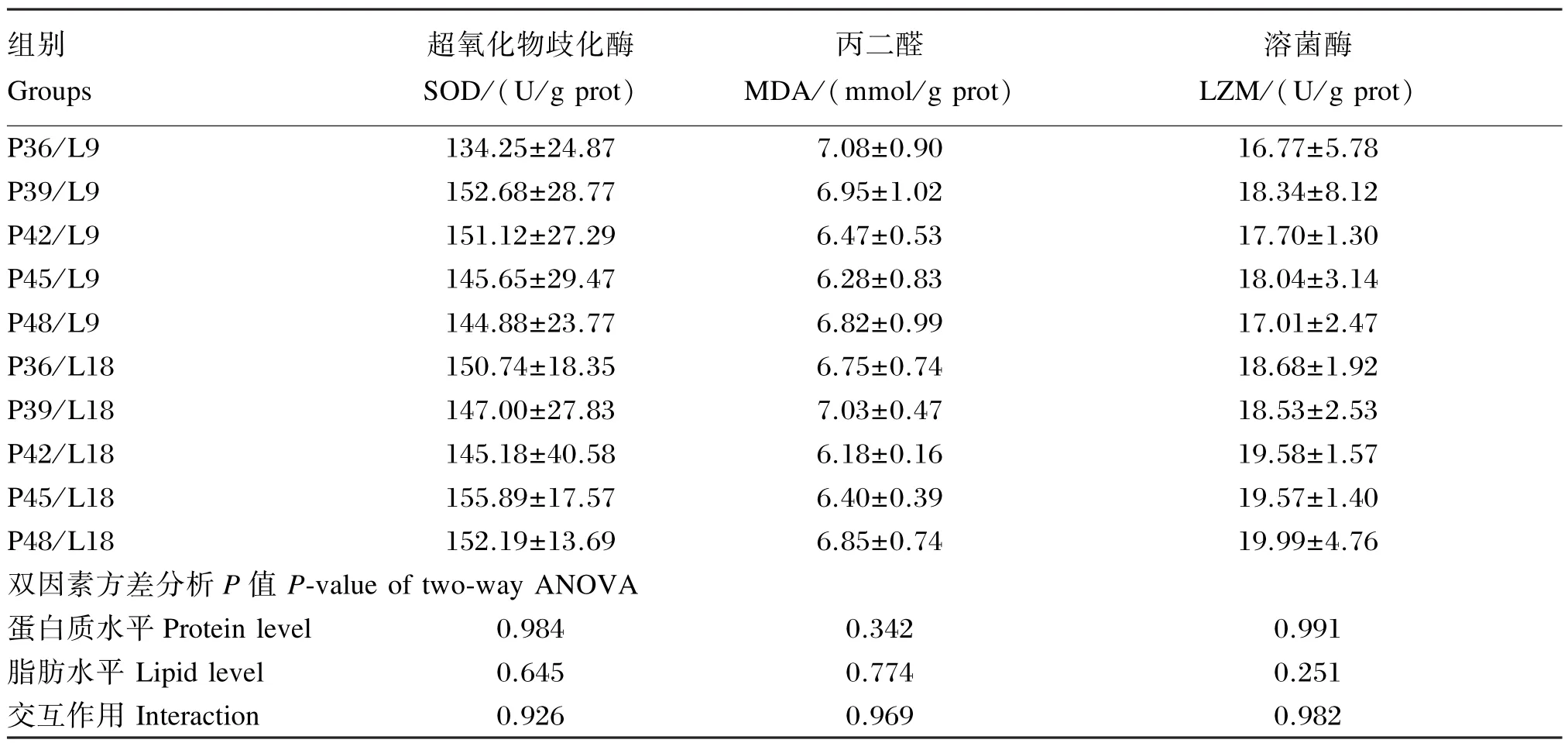
表5 饲料脂肪和蛋白质水平对亚东鲑亲鱼血清非特异性免疫和抗氧化指标的影响Table 5 Effects of dietary protein and lipid levels on serum nonspecific immune and antioxidant indices ofSalmo trutta fariobroodstock
3 讨 论
3.1 饲料蛋白质和脂肪水平对亚东鲑亲鱼生长性能和性腺指数的影响
本研究表明,饲料蛋白质水平可从48%降至42%而不影响亚东鲑亲鱼的生长性能和性腺指数。饲料中的蛋白质被鱼类摄入后消化分解成氨基酸,然后作为蛋白质合成的底物或者部分用于能源消耗。从增重率的结果来看,36%蛋白质水平组的生长性能明显低于45%~48%蛋白质水平组,结果类似于石斑鱼[11]等,这主要由于饲料蛋白质水平较低时,为鱼体供应的氨基酸不足。从增重率来看,饲料脂肪水平由9%升至18%时,增重率显著增加,因此,9%的脂肪水平尚不能满足亚东鲑亲鱼生长的需要。另外,本研究结果表明,低蛋白质水平下的饲料系数较高,这可能由于低蛋白质水平饲料缺乏氨基酸,特别是必需氨基酸,其导致饲料利用率降低。性腺必须与其相应的生态条件紧密地联系才能发育完善,当饲料蛋白质和脂肪水平不适宜时,鱼体代谢会发生紊乱[12]。本试验条件下,饲料蛋白质和脂肪水平对雌性亲鱼性腺指数无显著影响,而雄性亲鱼性腺指数随蛋白质水平的升高而降低,该结果与在花鲢[13]和绿色海胆[14]上的结果类似。 因此,仅以性腺指数来看,32%的蛋白质和9%的脂肪水平可能已能满足亚东鲑亲鱼性腺发育的需求。
3.2 饲料蛋白质和脂肪水平对亚东鲑亲鱼消化酶活性的影响
饲料成分对鱼类消化酶活性的影响具有复杂性和多样性[15]。研究表明,在一定范围内,饲料蛋白质水平与瓦氏黄颡鱼[16]、斑点叉尾[17]、圆斑星鲽[18]等鱼体的蛋白酶活性呈正相关。本试验结果显示,饲料脂肪水平为9%或18%时,随饲料蛋白质水平的升高,亚东鲑亲鱼胃、肠道和幽门盲囊的蛋白酶活性也有所增加。饲料高蛋白质水平可诱导鱼类消化道蛋白酶分泌且提高其活性,增强其对蛋白质的消化能力,从而促进鱼体蛋白质的合成[19]。然而,亚东鲑亲鱼消化道各部位蛋白酶活性未出现显著性差异,结果与鲶[20]类似,这可能由于亚东鲑亲鱼的规格较大,机体分泌的蛋白酶可以满足不同蛋白质水平(36%~48%)饲料的需要。 在一定范围内,圆斑星鲽[18]、瓦氏黄颡鱼[16]和团头鲂[21]消化道脂肪酶活性随饲料蛋白质水平的升高而降低,而印度野鲮[22]、瓦氏黄颡鱼[16]等鱼类的脂肪酶活性不受饲料蛋白质水平的影响,其主要与试验饲料中的脂肪水平一致有关。本结果显示,亚东鲑亲鱼胃、肠道和幽门盲囊的脂肪酶活性随饲料蛋白质水平的升高而有所增加,说明其对蛋白质和脂肪的利用较好[23]。饲料蛋白质水平对鱼类淀粉酶活性的影响因鱼的种类而异。一些鱼类的淀粉酶活性随饲料蛋白质水平的升高而增加,例如虹鳟[24]。本试验中亚东鲑亲鱼消化道各部位淀粉酶的活性无显著变化,结果与海鲷[25]等类似,可能其淀粉酶活性与饲料蛋白质水平并无直接关系,主要由其自身的遗传规律和发育过程决定。
饲料脂肪水平对鱼体脂肪酶活性的影响存在种类、季节和生长阶段的变化。在本试验条件下,随着饲料脂肪水平的升高,亚东鲑亲鱼胃、肠道和幽门盲囊的脂肪酶活性逐渐升高,该结果与在褐鲳鲉[26]、牙汉鱼[27]、点篮子鱼[28]、翘嘴红鲌[29]、舌齿鲈[30]等上所得结果类似,表明鱼体可通过提高体内脂肪酶活性来增强对高脂肪水平饲料的消化吸收。脂肪酶是诱导酶[31],且受饲料脂肪水平影响[32],这可能是亚东鲑亲鱼消化道各部位脂肪酶活性与饲料脂肪水平呈正相关关系的主要原因。然而,吉富罗非鱼[33]、闪光鸭嘴鲶[34]、军曹鱼[35]等的脂肪酶活性不受饲料脂肪水平的影响,真鲷[36]等的脂肪酶的活性随饲料脂肪水平的升高而降低。另外,本结果显示,即使饲料蛋白质水平(48%)较高时,亚东鲑亲鱼消化道各部位的脂肪酶活性仍表现为高(18%)脂肪水平组高于低(9%)脂肪水平组。可见,高脂肪水平饲料并未抑制亚东鲑亲鱼对蛋白质的利用(有研究认为过量的脂肪可抑制氨基酸吸收和代谢,降低蛋白质效率[37-38])。饲料脂肪水平对蛋白酶和淀粉酶活性也有一定影响。低脂肪水平可促进异育银鲫[39]、胭脂鱼[40]、七彩神仙鱼[41]肠道蛋白酶和淀粉酶的分泌,其原因可能是脂肪供给不足时,促使鱼体增强对蛋白质和碳水化合物的消化吸收能力以维持其正常的生命活动。对于有些鱼类来说,饲料脂肪水平在适宜范围内,其消化道蛋白酶和淀粉酶活性较高,当脂肪水平高于或低于最适范围时,消化道蛋白酶和淀粉酶活性降低[29,42]。适宜脂肪水平使消化道蛋白酶和淀粉酶活性保持最强,以提高其对饲料蛋白质和碳水化合物的消化和吸收,而过量脂肪的添加会抑制消化道中蛋白酶和淀粉酶的活性。本试验中亚东鲑亲鱼消化道各部位蛋白酶与淀粉酶活性在各组间无显著差异,可能是因为蛋白酶与淀粉酶活性在9%~18%脂肪水平范围内时不受脂肪水平的影响或受影响的程度不大。
3.3 饲料蛋白质和脂肪水平对亚东鲑亲鱼血清生化指标的影响
本试验中,饲料蛋白质和脂肪水平对血清葡萄糖含量没有产生显著影响,结果类似于在吉富罗非鱼[43]和大西洋白姑鱼[44]上所得结果,说明亚东鲑亲鱼的糖代谢没有受到饲料蛋白质和脂肪水平的影响,这可能是亲鱼体内糖原贮备充足,未出现能量供应不足而导致的血糖水平降低的现象[45]。另外,饲料蛋白质和脂肪水平对亚东鲑亲鱼血清总蛋白含量亦无显著影响,说明其可能通过自身调解蛋白质稳态以适应繁殖性能的需要。甘油三酯是血脂的重要组成部分,其含量在一定程度上反映了肝脏对脂肪的代谢状况[46]。本试验结果显示,血清甘油三酯的含量随饲料脂肪水平的升高有升高的趋势,结果与鳗鲡[32]、团头鲂[47]和草鱼[48]等所得结果相似。这表明饲料中较多的脂肪除了用于生长、繁殖等生理活动外,其余会转变成为脂类,储存在亚东鲑亲鱼体内,当饲料供给不足时,这一部分脂类会转化为能量,促进亲鱼生殖系统的脂肪合成。鱼体肝脏脂肪积累到一定程度时,血液中甘油三酯含量会减少,这在白甲鱼[49]、吉富罗非鱼[43]等的研究中均有报道,这可能是由于饲料中过高脂肪水平使肝脏出现营养诱导性脂肪蓄积,使肝细胞受到损伤,合成甘油三酯的能力减弱。
血清氨含量反映鱼体的蛋白质分解状况。饲料中的蛋白质经分解后产生氨基酸,通过吸收进入鱼体内的氨基酸代谢库,一部分氨基酸被氧化分解产生氨和尿素等,其中氨是鱼类氨基酸分解代谢的主要产物,产生的氨在血液中累积最终由鳃排出体外[50]。亚东鲑亲鱼血清氨含量随着饲料蛋白质水平的升高而上升,在大黄鱼[50]、克琳雷氏鲶[51]、银鲈[52]等研究中也得到类似的结果。 低蛋白质水平条件下,亚东鲑亲鱼的血清氨含量明显处于较低水平,此时蛋白质分解代谢较弱,主要用于蛋白质沉积和鱼体生长。当蛋白质过量时(48%蛋白质水平),蛋白质合成代谢下降,分解代谢增强,从而抑制亚东鲑亲鱼的蛋白质沉积。
3.4 饲料蛋白质和脂肪水平对亚东鲑亲鱼血清非特异性免疫和抗氧化指标的影响
饲料蛋白质和脂肪水平均会影响鱼体的免疫机能。研究发现,随着饲料蛋白质水平的升高,虹鳟的血清溶菌酶活性显著提高[53];饲料脂肪水平的升高会降低欧洲鲈吞噬细胞的呼吸爆发活性[53],但可增强点带石斑鱼血清溶菌酶活性和白细胞呼吸爆发活性[54];用嗜水气单胞菌对拉氏攻毒48 h后,低、中蛋白质组(23%和28%蛋白质组)的存活率高于高蛋白质组(33%和38%蛋白质组),低脂肪组(5%脂肪组)的存活率高于高、中脂肪组(11%和8%脂肪组)[55]。因此,饲料中适宜的蛋白质和脂肪水平对维持鱼体的免疫机能具有重要意义。然而,不同鱼类对饲料蛋白质和脂肪水平的敏感性有所不同。当饲料中脂肪水平超过10%时,草鱼的肝脏受到损伤[56]。亚东鲑为肉食性鱼类,本试验结果显示,饲料脂肪水平为18%时,血清溶菌酶活性未明显高于饲料脂肪水平为9%时,说明18%脂肪水平可能未对机体的非特异免疫力造成损伤。 对鲤[57]、异育银鲫[58]、奥尼罗非鱼[59]的研究表明,饲料蛋白质水平对血清溶菌酶活性无显著影响。本结果与之类似,饲料蛋白质水平在33%~48%范围内,亚东鲑亲鱼血清溶菌酶活性无显著变化,这可能与其生长阶段有关。以往的研究主要集中在仔稚鱼阶段,而本试验采用的亚东鲑亲鱼可能对试验条件下的蛋白质和脂肪水平的敏感性不强。此外,本试验由于加工工艺的原因,未进行较高脂肪水平的添加,因此后续研究有待进一步证实高脂肪水平对亚东鲑亲鱼免疫力的影响。
超氧化物歧化酶和过氧化氢酶主要清除超氧自由基,歧化、催化过氧化氢和活性氧。活性氧会影响内源性酶活性,或膜多不饱和脂肪酸,导致脂质过氧化和丙二醛的积累[60]。饲料的蛋白质和脂肪水平显著影响鱼体的抗氧化能力。在一定的范围内,饲料蛋白质水平降低时,可提高鲤血清与肝胰腺超氧化物歧化酶和过氧化氢酶的活性,当蛋白质水平低于26%时,丙二醛含量显著升高[57]。随着饲料蛋白质水平的升高,异育银鲫血清超氧化物歧化酶活性呈先上升后下降的趋势[58],锦鲤血清超氧化物歧化酶和过氧化氢酶活性呈上升趋势[61],而金头鲷超氧化物歧化酶和过氧化氢酶活性呈下降趋势[62]。随着饲料脂肪水平的升高,大西洋鲑血清、肝脏的超氧化物歧化酶活性增强,血清丙二醛含量降低[19]。本结果显示,饲料蛋白质和脂肪水平不影响亚东鲑亲鱼的血清超氧化物歧化酶活性和丙二醛含量,说明对机体的抗氧化力无显著影响。然而,高脂肪水平(>18%)对亚东鲑亲鱼是否会抑制鱼体代谢且加快机体的过氧化尚需进一步研究。
4 结 论
① 在一定范围内,适当提高饲料蛋白质和脂肪水平,能够提高亚东鲑亲鱼消化道内脂肪酶的活性,但是蛋白酶和淀粉酶活性没有显著变化。
②饲料蛋白质和脂肪水平分别为36%~48%和9%~18%时,亚东鲑亲鱼除血清氨和球蛋白含量有显著变化外,其他血清生化指标、非特异性免疫和抗氧化指标均无显著变化。
③ 以增重率为指标,经折线模型分析得出脂肪水平为9%和18%时,亚东鲑亲鱼对蛋白质需要量分别为43.11%和45.69%。
[1]IZQUIERDO M S,FERNÁNDEZ-PALACIOS H,TACON A G J.Effect of broodstock nutrition on reproductive performance of fish[J].Aquaculture,2001,197(1/2/3/4):25-42.
[2]常青,梁萌青,薛华,等.亲鱼营养的研究进展[J].海洋水产研究,2002,23(2):65-71.
[3]张春霖,王文滨.西藏鱼类初篇[J].动物学报,1962,14(4):529-536.
[4]VØLLESTAD L A,OLSEN E M,FORSETH T.Growth-rate variation in brown trout in small neighbouring streams:evidence for density-dependence?[J].Journal of Fish Biology,2002,61(6):1513-1527.
[5]ASBJØRN VØLLESTAD L,MOLAND OLSEN E.Non-additive effects of density-dependent and densityindependent factors on brown trout vital rates[J].Oikos,2008,117(11):1752-1760.
[6]蒲德永,王志坚,赵海鹏,等.亚东鲑消化系统的形态学和组织学观察[J].四川动物,2006,25(4):825-828.
[7]豪富华,陈毅峰,蔡斌.西藏亚东鲑的胚胎发育[J].水产学报,2006,30(3):289-296.
[8]豪富华,陈毅峰,唐卫星,等.亚东鲑的年龄与生长的研究[J].水生生物学报,2007,31(5):731-737.
[9]孟玮,杨天燕,海萨,等.基于线粒体COI基因序列的亚东鲑DNA条形码研究[J].水产学杂志,2010,23(1):6-10.
[10]桂远明.水产动物机能学实验[M].北京:中国农业出版社,2004:92-98.
[11]JIANG S T,WU X Y,LI W F,et al.Effects of dietary protein and lipid levels on growth,feed utilization,body and plasma biochemical compositions of hybrid grouper(Epinephelus lanceolatus×Epinephelus fuscoguttatus♀)juveniles[J].Aquaculture,2015,446(1):148-155.
[12]ZHANG Y L,SONG L,LIU R P,et al.Effects of dietary protein and lipid levels on growth,body composition and flesh quality of juvenile topmouth culter,Culter alburnusBasilewsky[J].Aquaculture Research,2016,47(8):2633-2641.
[13]SANTIAGO C B,CAMACHO A S,LARON M A.Growth and reproductive performance of bighead carp(Aristichthys nobilis)reared with or without feeding in floating cages[J].Aquaculture,1991,96(2):109-117.
[14]DAGGETT T L,PEARCE C M,TINGLEY M,et al.Effect of prepared and macroalgal diets and seed stock source on somatic growth of juvenile green sea urchins(Strongylocentrotus droebachiensis)[J].Aquaculture,2005,244(1/2/3/4):263-281.
[15]黎军胜,李建林,吴婷婷.饲料成分与环境温度对奥尼罗非鱼消化酶活性的影响[J].中国水产科学,2004,11(6):585-588.
[16]孙翰昌,徐敬明,庞敏.饲料蛋白水平对瓦氏黄颡鱼消化酶活性的影响[J].水生态学杂志,2010,3(2):84-88.
[17]蒋广震,刘文斌,王煜衡,等.饲料中蛋白脂肪比对斑点叉尾幼鱼生长、消化酶活性及肌肉成分的影响[J].水产学报,2010,34(7):1129-1135.
[18]吕云云,陈四清,于朝磊,等.饲粮蛋白脂肪比对圆斑星鲽(Verasper variegates)生长、消化酶及血清生化指标的影响[J].渔业科学进展,2015,36(2):118-124.
[19]柳阳,李勇,高婷婷,等.脂肪和蛋白质营养对封闭循环水养殖大西洋鲑生长和肌肉品质的效应[J].海洋科学,2013,37(6):47-59.
[20]LAZZARI R,RADÜNZ NETO J,PEDRON F D A,et al.Protein sources and digestive enzyme activities in jundiá(Rhamdia quelen)[J].Scientia Agricola,2010,67(3):259-266.
[21]HABTE-TSION H M,LIU B,GE X P,et al.Effects of dietary protein level on growth performance,muscle composition,blood composition,and digestive enzyme activityofWuchangbream (Megalobramaamblycephala)fry[J].The Israeli Journal of Aquaculture,2013,65(3):925-933.
[22]DEBNATH D,PAL A K,SAHU N P,et al.Digestive enzymes and metabolic profile ofLabeo rohitafingerlings fed diets with different crude protein levels[J].Comparative Biochemistry and Physiology Part B:Biochemistry and Molecular Biology,2007,146(1):107-114.
[23]DE ALMEIDA L C,LUNDSTEDT L M,MORAES G.Digestive enzyme responses of tambaqui(Colosso-ma macropomum)fed on different levels of protein and lipid[J].Aquaculture Nutrition,2006,12(6):443-450.
[24]KAWAI S I,IKEDA S.Studies on digestive enzymes of fishes.3.Development of the digestive enzymes of rainbow-trout after hatching and the effect of dietary change on the activities of digestive enzymes in the juvenile stage[J].Bulletin of the Japanese Society of Scientific Fisheries,1973,39(7):819-823.
[25]MOYANO F J,DÍAZ M,ALARCÓN F J,et al.Characterization of digestive enzyme activity during larval development of gilthead seabream (Sparus aurata)[J].Fish Physiology and Biochemistry,1996,15(2):121-130.
[26]岳彦峰,彭士明,施兆鸿,等.饲料脂肪水平对褐菖鲉生长,肠道消化酶及主要脂代谢酶活力的影响[J].南方水产科学,2012,8(6):50-56.
[27]GÓMEZ-REQUENIP,BEDOLLA-CÁZARES F,MONTECCHIA C,et al.Effects of increasing the dietary lipid levels on the growth performance,body composition and digestive enzyme activities of the teleost pejerrey(Odontesthes bonariensis)[J].Aquaculture,2013,416-417:15-22.
[28]章龙珍,朱卫,王妤,等.饲料脂肪水平对点篮子鱼消化酶活性和血液主要生化指标的影响[J].海洋渔业,2014,36(2):170-176.
[29]向枭,周兴华,陈建,等.饲料中脂肪含量对翘嘴红鲌幼鱼消化酶活性的影响[J].北京水产,2008(5):35-38.
[30]INFANTE J L Z,CAHU C L.High dietary lipid levels enhance digestive tract maturation and improve Dicentrarchus labrax larval development[J].The Journal of Nutrition,1999,129(6):1195-1200.
[31]ALIYU-PAIKO M,HASHIM R,SHU-CHIEN A C.Influence of dietary lipid/protein ratio on survival,growth,body indices and digestive lipase activity in Snakehead(Channa striatus,Bloch 1793)fry reared in re-circulating water system[J].Aquaculture Nutrition,2010,16(5):466-474.
[32]MA X,HU Y,WANG X Q,et al.Effects of practical dietary protein to lipid levels on growth,digestive enzyme activities and body composition of juvenile rice field eel(Monopterus albus)[J].Aquaculture International,2014,22(2):749-760.
[33]韩光明,王爱民,徐跑,等.饲料中脂肪水平对吉富罗非鱼幼鱼成活率、肌肉成分及消化酶活性的影响[J].上海海洋大学学报,2010,19(4):469-474.
[34]LEE H M,CHO K C,LEE J E,et al.Dietary protein requirement of juvenile giant croaker,Nibea japonicaTemminck& Schlegel[J].Aquaculture Research,2001,32(Suppl.1):112-118.
[35]汤保贵,陈刚,张健东,等.饵料系列对军曹鱼仔鱼生长、消化酶活力和体成分的影响[J].水生生物学报,2007,31(4):479-484.
[36]王重刚,陈品健,顾勇,等.不同饵料对真鲷稚鱼消化酶活性的影响[J].海洋学报,1998,20(4):103-106.
[37]MORAIS S,CONCEIÇÃO L E C,RØNNESTAD I,et al.Dietary neutral lipid level and source in marine fish larvae:effects on digestive physiology and food intake[J].Aquaculture,2007,268(1/2/3/4):106-122.
[38]MOHANTA K N,MOHANTY S N,JENA J K,et al.Optimal dietary lipid level of silver barb,Puntius gonionotusfingerlings in relation to growth,nutrient retention and digestibility,muscle nucleic acid content and digestive enzyme activity[J].Aquaculture Nutrition,2008,14(4):350-359.
[39]王爱民.饲料脂肪水平对吉富罗非鱼生长及脂肪代谢调节的研究[D].博士学位论文,南京:南京农业大学,2011.
[40]王朝明.岩原鲤幼鱼蛋白需求和胭脂鱼幼鱼脂肪需求研究[D].硕士学位论文.重庆:西南大学,2011.
[41]王吉桥,耿加振,姜玉声,等.饲料中脂肪含量对七彩神仙鱼幼鱼生长和消化的影响[J].水产学杂志,2009,22(1):24-30.
[42]FOUNTOULAKI E,ALEXIS M N,NENGAS I,et al.Effect of diet composition on nutrient digestibility and digestive enzyme levels of gilthead sea bream(Sparus aurataL.)[J].Aquaculture Research,2005,36(13):1243-1251.
[43]石桂城.脂肪对低温下吉富罗非鱼生化指标及脂肪酸组成的影响[D].硕士学位论文.湛江:广东海洋大学,2012.
[44]CHATZIFOTIS S,PANAGIOTIDOU M,PAPAIOANNOU N,et al.Effect of dietary lipid levels on growth,feed utilization,body composition and serum metabolites of meagre(Argyrosomus regius)juveniles[J].Aquaculture,2010,307(1/2):65-70.
[45]ZHOU M,WANG A L,XIAN J A.Variation of free amino acid and carbohydrate concentrations in white shrimp,Litopenaeus vannamei:effects of continuous cold stress[J].Aquaculture,2011,317(1/2/3/4):182-186.
[46]程汉良,夏德全,吴婷婷.鱼类脂类代谢调控与脂肪肝[J].动物营养学报,2006,18(4):294-298.
[47]LI X F,JIANG Y Y,LIU W B,et al.Protein-sparing effect of dietary lipid in practical diets for blunt snout bream (Megalobrama amblycephala) fingerlings:effects on digestive and metabolic responses[J].Fish Physiology and Biochemistry,2012,38(2):529-541.
[48]高攀,蒋明,文华,等.不同蛋白能量比饲料对草鱼幼鱼消化酶活性的影响[J].淡水渔业,2009,39(6):54-58.
[49]向枭,周兴华,陈建,等.饲料脂肪水平对白甲鱼幼鱼生长性能、体组成和血清生化指标的影响[J].动物营养学报,2013,25(8):1805-1816.
[50]林淑琴,成琳,张文兵,等.饲料蛋白质水平对大黄鱼体内蛋白质的沉积和代谢反应的影响[J].中国海洋大学学报,2014,44(5):32-39.
[51]MELO J F B,LUNDSTEDT L M,METÓN I,et al.Effects of dietary levels of protein on nitrogenous metabolism ofRhamdia quelen(Teleostei:Pimelodidae)[J].Comparative Biochemistry and Physiology Part A:Molecular& Integrative Physiology,2006,145(2):181-187.
[52]YANG S D,LIOU C H,LIU F G.Effects of dietary protein level on growth performance,carcass composition and ammonia excretion in juvenile silver perch(Bidyanus bidyanus)[J].Aquaculture,2002,213(1/2/3/4):363-372.
[53]KIRON V,WATANABE T,FUKUDA H,et al.Protein nutrition and defence mechanisms in rainbow troutOncorhynchus mykiss[J].Comparative Biochemistry and Physiology Part A:Physiology,1995,111(3):351-359.
[54]LIN Y H,SHIAU S Y.Dietary lipid requirement of grouper,Epinephelus malabaricus,and effects on immune responses[J].Aquaculture,2003,225(1/2/3/4):243-250.
[55]蒋湘辉,骆小年,金广海,等.不同蛋白质和脂肪水平饲料对拉氏幼鱼生长、体成分和免疫力的影响[J].动物营养学报,2016,28(3):864-871.
[56]JIN Y,TIAN L X,ZENG S L,et al.Dietary lipid requirement on non-specific immune responses in juvenile grass carp(Ctenopharyngodon idella)[J].Fish&Shellfish Immunology,2013,34(5):1202-1208.
[57]鹿璇,程镇燕,孙金辉,等.降低饲料中蛋白质水平对鲤鱼生长、肉质及免疫力的影响[J].饲料工业,2014,35(6):29-35.
[58]蔡春芳,吴康,潘新法,等.蛋白质营养对异育银鲫生长和免疫力的影响[J].水生生物学报,2001,25(6):590-596.
[59]乐贻荣,杨弘,徐起群,等.饲料蛋白水平对奥尼罗非鱼(Oreochromis niloticus×O.aureus)生长、免疫功能以及抗病力的影响[J].海洋与湖沼,2013,44(2):493-498.
[60]GÉRET F,JOUAN A,TURPIN V,et al.Influence of metal exposure on metallothionein synthesis and lipid peroxidation in two bivalve mollusks:the oyster(Crassostrea gigas)and the mussel(Mytilus edulis)[J].Aquatic Living Resources,2002,15(1):61-66.
[61]姜志强,石洪玥,崔培,等.不同蛋白水平的螺旋藻饲料对锦鲤体色,生长及免疫的影响[J].东北农业大学学报,2012,43(3):95-103.
[62]MOURENTE G,DÍAZ-SALVAGO E,BELL J G,et al.Increased activities of hepatic antioxidant defence enzymes in juvenile gilthead sea bream(Sparus aurataL.)fed dietary oxidised oil:attenuation by dietary vitamin E[J].Aquaculture,2002,214(1/2/3/4):343-361.
Effects of Dietary Protein and Lipid Levels on Growth Performance,Digestive Enzyme Activities and Serum Indices ofSalmo trutta farioBroodstock
WANG Chang’an HU Guo∗SUN Peng GU Wei WANG Bingqian∗XU Qiyou
(Heilongjiang River Fisheries Research Institute,Chinese Academy of Fishery Sciences,Harbin150070,China)
This study was conducted to estimate the effects of dietary protein and lipid levels on growth performance,gonad index,digestive enzyme activities and serum indices ofSalmo trutta fariobroodstock.Ten kinds of practical experimental diets were formulated to contain five protein levels(36%,39%,42%,45% and 48%)and two lipid levels(9%and 18%)using 5×2 two-factor randomized design,and each diet was randomly assigned 3 replicates of 19 bloodstocks.TheSalmo trutta fariowith the initial body weight of(462.53±45.40)g were cultured in the water flow-through system for 77 days.The results showed as follows:weight gain rate was increased with the increase of dietary protein level under the same lipid level,on the contrary,feed conversion ratio was gradually decreased,but was increased at the 48%dietary protein level.Dietary protein and lipid levels had no significant effect on the gonad index of female bloodstocks(P>0.05).The gonad index of male bloodstocks was decreased firstly with the increase of dietary protein level,and then was increased at the 48%dietary protein level.The lipase activity of stomach,intestine and pyloric caeca was gradually increased with the increase of dietary protein level under the same lipid level,and 48%dietary protein level was significantly higher than 36%dietary protein level(P<0.05),the protease activity of stomach,intestine and pyloric caeca was also increased,but no significant difference was observed among the groups(P>0.05).Under the same protein level,the protease and lipase activities of stomach,intestine and pyloric caeca in 18%lipid level groups were slightly higher than those in 9%lipid level groups.The serum ammonia content was gradually increased with the increase of dietary protein level under the same lipid level.Dietary protein and lipid levels had no significant effect on the activities of serum superoxide dismutase and lysozyme,and the contents of malonaldehyde,glucose,triglyceride,total protein,albumin(P>0.05).In conclusion,Salmo trutta fariobroodstock can respond to dietary protein(36%to 48%)and lipid(9%to 18%)levels.With the weight gain rate as an index,the dietary protein requirement forSalmo trutta fariobroodstock is 43.11%and 45.69% when the dietary lipid level is 9%and 18%base on the broken-line model analysis,respectively.[Chinese Journal of Animal Nutrition,2017,29(2):571-582]
Salmo trutta fariobroodstock;protein;lipid;growth;digestive enzymes;serum indices
S963
A
1006-267X(2017)02-0571-12
10.3969/j.issn.1006-267x.2017.02.025
(责任编辑 菅景颖)
2016-08-02
国家公益性(农业)科研专项经费(201403012);农业部948引进项目(2014-Z57);国家科技支撑计划(2012BAD25B10);黑龙省自然科学基金项目(QC2015041)
王常安(1981—),男,辽宁东港人,助理研究员,博士,从事水产动物营养与饲料学研究。E-mail:gordoncase@126.com
∗通信作者:户 国,副研究员,E-mail:huguo@126.com;王炳谦,研究员,硕士生导师,E-mail:wbqfish@163.com
∗Corresponding authors:HU Guo,associate professor,E-mail:huguo@126.com;WANG Bingqian,professor,E-mail:wbqfish@163.com
猜你喜欢
英语世界(2022年9期)2022-10-18
文史春秋(2022年4期)2022-06-16
保健医苑(2022年4期)2022-05-05
当代水产(2021年9期)2021-12-02
当代水产(2021年8期)2021-11-04
医学前沿(2021年18期)2021-04-14
肝博士(2021年1期)2021-03-29
小哥白尼(趣味科学)(2020年1期)2020-06-16
中学生理科应试(2017年2期)2017-04-01
