
热带假丝酵母ctfat1p基因在吸收脂肪族化合物中的功能研究
2017-04-07 12:53石莹杨晓慧马春玲王腾飞肖静王瑞明汪俊卿
中国酿造 2017年3期
石莹,杨晓慧,马春玲,王腾飞,肖静,王瑞明,汪俊卿*
热带假丝酵母ctfat1p基因在吸收脂肪族化合物中的功能研究
石莹,杨晓慧,马春玲,王腾飞,肖静,王瑞明,汪俊卿*
(齐鲁工业大学生物工程学院山东省微生物工程重点实验室,山东济南250353)
以热带假丝酵母(Candida tropicalis)1798中的ctfat1p基因为研究对象,利用重叠PCR将ctfat1p基因内约500 bp片段与G418抗性基因(kanr)相连接,经末端单酶切后电转化至C.tropicalis1798感受态细胞中,通过一次同源单交换,将抗性基因kanr插入至ctfat1p基因内部,实现目的基因的敲除,并通过发酵实验分析Ctfat1p在热带假丝酵母脂肪酸跨膜转运过程中的功能。结果表明,经过G418抗性筛选和基因组PCR鉴定,成功获得ctfat1p基因缺失菌株C.tropicalis1798Δctfat1p;分析发现ctfat1p基因敲除对C.tropicalis1798以油脂为底物培养12 h后重组菌OD600nm值仅为野生型菌株的47.9%,表明ctfat1p基因敲除后影响C.tropicalis1798对油脂吸收利用。通过基因敲除手段构建ctfat1p基因缺失菌株,可以减弱细胞对长链脂肪酸的摄取,验证了ctfat1p基因为热带假丝酵母油脂吸收和利用的关键基因。
ctfat1p基因;热带假丝酵母;基因敲除;同源单交换
热带假丝酵母(Candida tropicalis)能够利用烷烃和脂肪酸作为唯一碳源,合成长链二元酸[1-2]。长链二元酸是一种重要的化工原料和医药中间体,不能从自然界中直接获取,微生物发酵法具有成本低、安全性高以及绿色环保等优点,在国内外受到普遍重视。目前,利用热带假丝酵母发酵正构烷烃生产长链二元酸的发酵工业放大技术己成熟[3-4]。随着石油资源的枯竭,以烷烃为原料生产长链二元酸面临日益严重的成本压力和原料压力。因此,寻找可再生的、廉价的烷烃替代品作为原料,成为长碳链二元酸产业所面临的重大挑战。
前期通过大量实验发现,以油脂代替烷烃为底物进行发酵产二元酸是可行的,但与烷烃相比利用效率较低。RACHINSKI V V等[5-8]研究发现,热带假丝酵母吸收烷烃能够通过胞吞途径直接吸收,与之相比,热带假丝酵母在利用油脂作为底物进行发酵时,则需要将油脂水解成长链脂肪酸才能进行转运,而长链脂肪酸的性质与烷烃有很大的区别,因此推测热带假丝酵母对油脂的吸收利用途径与烷烃可能存在不同。目前,热带假丝酵母中长链脂肪酸摄取、构成过程的详尽机制还不明确,长链脂肪酸的吸收可能包括蛋白调控运输和被动扩散。由于脂肪酸是疏水性物质,不能直接进入组织细胞中被利用。它们必须与细胞中的特殊蛋白质一起组成一个亲水性的分子基团,才能进入组织细胞并在细胞中运输[9]。近年来,大量研究通过基因敲除或者基因过表达试验证明,除了扩散渗透的方式,还存在由多个蛋白介导的竞争性的脂肪酸转运体系。目前己经有众多的试验肯定了基因fat1p能显著地促进脂肪细胞吸收脂肪酸[10-11]。
在酿酒酵母(Saccharomyces cerevisiae)中,长链脂肪酸转运体系已被充分表征[12]。酿酒酵母中存在特殊的长链脂肪酸转运蛋白负责长链脂肪酸的跨膜转运,被命名为脂肪酸转运蛋白(Fat1p)。BLACK P N等[13-15]的研究显示,基因fat1p缺陷的酵母与野生型相比,胞内没有积累外源添加的长链脂肪酸,并且转运长链脂肪酸的能力也消失,实验证明Fat1p具有转运长链脂肪酸的功能。而热带假丝酵母中类似基因尚未被报道,以酿酒酵母fat1p基因为模板在美国国家生物技术信息中心(national center of biotechnology information,NCBI)数据库查询可知,热带假丝酵母基因组中存在相似基因序列,所编码的蛋白质与酿酒酵母中的Fat1p序列一致性为50%,推测其与长链脂肪酸转运有关。本研究利用聚合酶链反应(polymerase chain reaction,PCR)技术两步构建敲除片段,酶切后转化至热带假丝酵母中,通过一次同源单交换,在目的基因内部插入一段含有筛选标记基因的序列,实现热带假丝酵母的长链脂肪酸转运蛋白CtFat1p基因的快速敲除。本研究构建了ctfat1p基因缺失突变株,为进一步研究其在热带假丝酵母中的功能奠定了基础。
1 材料与方法
1.1 材料与试剂
1.1.1 菌株、质粒与引物
热带假丝酵母(Candida tropicalis)1798:中国工业微生物菌种保藏管理中心;含有G418抗性基因的质粒pPIC9k:宝生物工程(大连)有限公司;实验所涉及引物见表1所示。
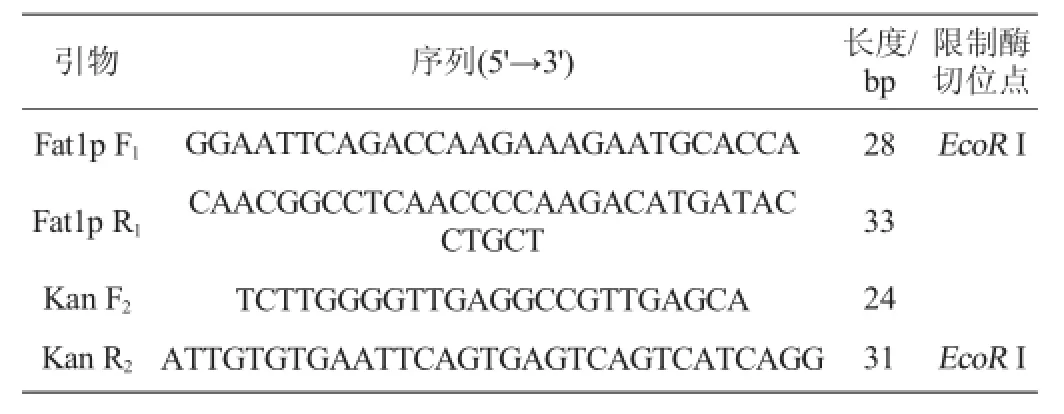
表1 实验所用引物Table 1 Primers used in this study
1.1.2 试剂
限制性内切酶:赛默飞世尔科技(中国)有限公司;G418、Ezup柱式基因组DNA抽提试剂盒、SanPrep柱式DNA胶回收试剂盒、高纯度质粒小量快速提取试剂盒:上海生工生物工程有限公司;其他试剂均为国产分析纯。
1.1.3 培养基
酵母浸出粉胨葡萄糖(yeast extract peptone dextrose,YEPD)培养基:葡萄糖20g/L、蛋白胨20g/L、酵母浸粉10g/L。
发酵培养基:(NH4)2SO41 g/L、酵母浸粉2 g/L、维生素B1(vitamin B1,VB1)0.2 g/L、NaCl 2 g/L、KH2PO48 g/L、Na2HPO4·12H2O10.08g/L、尿素3g/L、MgSO4·7H2O6.15g/L、葡萄糖(或十二烷、十六烷、油脂)62 g/L。
菌体增殖培养基:葡萄糖20 g/L、蛋白胨20 g/L、酵母浸粉10 g/L。
菌体复苏培养基:山梨醇1 mol/L。
以上培养基均需在高压蒸汽灭菌锅中115℃灭菌20min,葡萄糖单独灭菌;尿素和MgSO4·7H2O 110℃灭菌20 min,分开灭菌。
1.2 仪器与设备
ZQZY-CS恒温振荡培养箱:上海知楚仪器有限公司;ThermoblockPCR仪:德国Veriti公司;DYY-12型电泳仪:北京市六一仪器厂;MD2000H核酸超微量系列分光光度计:英国BioFuture公司;5804R型离心机、4308型电转仪:德国Eppendorf公司;7200可见分光光度计:尤尼柯上海仪器有限公司。
1.3 方法
1.3.1 重组同源性片段ctfat1p1-kanr的制备
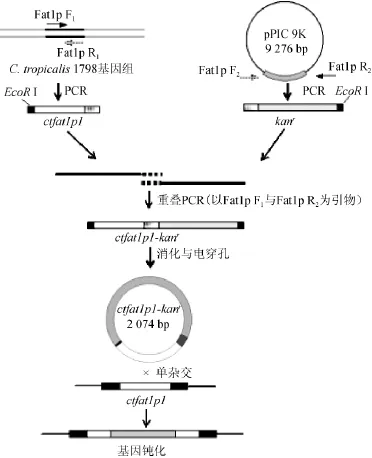
图1 C.tropicalis1798△ctfat1p构建流程图Fig.1 Flowchart of construction ofC.tropicalis1798△ctfat1p
使用上海生工Ezup柱式基因组DNA抽提试剂盒提取C.tropicalis1798菌体的DNA,并以此为模板,使用引物Fat1p F1和Fat1p R1进行PCR扩增,PCR扩增条件为:95℃预变性5 min;94℃变性30 s,57℃退火30 s,72℃延伸1.5 min,30个循环;72℃延伸10min,4℃保存,扩增获得长度为551bp用于ctfat1p基因敲除的同源臂ctfat1p1;使用高纯度质粒小量快速提取试剂盒提取质粒pPIC9k,以质粒pPIC9k为模板,使用引物Kan F2和Kan R2进行PCR扩增,PCR扩增条件为:95℃预变性5 min;94℃变性30 s,57℃退火30 s,72℃延伸3.5 min,30个循环;72℃延伸10 min,4℃保存,30个循环扩增获得长度为1 523 bp G418抗性基因片段kanr;使用SanPrep柱式DNA胶回收试剂盒(上海生工)胶回收制得的ctfat1p1片段与kanr片段为模板,Fat1p F1和Kan R2为引物进行PCR扩增进行重叠延伸PCR[16-18],PCR扩增条件为:(1)95℃预变性5 min;94℃变性30 s,57℃退火30 s,72℃延伸1.5 min,5个循环;72℃延伸2 min;(2)95℃预变性5 min,94℃变性30 s,55℃退火30 s,72℃延伸5 min,30个循环;72℃延伸10 min,4℃保存,扩增制得长度为长度2074bp的同源重组片段ctfat1p1-kanr(见图1),使用SanPrep柱式DNA胶回收试剂盒(上海生工)进行胶回收,于-20℃保存备用。
1.3.2 回收产物的酶切及浓缩
ctfat1p1-kanr基因片段胶回收产物经限制性内切酶EcoRI酶切处理,并向酶切产物中加入1/10体积3 mol/L醋酸钠和2.5倍体积无水乙醇,并置于-20℃冰箱冷却1 h,后经12 000 r/min离心5 min得沉淀;加入300 μL体积分数为70%乙醇重悬清洗,12 000 r/min离心5 min除去乙醇,37℃风干30 min,最后加入15 μL ddH2O重悬DNA。使用核酸超微量分光光度计测定回收DNA浓度,并最终获得质量浓度在200~1000ng/μL之间的DNA溶液。于-20℃保存,以备后用。
1.3.3 电转感受态细胞制备
将C.tropicalis1798接种到含50 mL菌体增殖培养基的250 mL三角瓶中,30℃、200 r/min,摇床过夜培养;将过夜培养的菌液涂布,得到C.tropicalis1798单菌落;用接种环挑取C.tropicalis1798单菌落到50 mL菌体增殖培养基中,30℃、200 r/min,12 h后转接,培养10 h;取1.5 mL菌液到EP管中,3 000 r/min,离心1 min,收集菌体,用1.5 mL预冷的无菌水吹打悬浮细胞;3000r/min,离心1min,弃上清,用1 mL预冷的无菌水悬浮细胞;3 000 r/min,离心1 min,弃上清,用1 mL 1 mol/L预冷的山梨醇悬浮细胞;3 000 r/min,离心1 min,弃上清,用80 μL预冷的山梨醇悬浮细胞,即制成热带假丝酵母电转化感受态;将制备好的感受态细胞于-80℃保存,以备后用。
1.3.4 电转化
1.3.5 阳性重组菌株的鉴定
挑取上述菌落接种到含G418抗性的液体YEPD培养基中,30℃培养过夜后8 000 r/min离心收集菌体,使用上海生物工程有限公司提供的试剂盒提取重组菌DNA,并以获得的基因组为模板,Fat1p F1和Kan R2为引物进行PCR扩增,扩增产物利用1%琼脂糖凝胶电泳进行验证,最终获得阳性重组菌株。
1.3.6 原始菌株和重组菌株的发酵培养与二元酸的测定
取500μL甘油管保藏的C.tropicalis1798和C.tropicalis 1798△ctfat1p分别接种于50 mL液体YEPD培养基,30℃、200 r/min培养12 h。取种子培养液转接于50 mL发酵培养基(含葡萄糖)中,接种量为10%、30℃、200 r/min摇床培养,待生长稳定后调节发酵液pH值为7.5,加入10%油脂,进入产酸期。在产酸期,每12 h调节pH至7.5,产酸期6 d。
将20 mL发酵液,加入2 mL 4 mol/L的NaOH溶液,沸水浴5 min,混匀,冷却后置于50 mL离心管中,10 000 r/min离心10min。吸取中间的水相于50mL锥形瓶中,滴加3 mol/L的硫酸溶液至pH值为2~3,使二元酸完全结晶析出。抽滤,并用去离子水洗涤锥形瓶和滤饼,至滤液和滤纸呈中性为止。将滤饼及滤纸移入150 mL锥形瓶,加入30 mL体积分数为95%乙醇溶液作溶剂,加热使二元酸完全溶解,加入2~3滴溴百里香酚蓝作指示剂,用NaOH标准液滴定至终点。
1.3.7 原始菌株和重组菌株生长速率分析
以2%的接种量分别接种于以葡萄糖、十二烷、十六烷和油脂为碳源的50 mL发酵培养基,30℃、200 r/min振荡培养,每2 h取样检测其OD600nm值。绘制生长曲线进行对比。
2 结果与分析
2.1 氨基酸序列比对
父亲走后,我已习惯睡前不再将房门锁上。母亲几乎每夜都会来到我的房里,不同的是,她从不在我的书桌上留下任何字句,也从不扭亮任何一点灯光。我依旧像从前那样:在母亲转动门把的时候翻过身去面对墙壁,眯着双眼;我依然不敢贸然起身惊动母亲,依然没有勇气在那样的时刻里与母亲的眼神相对。
目前关于酵母中fat1p基因功能的研究主要集中在S.cerevisiae和Y.lipolytica中。通过NCBI分别检索S.cerevisiae中Ssfat1p(NCBI No.:AJQ08071.1,669 aa)和Y.lipolytica中Ylfat1p(NCBI No.:SEI35013.1,639aa)蛋白的氨基酸序列,并利用Geneious进行序列比对分析,结果如图2所示。由图2可知,C.tropicalis中Ctfat1p蛋白的氨基酸序列与Ssfat1p和Ylfat1p的一致性均为50%。但Ssfat1p和Ylfat1p的功能并不完全相同[21],因此C.tropicalis中的Ctfat1p蛋白的具体功能还需要进一步探究。

图2 氨基酸序列相似性分析Fig.2 Analysis of amino acid sequence similarity
2.2 基因敲除片段ctfat1p1-kanr的构建、电转化及鉴定
以C.tropicalis1798菌体的基因组DNA为模板,使用引物Fat1p F1和Fat1p R1进行PCR扩增,1%琼脂糖凝胶电泳检测,结果见图3(a)。由图3(a)可知,在500 bp左右出现特异性电泳条带,与理论长度551 bp相符,表明以成功获得用于ctfat1p基因敲除的同源臂ctfat1p1;以质粒pPIC9k为模板,使用引物Kan F2和Kan R2进行PCR扩增,1%琼脂糖凝胶电泳检测,结果见图3(b)。由图3(b)可知,在1500bp左右出现特异性电泳条带,与理论长度1 523 bp相符,表明已成功获得G418抗性基因片段kanr;以胶回收制得的ctfat1p1片段与制得的kanr片段为模板,Fat1p F1和Kan R2为引物进行PCR扩增进行重叠延伸PCR,制得基因敲除片段ctfat1p1-kanr,1%琼脂糖凝胶电泳检验PCR产物,结果见图3(c)。由图3(c)可知,在2 000 bp左右出现特异性电泳条带,与理论长度2 074 bp相符,表明基因敲除片段ctfat1p1-kanr构建成功。
使用EcoRI对300 μL融合PCR产物进行单酶切,并使用乙醇沉淀法浓缩获得酶切片段,使用核酸超微量分光光度计测定回收DNA质量浓度,结果显示,DNA质量浓度为532.18 ng/μL。将20 μL回收片段与80 μLC.tropicalis 1798感受态细胞混合并电转化,并使用引物KanF2和KanR2对含G418平板上生长菌落的基因组进行验证,1%琼脂糖凝胶电泳检验PCR产物,结果见图3(d)。由图3(d)可知,表明ctfat1p1-kanr已同源重组至C.tropicalis1798基因组中,并完成ctfat1p基因的敲除。

图3 PCR产物电泳图Fig.3 Electrophoresis of PCR products
2.3 以油脂为底物C.tropicalis1798和C.tropicalis1798△ctfat1p的发酵验证
菌体生长进入稳定期后加入10%油脂,每12 h调节pH到7.5,产酸期6 d。用标准氢氧化钠溶液滴定,测得二元酸的质量浓度,结果表明,C.tropicalis1798△ctfat1p比C. tropicalis1798产二元酸的量明显降低,其产量为1.83 g/L,比C.tropicalis1798产二元酸降低了65.3%。fat1p基因具有转运脂肪酸的功能,fat1p基因敲除后,菌体转化脂肪酸的效率降低从而影响二元酸的产量。测定C.tropicalis1798△ctfat1p二元酸的产量可初步推测ctfat1p基因的功能。
2.4 C.tropicalis1798和C.tropicalis1798△ctfat1p生长速率分析
产量降低的原因可能为ctfat1p基因敲除后减弱了细胞对脂肪酸的摄取,为进一步验证ctfat1p基因的功能,分别以葡萄糖、十二烷、十六烷和油脂为唯一碳源对C.tropicalis 1798和C.tropicalis1798△ctfat1p菌株菌体的生长状况进行测定。
将C.tropicalis1798和C.tropicalis1798△ctfat1p接种于50 mL发酵液体培养基,接种量为2%,30℃、200 r/min的条件下培养,每个菌做3个平行,每隔2 h取样并检测其OD600nm值绘制生长曲线。
由图4可知,以葡萄糖为唯一碳源时,两个菌的生长速度基本一致,10 h后生长均进入稳定期(见图4a),表明ctfat1p基因的敲除对葡萄糖的代谢影响不大;以十二烷为唯一碳源时,两个菌的生长速度基本一致,10 h后生长均进入稳定期,14 h后由于营养物质的缺乏导致菌体自溶,OD600nm值下降,进入衰退期(见图4b),而以十六烷为唯一碳源时,两个菌的生长速度基本一致,12 h后生长均进入稳定期,14 h后由于缺乏营养物质导致菌体自溶,OD600nm值下降,进入衰退期(见图4c),表明ctfat1p基因的敲除对烷烃的代谢影响不大;以油脂为唯一碳源时,C.tropicalis1798△ctfat1p的生长速度明显滞后,生长缓慢(见图4d),培养12 h时重组菌OD600nm仅为野生型菌株的47.9%,由此可知,ctfat1p基因为热带假丝酵母油脂吸收的关键基因。

图4 C.tropicalis1798和C.tropicalis1798△ctfat1p生长曲线Fig.4 Growth curve ofC.tropicalis1798 andC.tropicalis1798△ctfat1p
3 结论
本实验利用PCR扩增技术获取热带假丝酵母ctfat1p基因同源臂ctfat1p1、抗性标签G418基因kanr,借助重叠延伸PCR技术构建热带假丝酵母ctfat1p基因敲除载体(ctfat1p1-kanr),通过一次同源重组即可实现目的基因的敲除;构建的敲除片段仅包含两段序列即同源臂和抗生素抗性标签基因,构建流程简单,基因敲除周期短、不依赖于敲除载体,并有效降低敲除成本,提高工作效率。
对ctfat1p基因敲除菌株经过初步发酵研究发现:以油脂为底物发酵生产二元酸时重组菌株C.tropicalis1798△ctfat1p比原始菌株C.tropicalis1798产二元酸的量明显降低,其产量为1.83 g/L,比C.tropicalis1798产二元酸降低了65.3%;在以葡萄糖、十二烷、十六烷为唯一碳源时对细胞的生长影响很小。在生长初期,突变株的生长速度略慢于原始菌株,这与酿酒酵母类似[22-23],说明ctfat1p基因对于维持热带假丝酵母正常生长起到一定的作用,敲除后细胞的生长能力减弱。在以油脂为唯一碳源时突变株的生长速度明显滞后,即不能有效利用油脂作碳源,细胞转运长链脂肪酸的能力变弱。因此,推测C.tropicalis1798中的ctfat1p基因为油脂吸收的关键基因。
此外,热带假丝酵母虽然具有有性生殖的过程[24],但此过程较难发生,且其单倍体阶段不能长期稳定存在,最终会转变成二倍体。优良性状的纯合二倍体可以减少变异的发生,维持稳定的发酵性能。因此,获取酵母的单倍体对于热带假丝酵母这类二倍体生物的菌种改良工作来说是十分重要的。本实验仅对C.tropicalis1798中的ctfat1p基因进行了单拷贝敲除,后续实验中需要构建双敲除载体对ctfat1p基因进行双拷贝敲除和增加ctfat1p基因拷贝数,进一步验证ctfat1p基因的功能。
[1]任刚,陈远童.十二碳二元酸的发酵研究[J].生物工程学报,2000,16(2):198-202.
[2]项峥,陈献忠,张利华,等.利用可重复使用的URA3标记基因建立热带假丝酵母基因敲除系统[J].遗传,2014,36(10):1053-1061.
[3]桂秋芬,姚嘉旻,蒋洋松,等.利用热带假丝酵母发酵生产长链二元酸的研究进展[J].化学与生物工程,2014,31(1):17-22.
[4]BLANDIN G,OZIER-KALOGEROPOULOS O,WINCKER P,et al.Genomic exploration of the hemiascomycetous yeasts:16.Candida tropicalis[J].FEBS Lett,2000,487(1):91-94.
[5]RACHINSKI V V,DAVIDOVA E G,LAPOTYSHKINA A I.Localization of n-paraffin oxidation by yeasts[J].Dokl Akad Nauk Sssr,1971, 200(2):457-460.
[6]KAEPPELI O,FIECHTER A.The mode of interaction between the substrate and cell surface of the hydrocarbon-utilizing yeastCandida tropicalis[J].Biotechnol Bioeng,1976,18(7):967-974.
[7]张志禹,朱涛,林荣胜,等.热带假丝酵母的二元酸分泌现象及其动力学[J].清华大学学报:自然科学版,1999,39(12):27-30.
[8]高弘.生物催化生产十三碳二元酸中β-氧化途径的代谢调控[D].北京:清华大学,2005.
[9]SCHWENK R W,HOLLOWAY G P,LUIKEN J J,et al.Fatty acid transport across the cell membrane:regulation by fatty acid transporters[J]. Prostag Leukotr Ess Fatty Acid,2010,82(4-6):149-154.
[10]齐仁立,黄金秀,杨飞云,等.脂肪酸转运蛋白家族及其介导的脂肪酸跨膜转运[J].动物营养学报,2013,25(5):905-911.
[11]COE N R,SMITH A J,FROHNERT B I,et al.The fatty acid transport protein(FATP1)is a very long chain acyl-CoA synthetase[J].J Biol Chem,2000,274(51):36300-363004.
[12]DULERMO R,GAMBOA-MELENDEZ H,DULERMO T,et al.The fatty acid transport protein Fat1p is involved in the export of fatty acids from lipid bodies inYarrowia lipolytica[J].Fems Yeast Res,2014,14 (6):883-896.
[13]BLACK P N,DIRUSSO C C.Transmembrane movement of exogenous long-chain fatty acids:proteins,enzymes,and vectorial esterification[J]. Microbiol Molecul Biol Rev,2003,67(3):454-472.
[14]FAERGEMAN N J,DIRUSSO C C,ELBERGER A,et al.Disruption of theSaccharomyces cerevisiaehomologue to the murine fatty acid transport protein impairs uptake and growth on long-chain fatty acids[J].J Biol Chem,1997,272(13):8531-8538.
[15]ZOU Z,DIRUSSO C C,CTMACTA V,et al.Fatty acid transport in Saccharomyces cerevisiae[J].J Biol Chem,2002,277(34):31062-31071.
[16]A MORETTI,L BONCIO,B POSTERARO,et al.Co-cutaneous infection in a dog:PCR-reverse identification ofCandida tropicalis,on skin biopsy[J].J De Mycol Médicale,2006,16(1):30-36.
[17]QIU D,LIU X,WANG J,et al.Artificial synthesis of TAT PTD-Tachyplesin fusion gene by overlap extension PCR[J].Agr Biotechnol,2013 (3):14-17.
[18]MENZIES M,SEIM I,JOSH P,et al.Cloning and tissue distribution of novel splice variants of the ovine ghrelin gene[J].Bmc Vet Res,2014, 10(1):1-9.
[19]F.M.奥斯博,R.E.金士顿,等.精编分子生物学实验指南[M].北京:科学出版社,1998:25-30.
[20]邢竹青,王彦宁,刘兆贤,等.Lactobacillus kefiranofaciens乳糖酶基因克隆及在毕赤酵母中表达[J].中国酿造,2016,35(1):10-13.
[21]DULERMO R,GAMBOAMELENDEZ H,DULERMO T,et al.The fatty acid transport protein Fat1p is involved in the export of fatty acids from lipid bodies inYarrowia lipolytica[J].FEMS Yeast Res,2014,14(6):883-896.
[22]WINZELER E A,LIANG H,SHOEMAKER D D,et al.Functional analysis of the yeast genome by precise deletion and parallel phenotypic characterization[J].Nov Found Sym,2000,229:105-111.
[23]LIN S S,MANCHESTER J K,GORDON J I.Enhanced gluconeogenesis and increased energy storage as hallmarks of aging inSaccharomyces cerevisiae[J].J Biol Chem,2001,276(38):36000-36007.
[24]PORMAN A M,ALBY K,HIRAKAWA M P,et al.Discovery of a phenotypic switch regulating sexual mating in the opportunistic fungal pathogenCandidatropicalis[J].Proceed Nat Acad Sci,2011,108(52):21158-21163.
Function of genectfat1pinCandida tropicalison the absorption of aliphatic compounds
SHI Ying,YANG Xiaohui,MA Chunling,WANG Tengfei,XIAO Jing,WANG Ruiming,WANG Junqing*
(Shandong Provincial Key Laboratory of Microbial Engineering,School of Bioengineering,Qilu University of Technology, Jinan 250353,China)
Using the genectfat1pinCandida tropicalis1798 as research object,the 500 bp genectfat1pfragment was connected with G418 resistance gene(kanr)by using overlapping PCR.After a single enzyme digestion at the end of the fragment,then the recombinant gene fragment was transformed intoC.tropicalis1798 competent cells.Through a homologous single exchange,the resistance genekanrwas inserted into the genectfat1p,and then the target gene was knocked successfully.The function of Ctfat1p in the process of fatty acid transmembrane-movement ofC.tropicaliswas analyzed by fermentation experiments.The results showed that a genectfat1pdeletion strain(namedC.tropicalis1798Δctfat1p)was obtained successfully by G418 resistance screening and genome PCR identification.The analysis was founded that the OD600nmvalue ofC.tropicalis1798Δctfat1pwith oil as the substrate culturing for 12 h was only 47.9%of the wild type strain,which indicated that deletion of genectfat1paffected absorption and utilization ofC.tropicalis1798 on oil.The establishment of gene ctfat1pdeletion strain by the gene knockout method could weaken intake of cell on long chain fatty acid,which verified that the genectfat1pwas a key gene of the absorption and utilization ofC.tropicalison oil.
genectfat1p;Candida tropicalis;gene knockout;homologous single exchange
Q815
0254-5071(2017)03-0132-06
10.11882/j.issn.0254-5071.2017.03.027
2016-12-14
山东省自主创新及成果转化专项(No.201422CX02602)
石莹(1990-),女,硕士研究生,研究方向为微生物酶技术。
*通讯作者:汪俊卿(1988-),男,讲师,博士,研究方向为发酵工程和微生物酶技术。
猜你喜欢
昆明医科大学学报(2022年1期)2022-02-28
昆明医科大学学报(2021年4期)2021-07-23
海南医学(2021年9期)2021-05-26
铁路通信信号工程技术(2021年3期)2021-04-03
肿瘤防治研究(2020年8期)2020-09-11
女报(2020年7期)2020-08-17
中国感染控制杂志(2020年1期)2020-02-05
合成树脂及塑料(2020年3期)2020-01-16
儿童故事画报(2019年8期)2019-08-14
艺术启蒙(2018年8期)2018-08-22
