
内侧杏仁核中催产素对布氏田鼠社会行为的影响
2017-04-08 07:18张楠闫立新高杰孙秀萍宋铭晶
四川动物 2017年2期
张楠, 闫立新, 高杰, 孙秀萍, 宋铭晶
(中国医学科学院医学实验动物研究所,北京协和医学院比较医学中心,卫生部人类疾病比较医学重点实验室,北京100021)
内侧杏仁核中催产素对布氏田鼠社会行为的影响
张楠, 闫立新, 高杰, 孙秀萍, 宋铭晶*
(中国医学科学院医学实验动物研究所,北京协和医学院比较医学中心,卫生部人类疾病比较医学重点实验室,北京100021)
目的 探究内侧杏仁核(MeA)脑区中催产素(OT)对布氏田鼠Lasiopodomysbrandtii社会行为的影响。方法 在雄性布氏田鼠MeA区注射催产素受体拮抗剂后,采用社会识别实验、中央竞技场实验确定MeA区催产素受体的变化对其社会行为的影响。结果 社会识别实验发现在MeA区注射人工脑脊液的布氏田鼠(对照组)对陌生鼠的探索时间显著长于探索熟悉鼠(0.001
布氏田鼠;催产素;杏仁核;社会记忆;亲社会性
无论是人还是动物都有一部分个体或物种表现出较多的社会友善行为,一部分表现出较多的反社会行为。啮齿动物既有以家族为单位群居的,表现出较多社会互助行为,如社会性强的橙腹田鼠Microtusochrogaster和布氏田鼠Lasiopodomysbrandtii等(Olazabal & Young,2006;Wangetal.,2011),也有独居的,在繁殖期和非繁殖期都有很强攻击性的山地田鼠Microtusmontanus和大仓鼠Tscherskiatriton(Liuetal.,2001;Panetal.,2010)。Pan等(2016)研究发现群居的橙腹田鼠在遇到种群内其他个体时表现出更多的探究行为,即社会探究行为,而独居的山地田鼠表现出对空笼的探究行为超过对其他个体的探究行为。鉴于不同啮齿动物社会行为的巨大差异,通过对其特定脑区的基因进行调控,改变其社会行为,可以研究该动物社会行为的遗传机制。
催产素(oxytocin,OT)是哺乳动物分泌的一种环状九肽分子神经激素,以其在分娩和泌乳中起神经调节作用而闻名,研究发现其在调节社会认知行为方面的影响显著,其中包括焦虑、学习记忆、社会认知、配偶选择等行为(孟雅婧等,2014;Shen,2015)。OT及其受体在动物脑区内广泛表达,不过目前的研究显示其在不同脑区的表达会影响不同的社会行为,已有研究通过对棕色田鼠特定脑区内侧杏仁核(medial amygdala,MeA)和伏隔核区OT的表达进行调控,发现雄性棕色田鼠的育幼行为会随着OT的表达减少而减弱(Caoetal.,2014)。Wang等(2015)对不同生殖经验的雄性棕色田鼠MeA脑区OT的研究发现OT及催产素受体(oxytocin receptor,OTR)的表达与棕色田鼠的生殖经验和亲子行为呈正相关。
布氏田鼠社会行为发达,是一种研究动物亲社会行为较为理想的模型动物,已有研究对群居的布氏田鼠与独居的大仓鼠脑区OT的分布进行检测,发现在MeA等脑区,群居的布氏田鼠OT的分布明显多于独居的大仓鼠(Xuetal.,2010),而本实验室对布氏田鼠与BALB/c小鼠不同脑区的OT及其受体的Western Blot检测显示布氏田鼠MeA区OT分布较多,且有研究表明脑部MeA区与社会识别行为有密切联系(Wangetal.,2013)。社会识别行为以社会记忆能力为生理基础,个体的社会记忆是个体间进行社会交往所必需的,是某个体在同物种内能够区分熟悉个体与陌生个体的能力,是种群内所有社会关系形成的基础(Fergusonetal.,2002)。为了明确大脑MeA区OT表达对动物社会行为和社会性的影响,本研究调控布氏田鼠MeA区OT的表达,并进行行为学分析,揭示动物社会行为的遗传调控机制。
1 材料与实验方法
1.1 材料
1.1.1 实验动物 4月龄雄性布氏田鼠60只,体质量45~55 g,均为中国医学科学院医学实验动物研究所饲养、繁育并净化。10只雄性3月龄SPF级BALB/c小鼠,体质量约25 g,购自北京维通利华实验动物技术有限公司,实验动物生产许可证号:SYXK(京)2012-0022。动物饲养在独立送排风净化动物笼。饮用水、垫料和笼具均经高压灭菌处理,饲料经60Co照射消毒达到SPF级动物饲料标准,光照时间14L∶10D。实验动物的使用通过了中国医学科学院医学实验动物研究所的实验动物使用和管理委员会批准,批准编号:ILAS-PG-2014-011。
1.1.2 仪器 脑立体定位仪,旷场装置(长50 cm×宽50 cm×高30 cm),三箱装置(长105 cm×宽35 cm×高35 cm),动物行为学视频跟踪系统(Noldus Information Technology,EthoVision XT,The Netherlandtiaokongs)。
1.2 实验方法
1.2.1 Western Blot检测 雄性布氏田鼠(n=6)和雄性BALB/c小鼠(n=6)取脑,扣取MeA区脑组织,由于单只MeA区脑组织较少,故将扣取的布氏田鼠和小鼠脑组织分别混合,提取蛋白,Western Blot检测其OT与OTR的本底表达量,对比布氏田鼠MeA脑区OT与OTR的表达量及其与小鼠的差异。
1.2.2 埋管注射 布氏田鼠(n=46)经水合氯醛麻醉(5%,0.6 mL/100 g)后固定在脑立体定位仪上,确认脑部前囟为零点坐标(X,Y,Z),根据MeA区三维坐标(X:-1.8 mm,Y:±2.3 mm,Z:-6.3 mm)开颅钻打孔,双侧脑部埋管,导管深入脑下6.3 mm,进入MeA脑区。套管通过螺丝和牙科水泥固定于头骨上,不进行注射时,用导管帽进行封闭。待伤口愈合,手术10 d后微量进样器给药,然后进行行为学实验。注射试剂:实验组(n=23)双侧注射催产素受体拮抗剂(oxytocin receptor antagonist,OTRA;Sigma,L-368899),注射剂量为单侧750 ng(Arakawaetal.,2010),注射速度0.1 μL·min-1,总体积0.5 μL。注射后留针2 min,使药物充分扩散,防止倒流;对照组(n=23)注射相同体积的人工脑脊液(artificial cerebrospinal fluid,ACSF)。
1.2.3 社会识别实验 三箱测试是在社会识别或社会新奇实验中用来评估个体社交能力和对新事物的兴趣程度的方法。实验过程参考Page等(2009)的实验步骤并加以修改,测试区域由3个相邻的箱体组成(长105 cm×宽35 cm×高35 cm),中间2个透明塑料分隔器分隔,并通过开放的门道使三箱连通起来。实验对象为实验组布氏田鼠(n=12)和对照组布氏田鼠(n=12)。实验分为3个阶段:
(1)第一阶段(适应阶段):社会交互箱两侧箱体空置,中间的2个隔板门关闭,将实验鼠放入中间箱体,适应10 min。
(2)第二阶段:适应结束后,同一时间段进行测试期。测试过程中,左侧箱体放入空笼,右侧箱体的笼中放入同性幼年刺激鼠。将实验鼠迅速放入中间箱体中,撤掉隔板,立即开始测试,动物跟踪系统自动记录实验鼠进入各箱的时间、次数、路程,接触空鼠笼和刺激鼠的潜伏期、时间,跟踪时间为10 min。10 min后将实验鼠取出。
(3)第三阶段:步骤(2)中所用刺激鼠作为熟悉鼠,同时,在左侧箱体的笼中,放入另外一只陌生刺激鼠,观察10 min,采集数据指标同步骤(2)。每只实验鼠结束实验后都需要将实验鼠的粪便清除干净,并用75%的酒精再次擦拭,掩盖老鼠的味道,待酒精挥发完后再进行下一只实验。
1.2.4 中央竞技场实验 中央竞技场实验参照李广宇(2012)的方法,在旷场箱中间设置一个能上下抽拉的隔板,将旷场箱分为左右相等的两部分。实验于暗光周期1 h后开始,行为过程采用摄像记录设备(夜视模式)记录后进行分析,实验时将实验鼠与刺激鼠分别置于中央竞技场隔板的两侧,并给予其5 min适应环境,每组实验时间为20 min,每次实验完成后清理箱底,用75%酒精擦拭箱底和箱壁,待晾干之后再进行下一组实验。实验对象为实验组布氏田鼠(n=11)和对照组布氏田鼠(n=11)。
1.3 数据分析
行为学数据采用SPSS 20.0进行统计学处理,用独立样本t检验分析MeA区OT对社会识别实验和中央竞技场实验数据的影响。P<0.05为差异有统计学意义,P<0.01为差异有高度统计学意义,数据以平均值±标准误(Mean±SE)表示,得到的结果用GraphPad Prism 6作图。
2 实验结果
2.1 Western Blot检测
Western Blot检测显示在雄性布氏田鼠MeA区OT的表达量远高于雄性BALB/c小鼠(图1:A),雄性布氏田鼠MeA区OTR的表达量与雄性BALB/c小鼠没有明显差异(图1:B)。

图1 Western Blot检测OT与OTR的蛋白表达
A. 雄性布氏田鼠与雄性BALB/c小鼠MeA区OT蛋白相对表达量的对比, B. 雄性布氏田鼠与雄性BALB/c小鼠MeA区OTR蛋白相对表达量的对比, 用GAPDH蛋白做对照, 比较OT或OTR与GAPDH灰度的比值;***P<0.001。
A. comparison of OT protein relative expression in the MeA between the maleLasiopodomysbrandtiiand the male BALB/c mice, B. comparison of OTR protein relative expression in the MeA between the maleL.brandtiiand the male BALB/c mice using GAPDH as control;***P<0.001.
2.2 社会识别实验
由于脑部埋管注射位置有偏差,通过脑部切片确认位置,统计分析注射正确位置样本的行为:实验组,n=8;对照组,n=7。对照组和实验组布氏田鼠对有刺激鼠的笼探索时间多于对空笼的探索时间,且差异有高度统计学意义(0.001
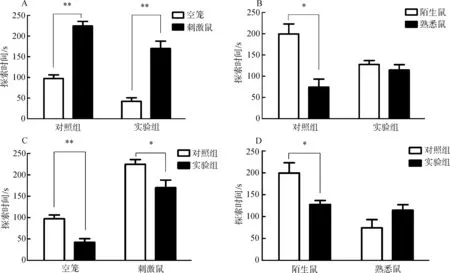
图2 实验组和对照组社会识别实验
Fig. 2 Social recognition experiment of experimental group and control group
A.对空笼和刺激鼠探索时间, B. 对熟悉鼠和陌生鼠探索时间, C. 对空笼和刺激鼠探索时间的组间对比, D. 对陌生鼠和熟悉鼠探索时间的组间对比;*P<0.05,**P<0.01。
A.the exploration time between empty and stimulus, B. the exploration time between novel and familiar, C. the exploration time of empty and stimulus between the groups, D. comparison the exploration time of novel and familiar between the groups;*P<0.05,**P<0.01.
2.3 中央竞技场实验
由于脑部埋管注射位置有偏差,通过脑部切片确认位置,统计分析注射正确位置样本的行为:实验组,n=7;对照组,n=7。实验组布氏田鼠的攻击潜伏期和攻击时长与对照组的差异均无统计学意义(P>0.05)(图3)。
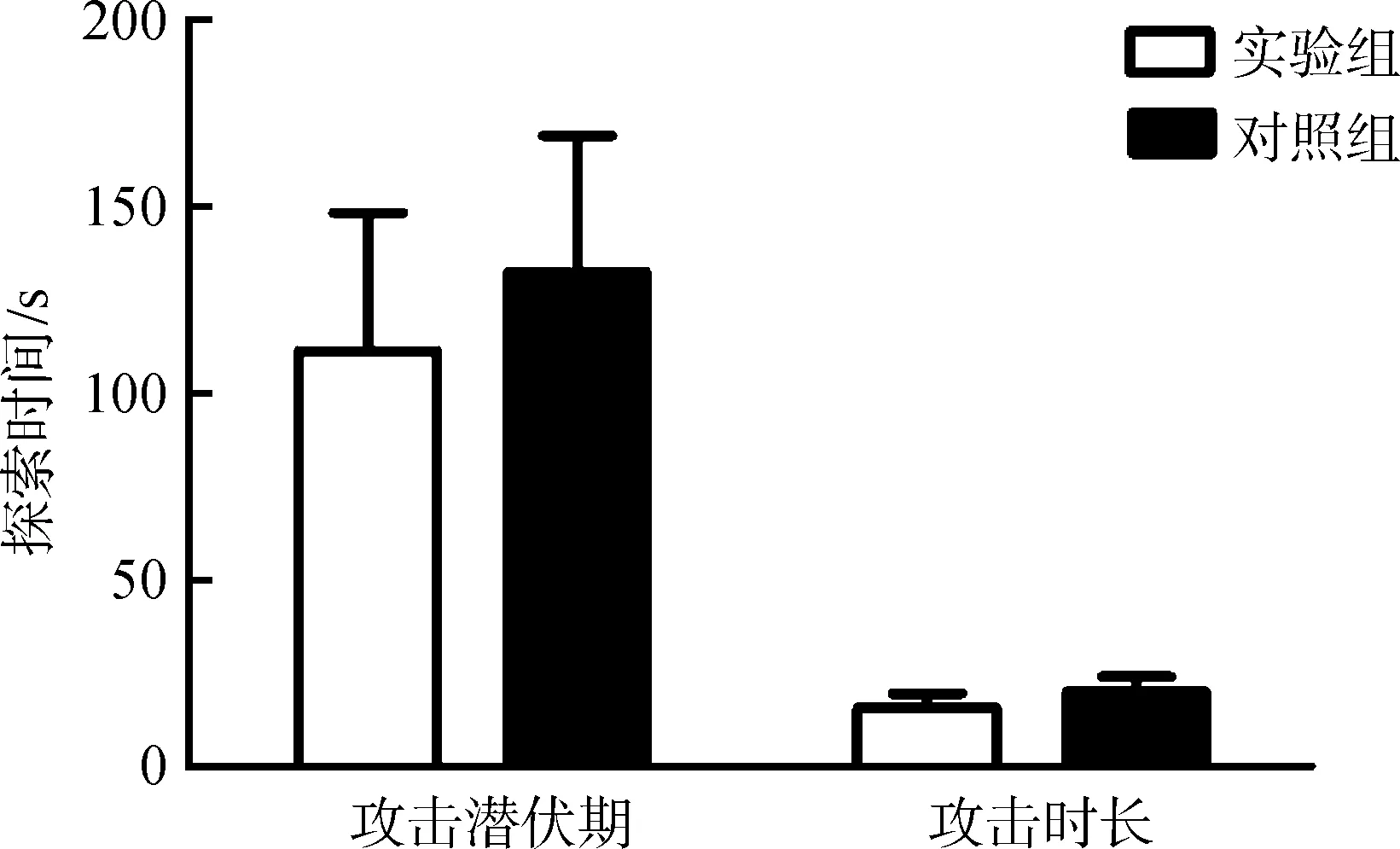
图3 催产素受体抑制剂对布氏田鼠攻击行为的影响
3 讨论
识别一个个体并对此个体形成一个社会记忆的过程是后期社会关系形成的一个主要步骤。社会识别实验是基于啮齿类动物探索新物体(或者个体)的时间要多于探索熟悉物体(个体)的时间这种现象而成立的(Winslow,2003)。MeA区注射ACSF的对照组布氏田鼠对陌生鼠的探索时间明显高于对熟悉鼠的探索时间,而注射OTRA的实验组布氏田鼠对陌生鼠和熟悉鼠的探索时间的差异无统计学意义,说明MeA区内OTR的抑制能影响布氏田鼠的社会行为。这种社会行为的改变可能有2个原因:一是由于社会记忆形成的障碍造成的;二是动物亲社会性的改变造成的。Pan等(2016)对独居的山地田鼠和群居的橙腹田鼠分别做社会识别实验,发现独居的山地田鼠对空笼探索的时间比对同种的其他个体更长,而且对熟悉个体和陌生个体的探索时间没有显著差异,推测因为独居动物的亲社会性水平低,所以它们更关注物体,而不是同种的陌生或熟悉个体,缺少探究行为。而野外实验研究也报道独居鼠在自然状态下遭遇时多回避或攻击(张健旭等,1999)。本实验结果显示,实验组和对照组雄性布氏田鼠在面对空笼和刺激鼠时,对刺激鼠的探索时间都显著高于对空笼的探索时间,初步说明MeA区OT阻断不影响雄性布氏田鼠的亲社会性。而对不同组间进行比较发现,注射OTRA的实验组布氏田鼠对空笼、刺激鼠以及陌生鼠的探索时间都明显低于注射ACSF的对照组布氏田鼠,说明OTRA能够降低布氏田鼠的社会探索行为。以上实验都证明在MeA区注射OTRA能够减少其社会探索行为。但是这种社会行为的改变是由于社会记忆障碍造成的还是亲社会性的改变造成的还需要进一步确定。因此为了进一步确定布氏田鼠的亲社会性有无改变,我们用中央竞技场的实验进行验证。结果证明,实验组与对照组相比,其攻击潜伏期和攻击时长都没有明显差异,表明OT在MeA区的阻断对布氏田鼠的主动攻击行为没有影响。综上所述,OT在MeA的表达是通过影响布氏田鼠的社会记忆,进而影响其社会行为,而不是改变布氏田鼠的亲社会性。
已有研究证明OT在不同脑区表达变化与自闭症、抑郁症等精神类疾病的形成有关(Penagarikanoetal.,2015)。Shen(2015)对近年OT的研究进行总结,发现有关OT对自闭症影响的研究中,OT的治愈能力不仅能在动物实验中呈现,在临床治疗中也得到了肯定,研究中还发现OT的剂量效应呈“U”型曲线,推测OT对精神类疾病的作用比较复杂,有待进一步探究。现有有关OT动物实验与临床治疗的研究表明,OT的作用随其表达脑区的位置和浓度梯度变化均有所改变,所以我们希望找到调控社会行为的主要脑区和OT对社会行为的调控机制,本实验室和其他研究都已经证明OT与社会行为有关,但要确定影响社会行为的主要脑区,还需要对布氏田鼠的不同脑区进行比较研究和基因调控才能实现。
李广宇. 2012. 配偶选择对黑线仓鼠脑内Dadr2表达的影响[D]. 山东曲阜: 曲阜师范大学.
孟雅婧, 杨洋, 刘莹, 等. 2014. 催产素受体配体及其中枢和外周功能研究进展[J]. 中国药理学通报, (6): 752-756.
张健旭, 张知彬, 王祖望. 1999. 大仓鼠在繁殖期的行为关系及交配行为[J]. 兽类学报, (2): 52-55.
Arakawa H, Arakawa K, Deak T. 2010. Oxytocin and vasopressin in the medial amygdala differentially modulate approach and avoidance behavior toward illness-related social odor[J]. Neuroscience, 171(4): 1141-1151.
Cao Y, Wu R, Tai F,etal. 2014. Neonatal paternal deprivation impairs social recognition and alters levels of oxytocin and estrogen receptor alpha mRNA expression in the MeA and NAcc, and serum oxytocin in mandarin voles[J]. Hormones & Behavior, 65(1): 57-65.
Ferguson JE, Head BH, Frank FH,etal. 2002. Misoprostol versus low-dose oxytocin for cervical ripening: a prospective, randomized, double-masked trial[J]. American Journal of Obstetrics and Gynecology,187(2): 273-280.
Liu Y, Fowler CD, Wang Z. 2001. Ontogeny of brain-derived neurotrophic factor gene expression in the forebrain of prairie and montane voles[J]. Developmental Brain Research, 127(1): 51-61.
Olazabal DE, Young LJ. 2006. Oxytocin receptors in the nucleus accumbens facilitate “spontaneous” maternal behavior in adult female prairie voles[J]. Neuroscience, 141(2): 559-568.
Page DT, Kuti OJ, Sur M. 2009. Computerized aassessment of social approach behavior in mouse[J]. Frontiers in Behavioral Neuroscience,3: 48.
Pan Y, Liu Y, Lieberwirth C,etal. 2016. Species differences in behavior and cell proliferation/survival in the adult brains of female meadow and prairie voles[J]. Neuroscience, 315: 259-270.
Pan Y, Xu L, Young KA,etal. 2010. Agonistic encounters and brain activation in dominant and subordinate male greater long-tailed hamsters[J]. Hormones and Behavior, 58(3): 478-484.
Penagarikano O, Lazaro MT, Lu XH,etal. 2015. Exogenous and evoked oxytocin restores social behavior in the Cntnap2 mouse model of autism[J]. Science Translational Medicine, 7(271): 271r-278r.
Shen H. 2015. Neuroscience: the hard science of oxytocin[J]. Nature, 522(7557): 410-412.
Wang B, Li Y, Wu R,etal. 2015. Behavioral responses to pups in males with different reproductive experiences are sssociated with changes in central OT, TH and OTR, D1R, D2R mRNA expression in mandarin voles[J]. Hormones & Behavior, 67: 73-82.
Wang D, Li D, Liu M,etal. 2011. Behavioral evaluation of quinestrol as a sterilant in male Brandt’s voles[J]. Physiology & Behavior, 104(5): 1024-1030.
Wang Y, Xu L, Pan Y,etal. 2013. Species differences in the immunoreactive expression of oxytocin, vasopressin, tyrosine hydroxylase and estrogen receptor alpha in the brain of mongolian gerbils (Merionesunguiculatus) and Chinese striped hamsters (Cricetulusbarabensis)[J]. PLoS ONE, 8(6): e65807. DOI:10.1371/journal.pone.0065807.
Winslow JT. 2003. Mouse social recognition and preference[J]. Current Protocols in Neuroscience, 8: 8-16.
Xu L, Pan Y, Young KA,etal. 2010. Oxytocin and vasopressin immunoreactive staining in the brains of Brandt’s voles (Lasiopodomysbrandtii) and greater long-tailed hamsters (Tscherskiatriton)[J]. Neuroscience, 169(3): 1235-1247.
Effect of Oxytocin in the Medial Amygdala on the Social Behavior ofLasiopodomysbrandtii
ZHANG Nan, YAN Lixin, GAO Jie, SUN Xiuping, SONG Mingjing*
(Key Laboratory of Human Diseases Comparative Medicine, Ministry of Health, Institute of Laboratory Animal Science, Chinese Academy of Medical Sciences & Comparative Medicine Center, Peking Union Medical College, Beijing 100021, China)
Objective To explore the influence of oxytocin (OT) in the medial amygdala (MeA) on the social behaviors of Brandt’s voles (Lasiopodomysbrandtii). Methods Oxytocin receptor antagonist (OTRA) was injected into the MeA of male Brants’s voles, and then the social behaviors were tested using social recognition experiment and neutral arena encounter experiment. Results The control group (injected with artificial cerebrospinal fluid) took more time to explore the strangers than the familiars (0.001
Lasiopodomysbrandtii; oxytocin; medial amygdala; social memory; prosocial
2016-10-21 接受日期:2016-12-15
国家自然科学基金项目(31301890); 农业虫害鼠害综合治理研究国家重点实验室开放研究基金项目(Chinese IPM1615)
张楠(1991—), 男, 硕士研究生, 主要从事实验动物学研究
*通信作者Corresponding author, E-mail:songmj@cnilas.org
10.11984/j.issn.1000-7083.20160288
Q959.837; Q95-33
A
1000-7083(2017)02-0193-05
猜你喜欢
作文大王·低年级(2021年11期)2021-12-22
小天使·一年级语数英综合(2021年2期)2021-05-08
小学阅读指南·低年级版(2020年9期)2020-10-12
小学生(看图说画)(2020年4期)2020-04-01
阅读与作文(小学低年级版)(2019年11期)2019-12-26
中国医药指南(2017年30期)2017-01-15
飞碟探索(2016年5期)2016-05-10
浙江农业科学(2016年11期)2016-05-04
中国人兽共患病学报(2015年8期)2015-05-09
中国人兽共患病学报(2015年8期)2015-01-24
