
玉米蛋白粉替代鱼粉对大黄鱼生长、血清生化指标及肝脏组织学的影响
2017-05-16 09:07何娇娇王建娄宇栋党慧邓
水生生物学报 2017年3期
何娇娇王 萍,冯 建娄宇栋党 慧邓 蓉
(1. 浙江海洋大学浙江省海洋养殖装备与工程技术重点室, 舟山 316000; 2. 宁波大学海洋学院, 宁波 315211)
玉米蛋白粉替代鱼粉对大黄鱼生长、血清生化指标及肝脏组织学的影响
何娇娇1王 萍1,2冯 建1娄宇栋1党 慧1邓 蓉1
(1. 浙江海洋大学浙江省海洋养殖装备与工程技术重点室, 舟山 316000; 2. 宁波大学海洋学院, 宁波 315211)
为探讨玉米蛋白粉替代鱼粉对大黄鱼(Larimichthys crocea) 幼鱼生长、血清生化指标及肝脏组织形态的影响, 进行了为期56d的养殖试验, 探索玉米蛋白粉替代大黄鱼幼鱼饲料鱼粉的适当比例。以初始体重为(10.49±0.03) g的大黄鱼幼鱼为研究对象, 用玉米蛋白粉替代基础饲料(含40%鱼粉)0、15%、30%、45%、60%和75%的鱼粉来配制6种等氮(粗蛋白含量45%)等脂(粗脂肪含量10%)的实验饲料, 分别标记为C0、C15、C30、C45、C60、C75组。除C0以外的替代组分别添加了适量的晶体氨基酸(赖氨酸和蛋氨酸)。结果表明,玉米蛋白粉替代水平对大黄鱼幼鱼存活率、试验鱼特定生长率、饲料系数均无显著性差异(P>0.05)。C45和C60组肌肉粗蛋白含量显著高于C0组(P<0.05), 肌肉粗脂肪含量C45组显著高于C0、C15和C75组 (P<0.05), C45、C60和C75组肌肉水分显著低于C0组(P<0.05), 全鱼粗蛋白、粗脂肪、水分含量无显著差异(P>0.05); 灰分含量有上升趋势, C75组显著高于其他组(P<0.05)。各替代组的血清白蛋白、白球比、谷草转氨酶、葡萄糖均无显著性差异(P>0.05); 随着替代比例的升高, 总胆固醇有降低的趋势, 除C15组与C0无显著性差异外(P>0.05), 4组均显著低于CO组(P<0.05); 实验探讨替代后对鱼的影响, C75组血清总蛋白和球蛋白含量显著低于C0组(P<0.05); C75组血清谷草转氨酶含量显著高于C0(P<0.05)。各处理组总抗氧化能力、过氧化氢酶、谷胱甘肽过氧化氢酶均没有显著性影响(P>0.05), 但替代组总抗氧化能力含量均高于C0; 丙二醛在C75组显著高于C0组(P<0.05)。通过肝组织学观察表明, 当替代比例高于45%时, 呈现肝细胞核偏位、胞浆内脂肪滴较多、细胞透明空泡化等症状。综上所述, 在实验条件下, 研究认为玉米蛋白粉替代鱼粉对大黄鱼幼鱼的适宜添加量为45%。
大黄鱼; 玉米蛋白粉; 生长性能; 血清生化指标; 抗氧化; 肝脏组织学
鱼粉是水产饲料优质的蛋白源, 其含有丰富且较为平衡的必需氨基酸和脂肪酸; 抗营养因子少且适口性好, 不仅可消化能高而且还含有许多如烟酸、小肽、胆碱等特殊免疫蛋白等, 能够满足水产鱼类的营养需求[1]。然而, 考虑到鱼粉的昂贵价格和鱼粉供应需求量不稳定等主要因素, 国内外学者对植物蛋白源替代鱼粉进行了大量研究[2—4]。
玉米蛋白粉是以玉米为原料, 在制作过程中通过分离及脱脂后除去淀粉、胚芽、外皮产生的副产品。淀粉和纤维含量较低、不含抗营养因子、蛋白质含量(60%—70%)高于鱼粉和豆粕中的蛋白质含量、含有丰富的维生素B和维生素E且玉米蛋白粉所含必需氨基酸总量比大豆粉和鱼粉中的必需氨基酸总量高, 其中总含硫氨基酸和亮氨酸含量大于大豆粉和鱼粉中的含量[5,6]。据报道, 玉米蛋白粉在大菱鲆(Scophthalmus maximus L.)[5,7]、凡纳滨对虾(Litopenaeus vannamei)[8]中的添加量可以达到33%—70%。这些研究表明玉米蛋白粉的适宜添加对水产动物的生长不产生负面影响。因此, 玉米蛋白粉可作为水产动物饲料鱼粉的替代蛋白源。
大黄鱼(Larimichthys crocea)是我国的一种具有高营养高商业价值的鲈形目(Perciformes)、石首鱼科(Sciaenidae)洄游类鱼, 对饲料蛋白质含量要求高。由于大黄鱼肉质鲜嫩, 具有重要的商业价值,在我国沿海地带兴起了养殖热潮。目前, 大黄鱼饲料鱼粉的替代研究只有少量报道[9—11], 研究表明,鱼粉替代的适宜比例因选择的蛋白源不同而有差异。本文用玉米蛋白粉对大黄鱼幼鱼饲料鱼粉进行了替代研究, 探索其对大黄鱼幼鱼的生长、体成分、血清指标以及肝脏组织学的影响, 旨在得到玉米蛋白粉替代大黄鱼幼鱼饲料鱼粉的适宜比例, 以期为大黄鱼的饲料应用提供依据。
1 材料与方法
1.1 实验设计与试验饲料
以鱼粉、小麦蛋白粉、玉米蛋白粉(购自宁波天邦股份有限公司)、豆粕为主要蛋白源, 鱼油、大豆油和大豆卵磷脂为主要脂肪源, 配制6种等氮(蛋白水平为45%)等脂(脂肪水平为10%)的实验饲料, 原料组成见表 1。以玉米蛋白粉分别替代0(C0)、15%(C15)、30%(C30)、45%(C45)、60%(C60)和75%(C75)的鱼粉, 各实验组以对照组(C0)饲料赖氨酸、蛋氨酸含量为基准, 分别添加相应水平的晶体赖氨酸和蛋氨酸。具体原料成分及营养成分见表 2、3。各原料经粉碎后过80目筛, 按照配方设计精确称重, 采用逐级扩大方法来将原料混合均匀, 并加水搅拌, 达到充分湿润的效果。采用双螺杆挤条机(华南理工大学, F-26(II)型)制作成条, 然后用制粒机制成粒径分别为2和4 mm的颗粒饲料, 于90℃烘箱熟化30min, 自然风干后保存在–20℃冰箱以备用。
1.2 实验鱼与试验管理
养殖实验在浙江省象山县西沪港区进行。正式实验开始前将大黄鱼放入3 m×6 m×3 m的海水网箱中, 使用商业饲料(粗蛋白45%, 粗脂肪10%)由宁波天邦科技有限公司提供; 暂养2周, 使其适应人工配合饲料。两周后, 挑选体格大小均一且健康无病,平均体质量为(10.49±0.03) g的大黄鱼随机分组。每个处理随机分配三个浮式海水网箱(1.5 m×1.5 m× 2 m), 每个网箱放养60尾鱼, 每天人工饱食投喂两次(05:00—07:00 和 17:00—19:00), 投料速度较为缓慢, 以确保饲料能基本被试验鱼摄入, 尽可能减少沉底浪费, 循环投喂至眼观饱食状态(以大部分鱼不再游到水层表面摄食为准), 并观察有无死鱼,称量计数。养殖周期为8周, 期间水温度为25.5—29.5℃, 盐度为27‰—30‰, 溶解氧含量高于7 mg/L。
1.3 样品采集与分析
样品采集与分析 在养殖试验结束后, 对实验鱼停食1d后, 用丁香酚(1:1000)麻醉, 然后计数,称重。随机取5尾鱼用于常规成分的分析。另外,分别从每个网箱中随机抽取5尾鱼, 用一次性无菌注射器自尾静脉取血, 室温下自然沉降4h, 然后离心(4000 r/min, 15min, 4℃), 制备血清保存于–20℃冰箱, 用于血清指标分析。解剖分离得到肝脏和内脏, 并称量其湿重, 用于肝体比和脏体比, 分离其肝脏用于抗氧化酶活性研究, 另随机取2尾鱼的肝脏在清理完表面的脂肪和结缔组织后放入波恩试液定保存。
饲料原料、饲料以及鱼体常规成分测定采用AOAC(1993)的方法。其中水分测定是用105℃烘箱, 烘至恒重。采用半微量凯氏定氮法(总氮×6.25, VELP, UDK142 automatic distillation unit意大利)测定粗蛋白。索氏抽提法(乙醚为抽提液)测定粗脂肪。而灰分的测定是在马福炉中(550℃)灼烧12h之后称其重量所得。氨基酸的测定送至浙江生态纺织品禁用染化料检测中心有限公司, 采用日立L-8800高速氨基酸分析仪进行分析。
血液生化指标测定及分析 血液生化指标的测定送至宁波大学附属医院进行测, 将所得结果进行分析。
抗氧化酶活性的测定与分析 总抗氧化能力T-AOC、超氧化物歧化酶(SOD)、丙二醛(MDA)、过氧化氢酶(CAT)、谷胱甘肽过氧化物酶(GSHPx)活性的测定均采用南京建成科技有限公司生产的试剂盒进行测定, 测定方法参考说明书进行。
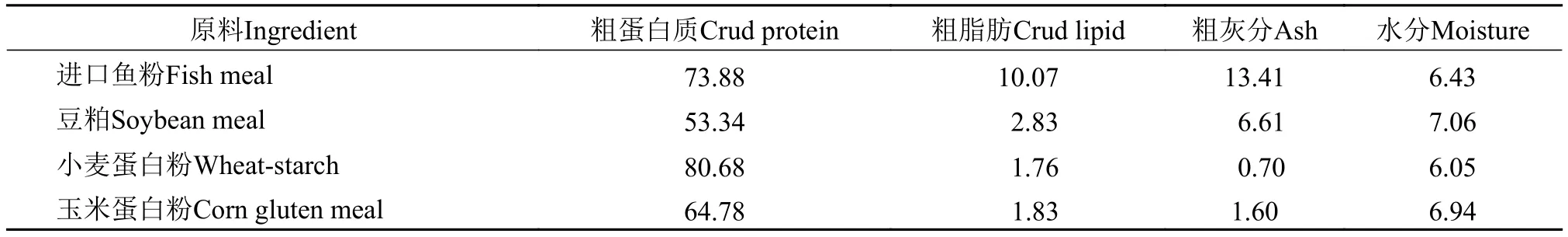
表 1 饲料原料成分(%干物质)Tab. 1 Ingredients of the experimental diets (% dry matter)
肝组织学观察及分析 所有样品在固定36h后, 进行无水乙醇脱水、石蜡包埋、组织切片(厚度为5 μm)、苏木精和伊红染色、中性树胶封片等步骤。使用光学显微镜观察肝脏的组织学特征。
1.4 数据处理及统计分析
特定生长率(Specific growth rate, SGR, %)= 100×(lnWt–lnW0)/t;
存活率(Survival rate, SR) (%)=100×Nt/N0;
饲料系数(Feed conversion ratio, FCR, %)=(T– S)/(Wt–W0);
肝体比(HSI, %)=(Wh/Wt)×100;
脏体比(VSI, %)=(Wv/Wt)×100;
肥满度(Condition factor, CF, %)=100×Wt/L3。
上述公式中W0、Wt分别表示实验鱼初始体重和终末体重; N0、Nt分别表示养殖实验开始时网箱中鱼的尾数和养殖试验结束时网箱中鱼的尾数; T、S分别表示总饲料量和剩余饲料量; Wh为鱼体肝脏质量(g), Wv为鱼体内脏质量(g), t表示实验天数, L是鱼体的体长。
采用SPSS17.0软件对所得数据进行单因素方差分析(ANOVA), 差异显著后进行Tukey多重比较,显著性水平设为P<0.05; 实验结果以平均值±标准差表示。
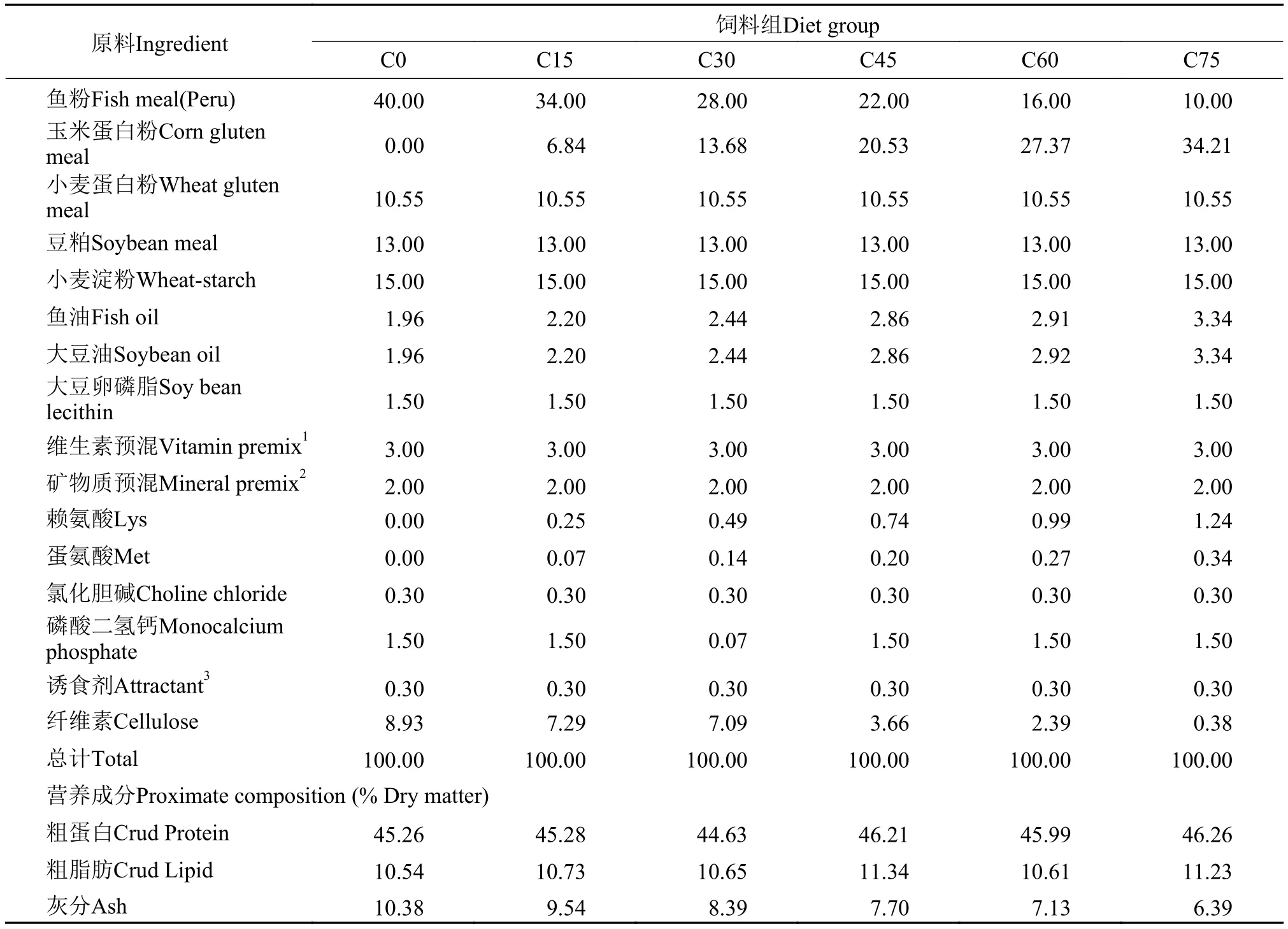
表 2 实验饲料组成及营养成分(%干物质)Tab. 2 Ingredients and nutrients of the experimental diets (% dry matter)
2 结果
2.1 CGM替代鱼粉对大黄鱼幼鱼生长性能和饲料利用的影响
从表 4中可以看出, 在各处理组中, 大黄鱼存活率较高, 均在93.89%—99.44%, 并未受到玉米蛋白粉替代鱼粉水平的显著影响(P>0.05)。各处理组,末体重、特定生长率、增重率、饲料系数等指标均无显差异(P>0.05)。
从表 5中可以看出, 玉米蛋白粉替代鱼粉在C30、C45组大黄鱼幼鱼肥满度显著高于其C0组(P<0.05), 而其他组之间无显著差异(P>0.05)。各处理组之间的肝体比(HSI)、脏体比(VSI)无显著性差异(P>0.05)。
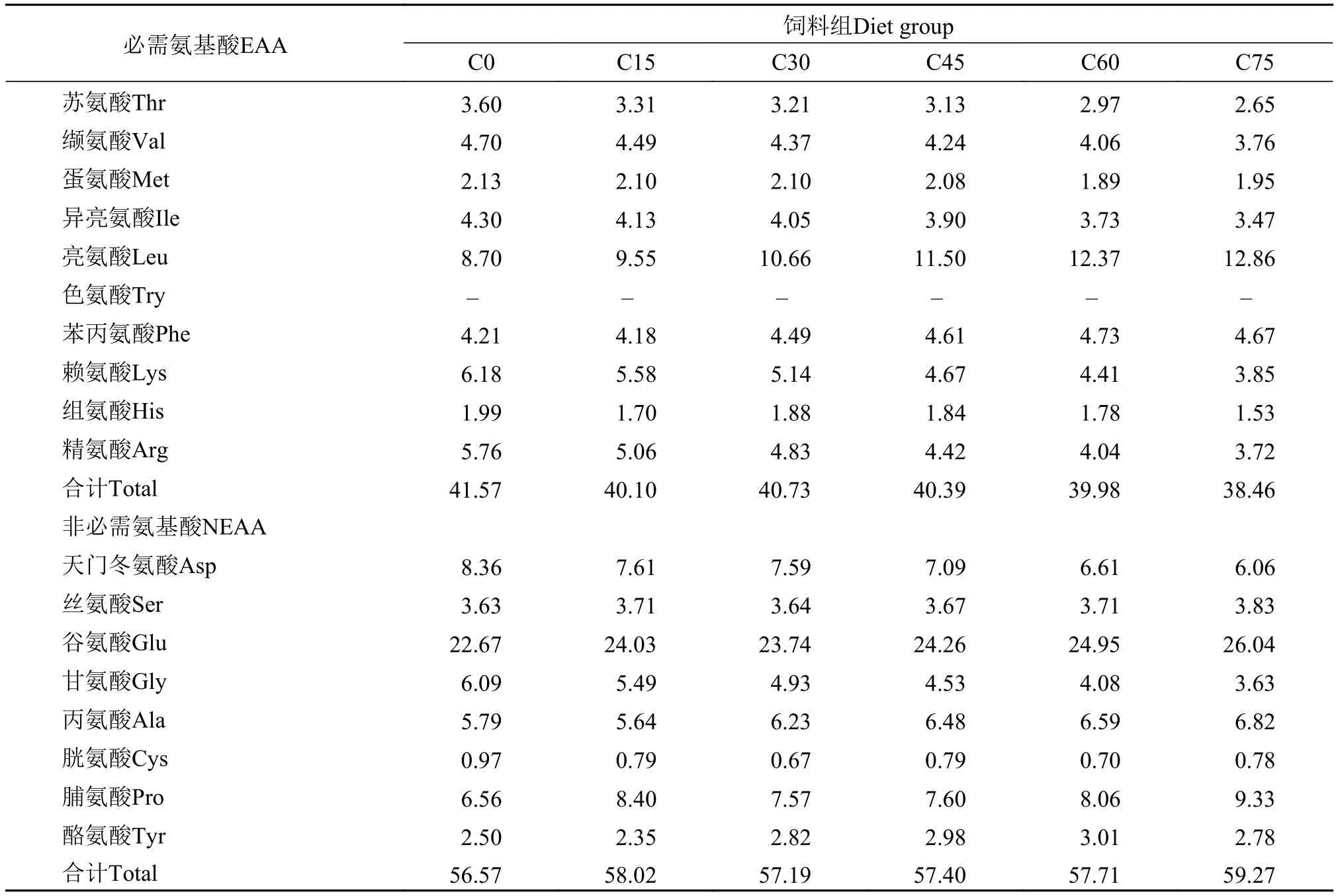
表 3 实验饲料氨基酸组成Tab. 3 The amino acid composition of the experimental diets (% dry matter)
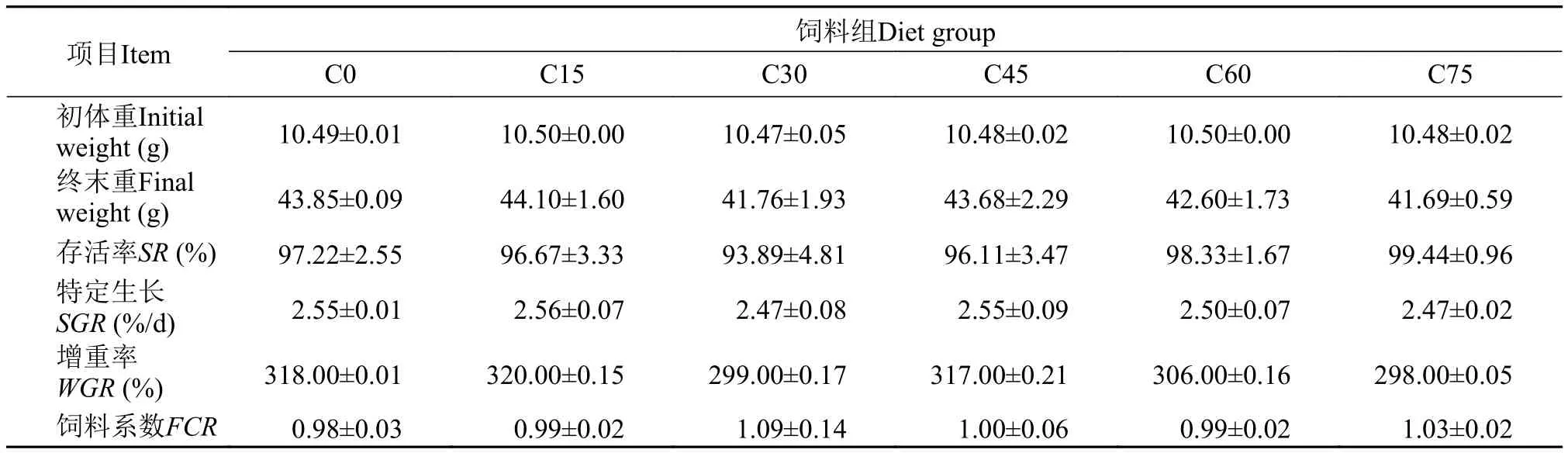
表 4 玉米蛋白粉替代鱼粉对大黄鱼幼鱼生长和饲料系数的影响Tab. 4 Effects of CGM on growth performance and FCR of juvenile large yellow croaker
2.2 CGM替代鱼粉对大黄鱼幼鱼体组成的影响
在实验结束后, 各处理组大黄鱼幼鱼肌肉和全鱼的水分、灰分、粗蛋白及粗脂肪分析数据见表 6。C45和C60组肌肉粗蛋白含量显著高于C0组(P<0.05); 肌肉粗脂肪含量C45组显著高于C0、C15和C75组(P<0.05), 有先升高后降低的趋势; C45、C60和C75组肌肉水分显著低于C0组(P<0.05); 各组肌肉灰分含量无显著性差异(P>0.05)。全鱼粗蛋白、粗脂肪、水分含量无显著差异(P>0.05); 灰分含量有上升趋势, C75组显著高于其他组(P<0.05)。
2.3 CGM替代饲料鱼粉对大黄鱼幼鱼血清指标的影响
从表 7中分析可得, C0组血清总蛋白与球蛋白含量显著高于C30和C75组(P<0.05), 其他组之间无显著差异(P>0.05); 各处理组血清白蛋白含量无显著差异(P>0.05); C75组血清谷丙转氨酶活力显著高于C0、C15和C30组(P<0.05), 其他组之间无显著差异(P>0.05); 各处理组血清谷草转氨酶活力虽无显著差异, 但C45、C60和C75组酶活力明显高于C0组; 血清总胆固醇和甘油三酯的含量随着玉米蛋白粉添加量的增加有下降趋势, C0组总胆固醇含量显著高于C30—C75组(P<0.05), 甘油三酯含量C0组显著高于其他组(P<0.05); 血清葡萄含量各处理组之间无显著性差异(P>0.05)。
2.4 CGM替代饲料鱼粉对大黄鱼幼鱼肝脏组织抗氧化能力的影响
从表 8可以看出, 肝脏超氧化歧化酶(SOD)在C15、C30组与C0有显著差异(P<0.05), 而C45、C60、C75与C0组无显著差异(P>0.05); C75组丙二醛(MDA)显著高于其他组(P<0.05), 其他组之间无显著性差异(P>0.05); 肝脏总抗氧化能力(TAOC)、过氧化氢酶(CAT)、谷胱甘肽过氧化酶(GSH-Px)在各处理组均无显著性差异(P>0.05)。
2.5 CGM替代饲料鱼粉对大黄鱼幼鱼肝脏组织的影响
从肝脏组织图观察显示图 1, C0、C15、C30和C45中实验鱼肝细胞排列整齐, 肝细胞索明显, 细胞较为正常。但随着替代比例的增加, C60和C75组实验鱼肝细胞核偏位, 肝细胞胞浆内有大小不一的脂肪滴, 且脂肪滴渗入细胞胞浆将细胞核挤压到一侧, 甚至肝细胞核心出现了萎缩, 整个肝细胞浆被脂肪滴取代, 使肝细胞肿胀, 胞浆外溢, 使细胞呈现出透明空泡化, 变性的肝细胞随后坏死, 使肝细胞核溶解或消失, 但其结构轮廓仍在。
3 讨论
3.1 CGM替代鱼粉对大黄鱼幼鱼生长性能和体成分的影响
玉米蛋白粉具有几乎不含抗营养因子、来源广、价格低廉等优点[5], 但其在饲料中的添加量因养殖品种而异。在异育银鲫(Carassiusauratus gibelio)的研究中指出, 玉米蛋白粉替代75%的鱼粉不会对其生长产生影响, 但与对照组相比100%替代组生长受到限制、饲料系数升高[12]。在本实验中玉米蛋白粉的替代水平对大黄鱼幼鱼的特定生长率、饲料系数等指标没有显著影响。而在牙鲆(Japanese flounder)的研究中指出, 玉米蛋白粉替代60%的鱼粉其生长指标显著低于对照组[13]。可能是因为添加诱食剂改善了饲料适口性, 没有显著降低大黄鱼幼鱼的摄食量, 从而对生长无显著影响。另外, 也可能是在饲料中添加了适量晶体赖氨酸和蛋氨酸, 晶体氨基酸的添加对植物蛋白源替代水平的效果虽然仍有争议, 但越来越多的研究结果表明,添加晶体氨基酸对提高水产动物对植物蛋白的利用是有作用的[14—16]。因此, 提高植物蛋白源的替代水平可以从适口性、氨基酸平衡等方面进行改善。肥满度和肝体比能够从一定程度上衡量鱼类生理或营养状况。有研究表明: 当植物蛋白源替代鱼粉含量增加后, 鱼体表现出肥满度降低, 肝体比上升的趋势[17]。而在本研究中, 鱼体的肥满度随着替代比例的增加, 有升高再降低的趋势。这种差异可能是因蛋白源替代物、替代比例及鱼的种类等因素有关。在本实验中肥满度在替代水平为45%时达到最高值与C0有显著差异, 表明在鱼体的营养生理层面在C45组能达到大黄鱼幼鱼生长的最适需求。而各组间, 肝体比、脏体比没有显著差异,这与苗星[18]在大黄鱼的饲料中添加不同水平的双低菜粕, 对肝体比、脏体比的影响相似。
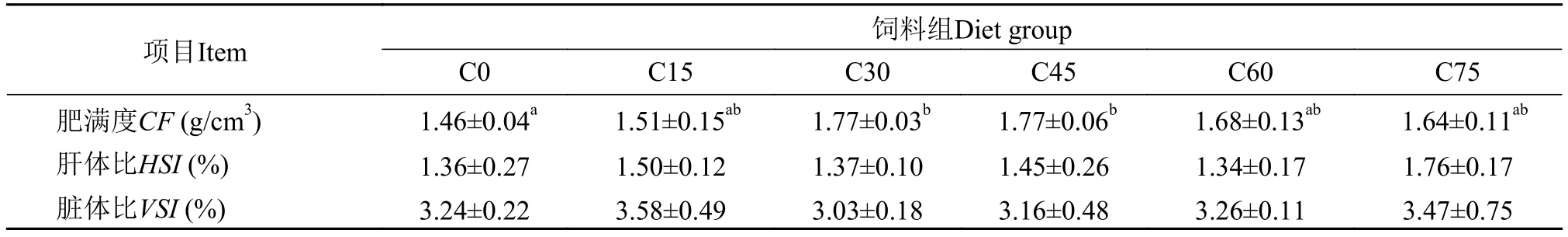
表 5 玉米蛋白粉替代鱼粉对大黄鱼幼鱼肝体指数的影响Tab. 5 Effects of dietary fish meal replacement by CGM on juvenile large yellow croaker somatic parameters (dry weight basis)
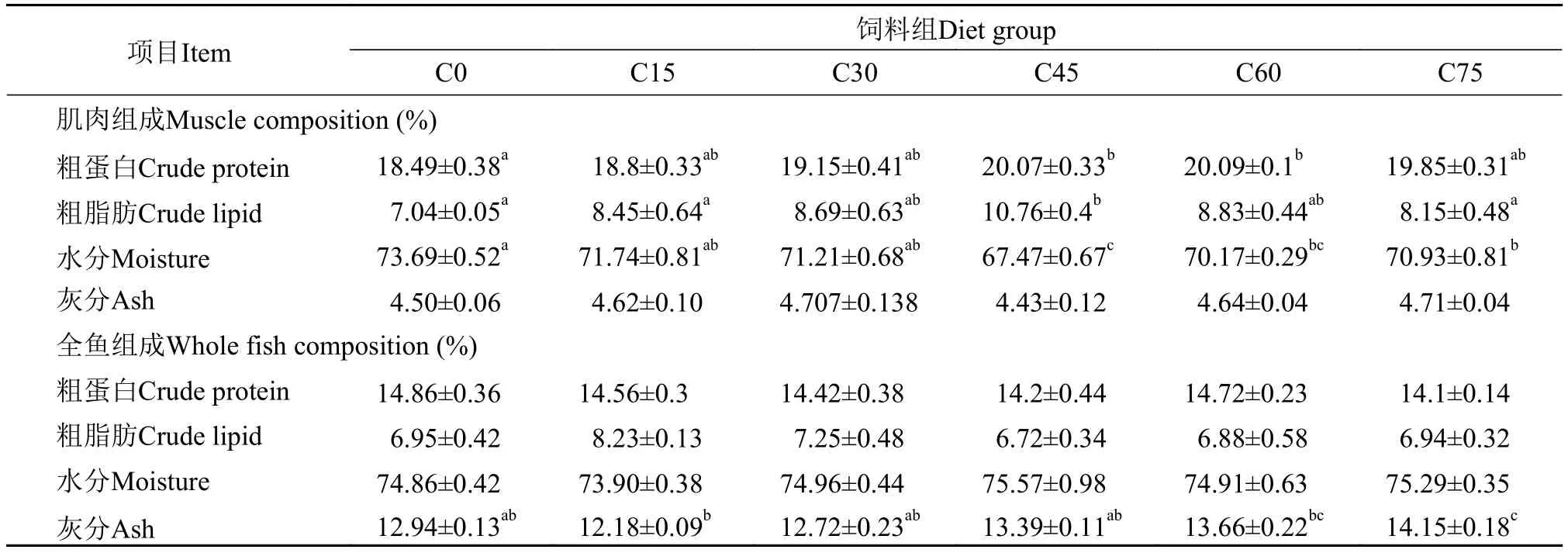
表 6 饲料中玉米蛋白粉替代鱼粉对大黄鱼幼鱼体成分 (湿重)的影响Tab. 6 Effects of dietary fish meal replacement by CGM on body composition (% w d) of juvenile large yellow croaker
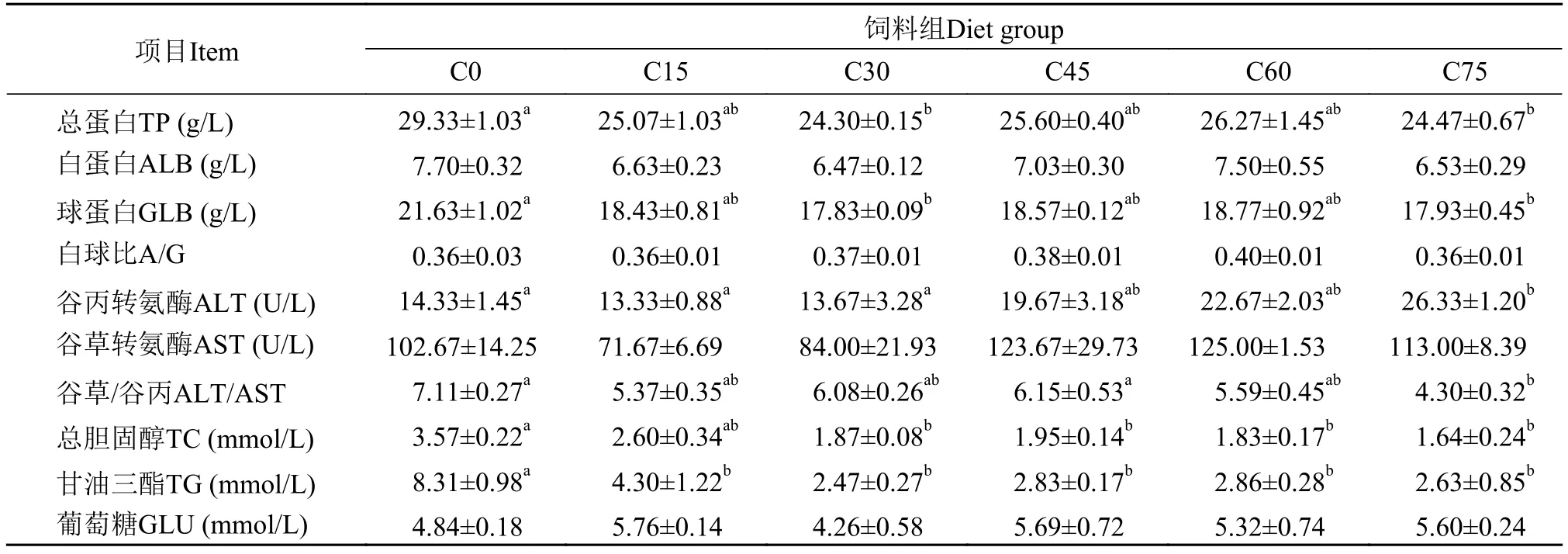
表 7 玉米蛋白粉替代鱼粉对大黄鱼幼鱼血清生化指标的影响Tab. 7 Effects of dietary fish meal replacement by CGM on serum biochem ical indices of juvenile large yellow croaker
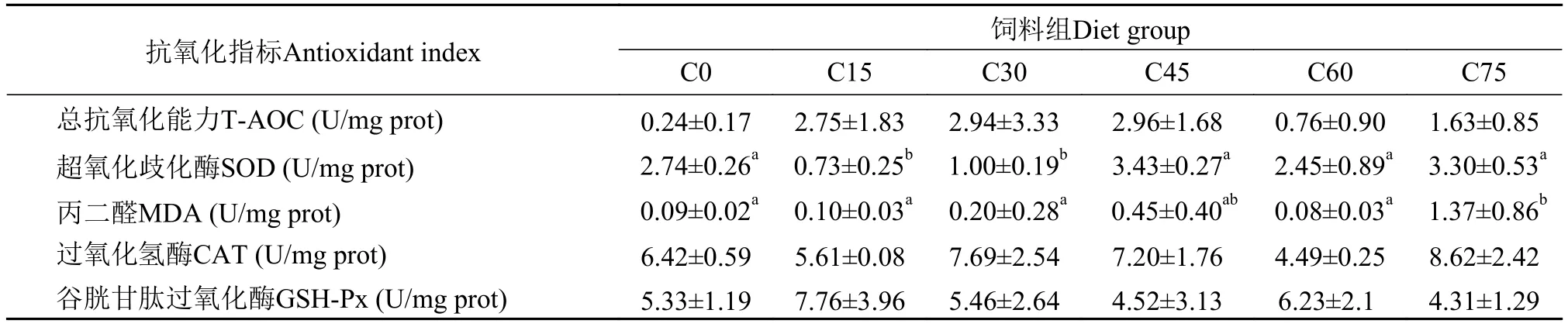
表 8 不同玉米蛋白粉替代鱼粉对大黄鱼肝脏抗氧化指标的影响Tab. 8 The effect of different levels of CGM on indices of antioxidant in juvenile large yellow croaker
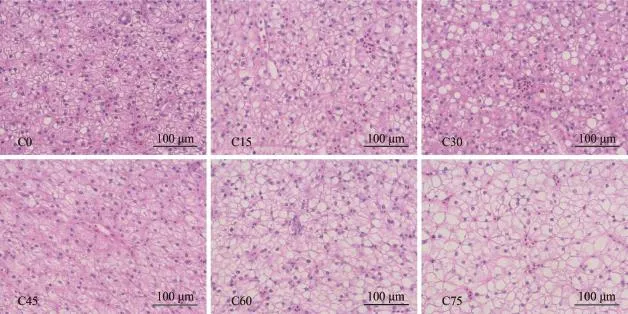
图 1 CGM替代鱼粉对大黄鱼幼鱼肝脏组织的影响(H & E染色, 400×)Fig. 1 Effect of replacement fish meal by CGM on liver histology of large yellow croaker (H & E, 400×)
在本研究中全鱼粗脂肪、粗蛋白各组间无显著性差异, 这与玉米蛋白粉替代鱼粉对大菱鲆[15](S. maximus)、凡纳滨对虾[8](L. vannamei) 的粗脂肪、粗蛋白无显著影响的研究结果一致。程媛嫒等[19]用玉米蛋白粉替代鱼粉对罗氏沼虾的研究结果表明玉米蛋白粉对罗氏沼虾体组成和肌肉中的粗蛋白、粗脂肪和粗灰分都没有显著影响, 而在本试验中肌肉粗蛋白在C45和C60组含量显著高于C0组,而全鱼粗蛋白含量各组间无显著差异, 这说明肌肉组织对饲料中的营养形式变化较为敏感, 反映了机体组织对营养调控的差异性[20]。肌肉粗蛋白升高可能是因为替代组添加的晶体氨基酸可以促进大黄鱼幼鱼饲料中必须氨基酸的平衡性, 减少氨基酸用于分解供能, 从而蛋白质合成量增加。在试验中用玉米蛋白粉替代鱼粉后全鱼粗脂肪含量与对照组没有显著差异, 其原因之一可能是玉米蛋白中不可消化的碳水化合物含量比较少, 二是因为在试验中添加了赖氨酸, 所以饲料中不缺乏生酮氨基酸。大黄鱼幼鱼全鱼灰分随着替代比例的升高而升高,该结果与圆斑星鲽[21](Verasper variegatus)研究结果相一致。
3.2 CGM替代饲料鱼粉对大黄鱼幼鱼血清指标及幼鱼肝脏的影响
CGM替代饲料鱼粉对大黄鱼幼鱼血清指标的影响 肝脏是蛋白质合成的主要场所, 血清总蛋白含量可以反映出肝脏对蛋白质的合成能力。对哲罗鱼(Hucho taimen)的研究中发现, 随着大豆分离蛋白替代饲料鱼粉水平的增加, 其血清中总蛋白含量降低[22]。而用蚕豆蛋白的研究发现, 白鲟(Huso huso)血清总蛋白含量并没有随着替代比例增加而表现出显著性差异[23]。在本研究中, 对照组总蛋白含量最高, C75组显著低于对照组, 这表明肝脏对鱼粉蛋白的吸收优于玉米蛋白, 过高的玉米蛋白添加影响了肝脏对蛋白质的合成能力。在正常情况下谷丙转氨酶(ALT)主要存在于肝脏细胞中,而谷草转氨酶(AST)主要存在于肝细胞线粒体, 它们在血清中的含量很低, 只有当细胞膜通透性增强或细胞坏死时才会大量进入血液, 使血清酶活增加[24]。因此, 血清ALT和AST可作为反映肝脏功能的重要指标。本研究发现, 血清ALT含量随着替代比例增加而升高, C60和C75组血清ALT含量显著高于对照组, 表明玉米蛋白粉替代达60%以上的鱼粉时对肝脏细胞造成了一定程度的损伤, 这与肝组织切片结果一致。植物蛋白源降低胆固醇的作用在很多研究中已经证实[13,25]。本研究也得到相似结果: 随着玉米蛋白粉的添加水平的升高, 血清总胆固醇含量呈下降趋势。可能原因是胆固醇在胃肠中的吸收受阻, 或是由于植物蛋白源替代中胆固醇不足等因素导致其下降[26]。
CGM替代饲料鱼粉对大黄鱼幼鱼肝脏抗氧化能力的影响 在正常生理条件下, 活性氧(自由基)类高活性分子的生成和清除维持在稳态水平和平衡状态。当活性氧的生成超过机体的清除能力时, 就会使氧化还原平衡紊乱, 导致氧化应激的发生, 造成多种大分子(DNA、蛋白质、脂质)的损伤,进而导致多种疾病的发生。抗氧化酶, 对于抵抗氧化损伤, 维持氧化还原平衡具有重要作用[27]。机体的抗氧化能力还是评价水生动物健康的重要指标之一。总抗氧化能力(T-AOC)是机体抗氧化系统功能的综合体现, 可以反映出机体的抗氧化应激能力[28]; 超氧化物歧化酶(SOD)是抗氧化体系中首先被激活的酶类, 具有清除自由基的作用[29]; 过氧化氢酶(CAT)是动物体内的重要抗氧化酶, 它能将H2O2进一步转化成水[30], CAT活力的高低表现为H2O2浓度的增加, H2O2的浓度可能与SOD的应激反应有关[27]; 谷胱甘肽过氧化酶(GSH-Px)的功能与CAT相似, 对H2O2具有极强的清除作用, 对于保护生物大分子和细胞膜结构不受氧化物质的损伤具有重要作用[24]。丙二醛(MDA)是脂质过氧化的产物, 可以作为判断机体细胞受氧自由基损伤程度的指标[29]。实验结果表明: 各处理组T-AOC、GSHPx和CAT没有显著性差异, 但替代组T-AOC的含量均高于对照组, 说明饲料中玉米蛋白粉的添加对大黄鱼幼鱼的抗氧化能力有一定的促进作用。C75组与C0组MAD有显著差异且是最大值, 说明替代比例超过60%时, 大黄鱼幼鱼处于氧化应激状态, 这与曹俊明等[31]用蝇蛆粉替代鱼粉水平超过60%时,凡宾纳对虾处于氧化应激状态的研究结果一致。其出现氧化应激的原因可能是与饲料中脂肪含量过多有关, 王朝明等[32]等研究表明饲料中脂肪含量过多从某种程度上会使机体清除自由基的能力减弱, 从而使机体MDA的含量增加。
CGM替代饲料鱼粉对大黄鱼幼鱼肝脏肝脏组织的影响 肝脏组织学观察, 从组织学的角度对养殖鱼类的摄食营养进行组织学分析, 是评价鱼体对植物蛋白源吸收利用率的方法之一。由于养殖鱼类食用人工配合饲料, 轻度脂肪堆积在养殖鱼类中十分常见[33]。在Couto等[34]对白姑鱼(Argyrosomus regius)的研究指出, 饲料中添加225 g/kg角斗胚芽粕不会引起肝组织病变。但一些研究发现, 角斗胚芽粕添加的增加, 真鲷(Sparus aurata)肝细胞空泡化加剧[35]; 在混合蛋白替代鱼粉的研究中也发现, 高比例的混合蛋白添加会导致脂肪变性[36]。本替代组织学观察表明, 玉米蛋白粉添加从0—45%,肝脏脂肪堆积不明显, 进一步添加时脂肪堆积加重,高替代比例75%添加组观察到严重脂肪变性, 其可能的原因是随着替代比例的增加, 饲料脂肪的含量也随着增加, 使鱼体能量过剩, 肝细胞中脂肪沉积,肝的生长机能降低, 是脂蛋白合成减少, 使肝中的脂肪不能及时运出, 从而导致肝组织损伤[37]。这与脂质抗氧化指标MAD检测结果相吻合, 其MAD含量在替代比例为75%时较对照组显著升高。其原因是高替代比例试验组, 自由基对肝脏中的多不饱和脂肪酸具有很高的亲和力, 引发脂质氧化, 形成不稳定的氢过氧化物, 并迅速降解成MDA等物质,促使MDA含量升高, 如此过量的抗氧化剂伴随着自由基对肝脏的损伤会加重。此外, 谷草转氨酶(AST)和谷丙转氨酶(ALT)在机体蛋白质代谢中起重要作用, 其活性变化亦是反映肝细胞受损伤的主要敏感指标[24]。在健康动物血清中转氨酶的活力很小。因为脊椎动物在正常情况下肝脏内的转氨酶只有少量被释放到血液中, 因此血清中的转氨酶活性较小。而在本实验中, 随着替代比例的增加,血清ALT呈上升趋势, 且替代比例达到75%时显著高于对照组, 此时细胞膜的通透性加大, 大量ALT渗入血液中, 表明肝组织受到损伤, 这与肝组织切片观察结果相吻合。
4 结论
综合大黄鱼幼鱼的生长、体成分、血清生化指标及肝组织学结果, 认为在本实验条件下, 建议玉米蛋白粉添加量为45%。
[1]Ai C X, Tao C Y. Fish meal replacement-expensive fish meal is run with aquatic feed research and development of technical countermeasures [J]. Feed Industry Magazine, 2013, 34(10): 1—7 [艾春香, 陶春燕. 鱼粉替代-鱼粉高价运行下水产配合是饲料研发的技术对策. 饲料工业, 2013, 34(10): 1—7]
[2]Zhou Q C, Mai K S, Liu Y J, et al. Advances in animal and plant protein sources in place of fish meal [J]. Journal of Fisheries of China, 2005, 26(3): 404—410 [周歧存, 麦康森, 刘永坚, 等. 动植物蛋白源替代鱼粉研究进展. 水产学报, 2005, 26(3): 404—410]
[3]Yao J P, Xiao Q. Advances in the study of alternative protein sources in place of fish meal in aquaculture [J]. Feed Research, 2015, 24: 39—46 [姚俊鹏, 肖勤. 水产养殖中鱼粉替代蛋白源的研究进展. 饲料研究, 2015, 24: 39—46]
[4]Gatlin D M, Barrows F T, Brown P, et al. Expanding the utilization of sustainable plant products in aquafeeds: a review [J]. Aquaculture Research, 2007, 38(6): 551—579
[5]Regoast C, Arzel J, Kaushik S J. Partial or total replacement of fish meal by corn gluten meal in diet for turbot (Psetta maxima) [J]. Aquaculture, 1999, 180(1—2): 99—117
[6]Luo Y W, Ai Q H, Mai K S, et al. Effects of dietary corn gluten meal on growth performance and protein metabolism in relation to IGF-I and TOR gene expression of juvenile cobia (Rachycentron canadum) [J]. Journal of Ocean University of China, 2013, 12(3): 418—426
[7]Fournier V, Huelvan C, Desbruyeres E. Incorporation of a mixture of plant feedstuffs as substitute for fish meal in diets of juvenile turbot (Psetta maxima) [J]. Aquaculture, 2004, 236(1): 451—465
[8]Han B, Huang X X, Hua X M, et al. Effects of partial replacement of fish meal by corn gluten meal on daily ration growth and nutrient ingredients in muscles of Litopenaeus vannamei [J]. Journal of Fisheries of China, 2009, 33(4): 658—665 [韩斌, 黄旭雄, 华雪铭, 等. 玉米蛋白粉替代部分鱼粉对凡纳滨对虾摄食量、生长和肌肉成分的影响. 水产学报, 2009, 33(4): 658—665]
[9]Ai Q H, Mai K S, Tan B P, et al. Replacement of fish meal by meat and bone meal in diets for large yellow croaker, Pseudosciaena crocea R. [J]. Aquaculture, 2006, 260(1—4): 255—263
[10]Zhang L, Mai K S, Ai Q H, et al. Use of a compound protein source as a replacement for fish meal in diets of large yellow croaker, Pseudosciaena crocea R. [J]. Aquaculture, 2008, 39(1): 83—90
[11]Li J, Zhang L, Mai K S, et al. Potential of several protein sources as fish meal substitutes in diets for large yellow croaker, Pseudosciaena crocea R. [J]. Aquaculture, 2010, 39(1): 278—283
[12]Chen R, Hua X M, Zhou H Q, et al. Effects of replacement of fish meal by corn gluten meal in diets on growth, protease activities and apparent digestibilities of allogynogenetic silver crucian carp [J]. Journal of Shanghai Jiaotong University (Agricultural Science), 2009, 27(4): 358—367 [陈然, 华雪铭, 黄旭熊, 等. 玉米蛋白粉替代鱼粉对异育银鲫生长、蛋白酶活性及表观消化率的影响. 上海交通大学学报 (农业科学版), 2009, 27(4): 358—367]
[13]Kikuchi K. Partial replacement of fish meal with corn gluten meal in diets for Japanese flounder Paralichthys olivaceus [J]. Journal of the World Aquaculture Society, 1999, 30(3): 357—363
[14]Kaushik S J, Cravedi J P, Lalles J P, et al. Partial or total replacement of fish meal by soybean protein on growth, protein utilization, potential estrogenic or antigenic effects, choleste- rolemia and flesh quality in rainbow trout, Oncorhynchus mykiss [J]. Aquaculture, 1995, 133(3): 257—274
[15]Liu X W, Mai K S, Ai Q H, et al. Replacement of fish meal by corn gluten meal in diets of Scophthatmus maximus [J]. Journal of Fisheries of China, 2012, 36(34): 66—472 [刘兴旺, 麦康森, 艾庆辉, 等. 玉米蛋白粉替代鱼粉对大菱鲆摄食、生长及体组成的影响. 水产学报, 2012, 36(34): 466—472]
[16]Zhang J L, Yu P, Huang J Q, et al. Effects of fish meal replacement by defatted silkworm pupae on growth performance body composition and health status of jian carp Cyprinus carpio var. Jian [J]. Chinese Journal of Animal Nutrition, 2013, 25(7): 1568—1578 [张建禄, 余平, 黄吉芹, 等. 脱脂蚕蛹替代饲料中鱼粉对建鲤生长性能、体成分及健康状况的影响. 动物营养学报, 2013, 25(7): 1568—1578]
[17]Yue Y R, Zhou Q C. Effect of replacing soybean meal with cottonseed meal on growth, feed utilization, and hematological indexes for juvenile hybrid tilapia, Oreochromis niloticus×O. aureus [J]. Aquaculture, 2008, 284(1): 185—189
[18]Miao X. Studies on the tolerance of large yellow croaker ton dietary soybean meal [D]. Thesis for Master of Science. Fisheries College, Ocean university of China. Qingdao. 2014 [苗星. 大黄鱼对豆粕和双低菜粕的耐受性研究. 硕士学位论文, 中国海洋大学水产学院, 青岛. 2014]
[19]Cheng Y Y, Zhou H Q, Hua X M, et al. Effects of partial replacement of dietary fish meal by corn gluten meal on growth, nutrient ingredients and amino acid retention in muscle of Macrobrachium rosenbergii [J]. Journal of Fishery Sciences of China, 2009, 16(4), 572—579 [程媛媛, 周洪琪, 华雪铭, 等. 玉米蛋白粉部分替代鱼粉对罗氏沼虾生长, 氨基酸沉积率和肌肉营养成分的影响. 中国水产科学, 2009, 16(4): 572—579]
[20]Kader M A, Koshio S, Ishikawa M, et al. Growth, nutrientutilization, oxidative condition, and element composition of juvenile red sea bream Pagrus major fed with fermented soy-bean meal and scallop by-product blend as fishmeal replacement [J]. Fisheries Science, 2011, 77(1): 119—128
[21]Lu Y Y, Chang Q, Chen S Q, et al. The effects of dietary fermented soybean meal on the growth and digestive ability of spotted halibut, Verasper variegatus [J]. Acta Hydrobiologica Sinica, 2016, 40(1): 10—18 [吕云云, 常青,陈四清, 等. 发酵豆粕对圆斑星鲽生长及消化能力的影响. 水生生物学报, 2016, 40(1): 10—18]
[22]Xue Q Y, Wang C A, Xu H, et al. Effects of replacing fish meal with soy protein isolated on the growth performance body composition and biochemical indexies of juvenile hucho taimen [J]. Acta Hydrobiologica Sinica, 2008, 32(6): 941—946 [徐奇友, 王常安, 许红, 等. 大豆分离蛋白替代鱼粉对哲罗鱼稚鱼生长、体成分和血液生化指标的影响. 水生生物学报, 2008, 32(6): 941—946]
[23]Soltanzadeh S, Fereidouni A E, Ouraji H, et al. Growth performance, body composition, hematological, and serum biochemical responses of beluga (Huso huso) juveniles to different dietary inclusion levels of faba bean (Vicia faba) meal [J]. Aquaculture International, 2016, 24(1): 395—413
[24]Deng J M, Mai K S, Chen L Q, et al. Effects of replacing soybean meal with rubber seed meal on growth, antioxidant capacity, non-specific immune response, and resistance to Aeromonas hydrophila in tilapia (Oreochromis niloticus×O. aureus) [J]. Fish & Shellfish Immunology, 2015, 44(2): 436—444
[25]Dai W W, Mai K S, Xu W, et al. Effects of replacing fish meal with plant-based protein on growth, physiological and biological indices, and intestinal histology in tongue sole, Cynoglossus semilaevis Güntuer [J]. Journal of Fishery Sciences of China, 2016, 23(1): 125—137 [代伟伟, 麦康森, 徐玮, 等. 复合植物蛋白源替代鱼粉对半滑舌鳎生长、生理生化指标和肠组织结构的影响. 中国水产科学, 2016, 23(1): 125—137]
[26]Yun B, Ai Q H, Mai K S, et al. Synergistic effects of dietary cholesterol and taurine on growth performance and cholesterol metabolism in juvenile turbot (Scophthalmus maximus L.) fed high plant protein diets [J]. Aquaculture, 2012, 324—325: 85—91
[27]Sun Q G, Long Z, Zhang X D, et al. Novel progress in antioxidant system [J]. Progress in Modern Biomedicine, 2016, 16(20): 2197—2200 [孙全贵, 龙子, 张晓迪, 等. 抗氧化系统研究新进展. 现代生物医学进展, 2016, 16(20): 2197—2200]
[28]Wang J Y, Jiang K J, Xia B, et al. Effects of small dietary peptides on digestive enzyme activity, antioxidative capability, and biochemical composition in tissues of juvenile starry flounder Platichthys stellatus [J]. Journal of Fishery Sciences of China, 2014, 21(6): 1154—1164 [王际英, 姜柯君, 夏斌, 等. 小肽对星斑川鲽幼鱼消化酶活性、抗氧化能力和生化组成的影响. 中国水产科学, 2014, 21(6): 1154—1164]
[29]Sun L M, Chen L Q, Li E C, et al. Effects of dietary methionine supplementation on feeding, growth and antioxidant ability of juvenile Chinese mitten crab, Eriocheir sinensis [J]. Acta Hydrobiologica Sinica, 2013, 37(2): 336—343 [孙立梅, 陈立侨, 李二超, 等. 高比例棉粕饲料中补充蛋氨酸对中华绒螯蟹幼蟹摄食、生长及抗氧化酶活性的影响. 水生生物学报, 2013, 37(2): 336—343]
[30]Gao C S, Wang C X, Zhang S S. Effects of copper on activities of antioxidant enzymes and total antioxidative competence in hepatopancreas of Cyprinus Carpio [J]. Journal of Agro-Environment Science, 2008, 27(3): 1157—1162 [高春生, 王春秀, 张书松. 水体铜对黄河鲤肝胰脏抗氧化酶活性和总抗氧化能力的影响. 农业环境科学学报, 2008, 27(3): 1157—1162]
[31]Cao J M, Yan J, Huang Y H. Effects of replacement of fish meal with housefly maggot meal on growth performance, antioxidant and nonspecific immune indexes of juvenile Litopenaeus vannamei [J]. Journal of Fisheries of China, 2012, 36(4): 529—537 [曹俊明, 严晶, 黄燕华,等, 家蝇蛆粉替代鱼粉对凡纳滨对虾生长、抗氧化和免疫指标的影响. 水产学报, 2012, 36(4): 529—537]
[32]Wang C M, Luo L, Zhang G Z. Effect of dietary lipid level on growth performance, body composition and antioxidant capacity of juvenile Chinese sucker (Myxocyprinus asiaticus) [J]. Freshwater Fisheries, 2010, 40(5): 47—53 [王朝明, 罗莉, 张桂众, 等. 饲料脂肪水平对胭脂鱼幼鱼生长、体组成和抗氧化能力的影响. 淡水渔业, 2010, 40(5): 47—53]
[33]Spisni E, Tugnoli M, Ponticelli A, et al. Hepatic steatosis in artificially fed marine teleosts [J]. Journal of Fish Diseases, 1998, 21(3): 177—184
[34]Couto A, Barroso C, Guerreiro I, et al. Carob seed germ meal in diets for meagre (Argyrosomus regius) juveniles: Growth, digestive enzymes, intermediary metabolism, liver and gut histology [J]. Aquaculture, 2016, 451: 396—404
[35]Silvia M L, Rosa B A, Silvia N M, et al. Carob seed germ meal as a partial substitute in gilthead sea bream (Sparus aurata) diets: Amino acid retention, digestibility, gut and liver histology [J]. Aquaculture, 2012, 338—341: 124—133
[36]Hu L, Yun B, Xue M, et al. Effects of fish meal quality and fish meal substitution by animal protein blend on growth performance, flesh quality and liver histology of Japanese seabass (Lateolabrax japonicus) [J]. Aquaculture, 2013, 372—375: 52—61
[37]Shi X, Luo Z, Huang C, et al. Effect of substituting chlorella sp. for regular fishmeal on growth, body composition, hepatic lipid metabolism and histology in crucian carp Carassius auratus [J]. Acta Hydrobiologica Sinica, 2015, 39(3): 498—506 [石西, 罗智, 黄超, 等. 小球藻替代鱼粉对鲫生长、体组成、肝脏脂肪代谢及其组织学的影响. 水生生物学报, 2015, 39(3): 498—506]
EFFECTS OF REPLACING FISH MEAL WITH CORN GLUTEN MEAL ON THE GROWTH, SERUM BIOCHEMICAL INDICES AND LIVER HISTOLOGY OF LARGE YELLOW CROAKER LARIMICHTHYS CROCEA
HE Jiao-Jiao1, WANG Ping1,2, FENG Jian1, LOU Yu-Dong1, DANG Hui1and DENG Rong1
(1. Zhejiang Province Key Laboratory of Marine Aquaculture Equipment and Engineering Technology, Zhejiang Ocean University, Zhoushan 316000, China; 2. School of Marine Sciences, Ningbo University, Ningbo 315211, China)
To investigate the effects of replacing fish meal by corn gluten meal (CGM) on fish growth, serum biochemical indices and liver histology, six isonitrogenous (45% crude protein) and isolipidic (10% crude lipid) diets with various dietary protein levels (0, 15%, 30%, 45%, 60% and 75%) (designated as C0, C15, C30, C45, C60, C75) were formulated to feed triplicate groups of juvenile large yellow croaker [mean initial weight: (10.49±0.03) g] for a 56 days trail. In addition, all diets except the control diet were supplemented with crystalline amino acids such as lysine and methionine. Results showed that the survival rate (SR), specific growth rate (SGR) and feed conversion ratio (FCR) were not significantly affected (P>0.05). Muscle crude protein content in C45 and C60 were significantly higher than that of C0 (P<0.05). Muscle crude lipid content in C45 was significantly higher than that of C0, C15 and C75 (P<0.05). Muscle moisture in C45, C60 and C75 were significantly lower than that of C0. CGM had no effects on crude protein, crude lipid and moisture of whole body (P>0.05), while, the increased CGM replacement enhanced the whole body ash. CGM did not impact serum ALB, A/G, AST and GLU. The increasing dietary CGM level decreased total TC with a significant difference when the substitution level was more than 30% (P<0.05). The serum TP and GLB content of C0 was significantly higher than that of C75 (P<0.05). Serum AST of C60 and C75 were significantly higher than that of C0 (P<0.05) with the highest in the C75. No significant differences were observed in the T-AOC, CAT and GHS-Px by CGM (P>0.05). While CGM can increase the T-AOC content. Compared to C0 diet, C75 had significant and highest value in MDA (P<0.05). From the liver histology, replacement proportion of fish meal by CGM more than 45% mediated liver cell nucleus location and increased lipids accumulation that were translucent with vacuole. These results suggest that the optimal dietary protein level in juvenile large yellow croaker is about 45%.
Large yellow croaker; Corn gluten meal; Growth performance; Serum biochemical indices; Antioxidase; Liver histology
S965.3
A
1000-3207(2017)03-0506-10
10.7541/2017.65
2016-06-22;
2016-11-10
浙江省自然科学重点基金(Z16E090006); 国家海洋公益性行业科研专项(201505025); 舟山市海洋专项(2015C41001); 浙江省重中之重学科开放基金(XKZSC1406); 国家自然科学基金(31602205); 浙江海洋大学省重中之重学科项目(20161105)资助[Supported by the National Natural Science Foundation of Zhejiang Province (Z16E090006); Public Science and Technology Research Funds Projects of Ocean (201505025); the Special Funds Projects of Ocean in Zhoushan, Zhejiang Province (2015C41001); the Key Discipline Open Fund of Zhejiang Province (XKZSC1406); the National Science Foundation of China (31602205); the Key Discipline Projects for Zhejiang Ocean University of Zhejiang Province (20161105)]
何娇娇(1990—), 女, 贵州省湄潭县人; 硕士研究生; 研究方向为海洋生物养殖学。E-mail: 915316064@qq.com
王萍, 副教授, 硕士研究生导师; E-mail: wp77319@163.com
猜你喜欢
当代水产(2022年8期)2022-09-20
当代水产(2022年6期)2022-06-29
农村百事通(2021年31期)2021-12-13
中老年保健(2021年3期)2021-08-22
农村百事通(2021年11期)2021-01-17
家庭医药(2020年1期)2020-02-10
当代水产(2019年6期)2019-07-25
当代水产(2019年6期)2019-07-25
当代水产(2019年1期)2019-05-16
农产品市场周刊(2017年45期)2017-12-28
