
早胜牛遗传多态性与母系遗传背景分析
2017-07-18 12:10容维中徐建峰郭海龙保国俊桑国俊
中国牛业科学 2017年2期
容维中, 徐建峰, 王 珂, 郭海龙, 保国俊, 桑国俊, 杨 明
(甘肃省畜牧兽医研究所,甘肃 平凉 744000)
早胜牛遗传多态性与母系遗传背景分析
容维中, 徐建峰*, 王 珂, 郭海龙, 保国俊, 桑国俊, 杨 明
(甘肃省畜牧兽医研究所,甘肃 平凉 744000)
[目的] 为评价和挖掘早胜牛遗传资源及种质资源,开展了早胜牛母系遗传背景及分子遗传特性研究。[方法] 以早胜牛mtDNA D-Loop区为标记位点,对基因组DNA进行了PCR扩增和测序;以8个中国地方黄牛品种和5个引进品种的mtDNA D-Loop区核苷酸序列为对照,分析了核苷酸多态位点、核苷酸多样性、单倍型数等遗传多态性指标,构建了系统发育树。[结果] 早胜牛及其杂交类群mtDNA D-Loop区富含A和T,检测到80个核苷酸变异位点,存在转换、颠换、转换和颠换共存3种突变类型,且以转换为主;界定了59个单倍型,其中8个共享单倍型,51个特有单倍型;我国地方黄牛分为两大支系,多数与非洲瘤牛、欧洲普通牛聚为一类,少数与印度瘤牛聚为一类;与其他地方黄牛品种相比,早胜牛及其杂交类群与秦川牛的亲缘关系最近。[结论] 早胜牛及其杂交类群mtDNA D-Loop区核苷酸变异丰富,群体变异程度较高,与秦川牛的亲缘关系最近。
早胜牛;线粒体D-loop;系统发育;亲缘关系
早胜牛作为秦川牛的优秀地方类群之一[1],因其体格高大、体躯紧凑、肌肉丰满等特性,已成为甘肃省陇东地区养殖的主导肉牛群体。然而由于近年来盲目规模引种,造成牛种混杂,生产性能不稳定或下降,尤其是经过多年积淀形成的固有特性及优势基因逐渐流失,对地方种质资源的保护和利用形成了较大威胁。
线粒体DNA(mtDNA)是动物核外唯一的遗传物质,具有结构简单稳定、进化速度快、无组织特异性等特点,核苷酸变异丰富,被广泛应用于动物的系统发育、演化及遗传多样性研究中[2~3]。为了客观评价早胜牛的母系遗传背景及分子遗传特性,本研究以mtDNA D-loop区为分子标记位点,对早胜牛及其杂交类群的遗传多样性性及群体结构进行了研究,分析了与8个中国地方黄牛品种和5个引进品种的亲缘关系,旨在理清早胜牛的遗传背景及起源,为早胜牛遗传资源保护和种质资源创新利用提供理论依据。
1.材料与方法
1.1 血样采集
在庆阳市早胜塬等主产区采集早胜牛及其杂交类群血样86头,其中:早胜牛52头(简称ZS),南德温×早胜牛杂交一代7头(简称NZ),安格斯×早胜牛杂交一代27头(简称AZ),采集血样7ml,低温条件下带回实验室,于-20℃超低温冰箱保存。
1.2 基因组DNA提取
采用Omegabiotek公司生产的D3471型血液DNA提取试剂盒提取基因组DNA,用1%浓度的琼脂糖凝胶电泳检测DNA纯度和分光光度计检测OD260/OD280值。
1.3 引物设计与PCR扩增
以黄牛mtDNA D-Loop区为标记,参照已发表在GenBank中的黄牛核苷酸序列(Accession NO.NC_006853)设计引物[4],送至北京三博远志生物技术有限责任公司进行合成。引物序列:上游: 5′-CTGCAGTCTCACCATCAACC-3′,下游:5′-GGGGTGTAGATGCTTGC-3′。
PCR反应体系:总体积为50 μL,其中2×Taq Master Mix 混合液25 μL,上下游引物(10 pmol/μL)各2 μL,DNA模板(100 ng/μL)2μL,ddH2O 19 μL。反应条件:预变性94℃/4min,31个循环(变性94℃/1min,退火58℃/1min,延伸72℃/1min),延伸72℃/10min,4 ℃保存。
1.4 基因测序
对经1%浓度琼脂糖凝胶电泳检测合格的PCR产物,送至北京三博远志生物技术有限责任公司进行纯化和测序。
1.5 数据处理与软件分析
对测序获得的早胜牛及其杂交类群mtDNA D-Loop区核苷酸序列,采用EeitSeq软件进行人工编辑,并以秦川牛、晋南牛、鲁西黄牛、南阳牛、延边牛、西镇牛、蒙古牛、哈萨克牛以及野牦牛、欧洲普通牛、印度瘤牛、非洲瘤牛的mtDNA D-Loop区核苷酸序列为对照,通过 SeqMan软件进行同源性比对分析,采用MAGE5.0软件分析核苷酸多态位点、核苷酸多样性、单倍型数等遗传多态性指标,通过MAGE5.0及DnaSP 5.10软件构建系统发育树。
2 结果
2.1 PCR扩增结果
对早胜牛及其杂交类群D-Loop区基因组DNA进行扩增,由图1可见,PCR扩增产物条带清晰,特异性较好,且与目的片段大小基本一致。
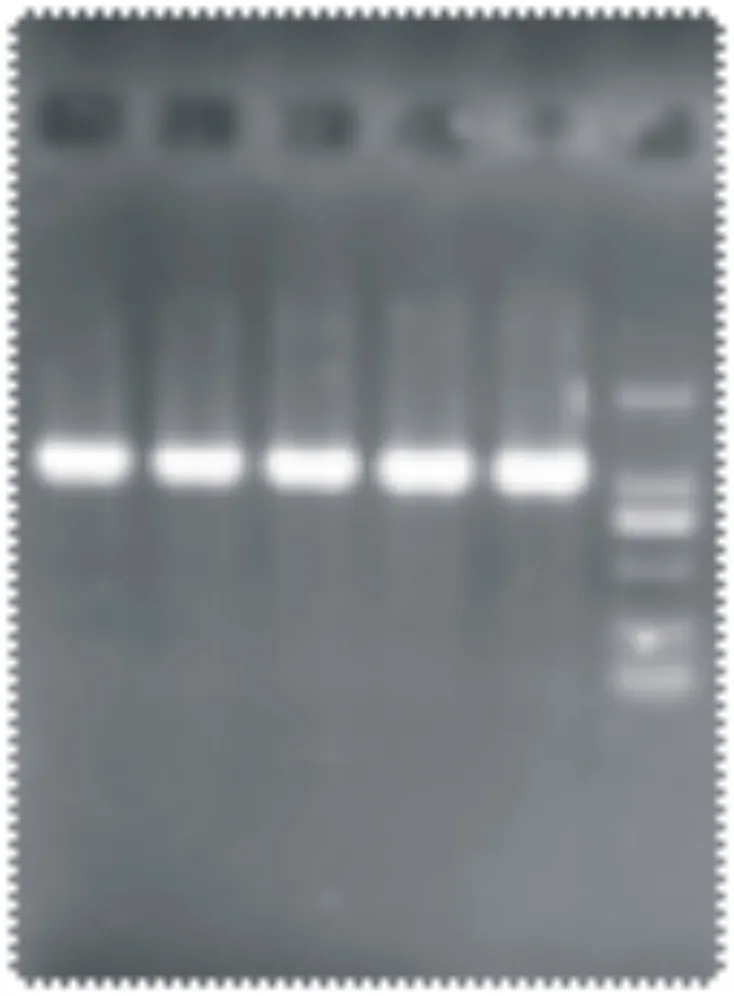
图1 mtDNA D-Loop区 PCR 扩增产物
2.2 核苷酸变异及单倍型分布趋势
早胜牛及其杂交类群mtDNA D-Loop区遗传多态性分析结果表明(表1),mtDNA D-Loop区碱基序列富含A和T,(A+T碱基含量为61.6%,G+C含量38.4%)。在不考虑碱基插入、缺失和Poly(C)末端长度变异的情况下,对长度805 bp的核苷酸序列比对后,发现了80个变异位点,占核苷酸总数的9.94%;检测到转换、颠换、转换和颠换共存3种核苷酸突变类型,且以转换为主。其中,转换位点69个,占总数的86.3%;颠换位点8个,占10%;转换和颠换共存位点3个,占3.7%。
根据变异位点界定了59个单倍型,从单倍型频率来看,H11单倍型频率最高,有13个个体;H9、H14和H20频率依次降低,分别为6、5和3个;H2、H3、H30、H42都为2个;其它51个单倍型频率最低,均为1个。从单倍型类型来看,共享单倍型共8个,分别为H2、H3、H9、H11、H14、H20、H30和H42,特有单倍型51个。从群体类别来看,早胜牛类群、安杂类群、南杂类群特有单倍型数分别为28、18、5个,分别占各自单倍型数的93.33%、85.71%和83.33%。
2.3 核苷酸序列歧义度
参考Genbank上公布的秦川牛、南阳牛、鲁西黄牛、晋南牛、延边牛mtDNA D-Loop区核苷酸序列,采用Clustal W软件对早胜牛、安杂、南杂类群及上述地方黄牛品种mtDNA D-Loop区核苷酸序列进行了品种内和品种间同源性比对,结果表明(表2),品种内歧义度,以鲁西黄牛最高,达到5.32%;南阳牛最低,其它黄牛品种及杂交类群介于之间;品种间歧义度在0.73%~6.20%之间,以早胜牛和南杂类群最低;以晋南牛和安杂类群最高。
2.4 系统发育树
以秦川牛、晋南牛、鲁西黄牛、南阳牛、延边牛、西镇牛、蒙古牛、哈萨克牛等我国地方黄牛及野牦牛、欧洲普通牛、印度瘤牛、非洲瘤牛共21头个体的mtDNA D-Loop区核苷酸序列为对照(序列来源见表3),对早胜牛及其杂交类群86个个体的59种单倍型进行聚类分析,并采用MAGE5.0及DnaSP 5.10 软件构建系统发育树。
结果表明(图2),我国地方黄牛、早胜牛及其杂交类群、瘤牛、欧洲普通牛可以聚为三类,即单倍型组I、单倍型组II、单倍型组III。其中:单倍型组I共66头个体,包括26头早胜牛、6头南杂、18头安杂及2头非洲瘤牛、2头安格斯、1头夏洛来、3头秦川牛、1头晋南牛、1头鲁西黄牛、2头延边牛、2头西镇牛、1头蒙古牛、1头哈萨克牛,可见我国地方黄牛与非洲瘤牛和欧洲普通牛(安格斯、夏洛来)亲缘关系较近;单倍型组II共13头个体,包括7头早胜牛、2头安杂及2头南阳牛、2头印度瘤牛,反映出我国地方黄牛与印度瘤牛聚为一类;单倍型组III仅为1头野牦牛,表明牦牛与我国地方黄牛亲缘关系较远。从系统发育树来看,早胜牛及其杂交类群与秦川牛的亲缘关系最近。
表1 早胜牛及其杂交类群mtDNA D-Loop区核苷酸变异位点

表2 部分黄牛品种(类群)品种内与品种间mtDNA D-Loop区序列歧义度

(单位:%)
注:ZS为早胜牛、AZ为安格斯×早胜牛杂交一代、NZ为南德温×早胜牛杂交一代、QC为秦川牛、NY为南阳牛、LX为鲁西黄牛、JN为晋南牛、YB为延边牛。

图2 系统发育树(以普通牛和瘤牛为对照)

表3 不同牛种(类群)Genebank号及参考来源
3 分析与讨论
3.1 早胜牛及其杂交类群遗传多样性
近年来,对D-Loop区核苷酸序列变异分析已成为哺乳动物线粒体DNA研究的热点。本研究对52头早胜牛、7头南德温×早胜牛杂交一代、27头安格斯×早胜牛杂交一代mtDNA D-Loop区805bp核苷酸序列分析表明,存在59个单倍型,种类相对丰富,这表明经过长期的自然选择和人工选择,早胜牛及其杂交类群已经形成了各自的独特的遗传结构。此外,安杂、南杂类群分别与早胜牛类群存在共享单倍型,这说明虽然各自遗传结构不同,但选育过程都与早胜牛存在密切关系。检测到80个核苷酸变异位点,其中,转换69个,颠换8个,转换和颠换共存3个,这高于Loftus等[5]、雷初朝等[2]报道的同类指标。核苷酸变异类型以转化为主,所占比例高达86.3%,这与马云[4]的研究结果类似,也符合哺乳动物进化的特点。
研究证实[6~8],mtDNA中G+C含量一般在21%~50%之间,其中脊椎动物为37%~50%,本研究发现A+T含量为61.6%,G+C含量为38.4%,这符合上述规律,表明我国地方黄牛品种mtDNA D-Loop区富含碱基A和T,具有较高的遗传多态性。以秦川牛、晋南牛、鲁西黄牛、南阳牛、延边牛、早胜牛及其杂交类群等地方黄牛品种(类群)mtDNA D-Loop区核苷酸序列为对照,通过核苷酸同源性分析比对,发现品种内歧义度为0.58%~5.32%,品种间歧义度为0.73%~6.20%,这都高于Loftus等[5]报道的欧洲牛、印度瘤牛和非洲牛间的同类指标,说明中国地方黄牛品种(类群)线粒体基因组核苷酸变异比较丰富,群体变异程度较高,高于国外部分牛种。
3.2 早胜牛母系遗传背景及起源进化
关于我国黄牛的起源,观点不一。多数研究认为[9~11],我国黄牛具有普通牛和瘤牛两个不同的母系起源,另有研究表明[12],我国黄牛起源于普通牛、瘤牛、印度野牛、非洲瘤牛等,还有一个不明背景的母系起源[13]。本研究结果表明,我国地方黄牛分为两大支系,多数与非洲瘤牛、欧洲普通牛聚为一类,少数与印度瘤牛聚为一类,这进一步证实了我国地方黄牛为普通牛和瘤牛的混合起源学说。当然,也有文献报道[4],我国黄牛和牦牛亲缘关系较近,而与水牛亲缘关系较远,尚有待于进一步深入研究。与其他地方黄牛品种相比,早胜牛及其杂交类群与秦川牛的亲缘关系最近,这与陇东地区引入秦川牛作为母本开展纯繁纯育和杂交选育,长期受秦川牛血统的影响有关。
根据《中国牛品种志》[14],我国地方黄牛可以划分为中原黄牛、北方黄牛和南方黄牛三类。本研究中涉及的部分黄牛品种,按照上述分类,秦川牛、早胜牛、鲁西黄牛、晋南牛属于中原黄牛,延边牛、蒙古牛和哈萨克牛属于北方黄牛,南阳牛和西镇牛属于南方黄牛。从系统发育树可以看出,除南方黄牛为印度瘤牛支系,中原黄牛和北方黄牛多数属于普通牛和瘤牛支系,这与北方黄牛受普通牛的影响大,南方黄牛受瘤牛的影响大和中原黄牛受普通牛和瘤牛影响大的说法基本一致[15]。当然,关于母系遗传背景及起源学说的研究,既要深入开展分子生物学研究,又要综合考虑历史原因、地理隔离、驯化过程及环境差异等因素,才能客观评价不同品种独特的遗传特征。
[1] 甘肃省畜牧厅主编.甘肃省畜禽品种志[M].兰州:甘肃人民出版社,1986.45-47.
[2] 雷初朝,陈宏,杨公社,等.中国部分黄牛品种mtDNA 遗传多态性研究[J].遗传学报,2004,31(1):57-62.
[3] SACCONE C,GISSI C,LANAVE C,et al. Evolution of the mitochondrial Genetic system:an overview[J].Gene,2000,261(1):153-159.
[4] 马云,于波,徐永杰,等.中国部分地方牛种mtDNA D-loop 区全序列的遗传多样性与系统进化分析[J].信阳师范学院学报(自然科学版),2012,25(2):202-205.
[5] Loftus R T,MacHugh D E,Bradley D G,et al. Evidence for two independent domest ication in catt le. Proc Natl Acad Sci USA,1994,91:2 575-2 761.
[6] 蔡 欣,雷初朝,王珊,等.基于mtDNA cyt b基因变异的中国黄牛种间遗传关系分析[J].四川大学学报(自然科学版),2012,49(5):1 115-1 120.
[7] 李永红,常洪,耿荣庆,等.雷琼牛GH基因第五外显子遗传多样性及其分化[J].生态学报,2007,27(12):5 325-5 330.
[8] 赖松家,刘延鑫,李学伟,等. 四川黄牛品种线粒体DNA 遗传多样性研究[J].畜牧兽医学报,2005,36(9):887-892.
[9] 房兴堂,周艳,陈宏,等.中国黄牛mtDNA D-loop 遗传多样性及起源[J].动物学报,2007,53(5):928-933.
[10] 赖松家,王玲,刘 益,等.中国部分牦牛品种线粒体DNA遗传多态性研究[J].遗传学报,2005,32(5):463-470.
[11] 赵生国,李文彬,陈富国,等.平凉地方牛群体母系遗传背景[J].动物学杂志,2013,48(1):109-117.
[12] 陈幼春,王毓英,曹鹤红,等.中国黄牛生态种特征及其利用方向[M].北京:中国农业出版社,1990:1-5.
[13] 雷初朝,陈宏,胡沈荣.Y染色体多态性与中国黄牛起源和分类研究[J].西北农业学报,2000,23(4):32-36.
[14] 邱怀,秦志锐,陈幼春,等.中国牛品种志[M].上海:上海科技出版社,1988:6-9.
[15] 陈宏,邱怀,詹铁生,等.中国四个地方黄牛品种性染色体多态性的研究[J].遗传,1993,15(4):14-17.
The Analysis of Genetic Polymorphism and Maternal Genetic Background of Zaosheng Cattle
RONG Wei-zhong, XU Jian-feng*, WANG Ke, GUO Hai-long, BAO Guo-jun, SANG Guo-jun, YANG Ming
(InstituteofAnimalHusbandryandVeterinaryinGansuProvince,Pingliang,Gansu744000,P.R.China)
[Objective] In order to evaluate and excavate genetic resources and germplasm resources of Zaosheng cattle, this study studied the genetic polymorphism and maternal genetic background of Zaosheng cattle. [Method] PCR amplification and sequencing of genome DNA were conducted based on the marker sites within mtDNA D-Loop region. And further this study analyzed genetic polymorphism indexes, such as nucleotide polymorphism loci, nucleotide diversity and number of haplotype of the nucleotide sequence of mtDNA D-Loop of 8 indigenous yellow cattle species and 5 introduced species as control, and constructed phylogenetic tree. [Result] mtDNA D-Loop of Zaosheng cattle and Zaosheng cattle hybrid groups were rich in nucleotide A and T. A total of 80 nucleotide mutation loci were detected in this region, containing three types, base transition, transversion and both the two types coexist, in which transition was the main mutation types. Moreover, 59 haplotypes were defined, which contained 8 shared haploid types and 51 unique haploid type. In China, most of indigenous yellow cattle were clustered into African zebu and Common European cattle, and a few was clustered into India zebu. Zaosheng cattle and Hybrid groups had closest relationship with Qinchuan cattle, comparing with other local yellow cattle breed in China. [Conclusion] Zaosheng cattle and Zaosheng hybrid groups were rich in nucleotides mutations and higher population variation degree, and which had the closest genetic relationship with Qinchuan cattle.
Zaosheng cattle;mtDNA D-loop;phylogeny;genetic relationship
2016-11-20 接收日期:2016-12-25
甘肃省技术研究与开发专项计划项目(1207TCYL016)。
容维中(1965-),男,甘肃平凉人,副研究员,研究方向为动物营养及牛羊繁殖技术。
*通讯作者:徐建峰(1981-),男,汉,甘肃天水人,助理研究员,从事分子生物学研究,Email:710401384@qq.com。
S823.2
A
1001-9111(2017)02-0007-06
猜你喜欢
世界科学技术-中医药现代化(2022年3期)2022-08-22
肝博士(2022年3期)2022-06-30
农业科技与信息(2020年14期)2020-12-18
绿色科技(2019年14期)2019-11-19
创新作文(3-4年级)(2019年3期)2019-09-03
江苏农业科学(2019年5期)2019-09-02
陕西农业科学(2019年6期)2019-07-19
中国饲料(2019年19期)2019-03-25
创业家(2015年2期)2015-02-27
