
黑曲霉菌株TLSF2对水中Cu、Cd的吸附机理探讨
2017-08-08 19:52罗鹏林张然然师莉升郑进康
湖北农业科学 2017年13期
关键词:黑曲霉
罗鹏林++张然然++师莉升++郑进++康薇


摘要:黑曲霉(Aspergillus niger)菌株TLSF2是1株新發现的耐重金属真菌,为深入了解该菌株对水中Cu、Cd的吸附机理,借助原子分光光度法,通过吸附试验探讨了菌体对Cu、Cd的吸附能力,用化学试剂逐步提取法分析了Cu、Cd在黑曲霉菌体中的化学形态及其Cu、Cd含量,采用差速离心法分离黑曲霉菌体的亚细胞结构并测定其各组分Cu、Cd含量,并对Cu、Cd处理后的黑曲霉菌株TLSF2的细胞壁进行了红外光谱分析。结果表明,Cd浓度保持50 mg/L条件下,Cu浓度为200 mg/L时,菌株TLSF2对Cu的吸附量最大,达到17.76 mg/g;Cu浓度保持200 mg/L条件下,Cd浓度为50 mg/L时,菌株TLSF2对Cd的吸附量最大,达5.19 mg/g,表明该菌株对Cu、Cd有很强的耐受性和富集能力。并且证实细胞壁是菌株TLSF2富集Cu、Cd的主要部位,Cu、Cd的化学形态均以氯化钠提取态为主。菌株TLSF2吸附Cu的化学基团主要涉及羧基、氨基和羟基等,吸附Cd的化学基团主要包括酰胺基、氨基、羧基、羟基以及磷酸基等。
关键词:黑曲霉(Aspergillus niger)TLSF2;Cu;Cd;吸附机理
中图分类号:X172 文献标识码:A 文章编号:0439-8114(2017)13-2429-05
DOI:10.14088/j.cnki.issn0439-8114.2017.13.008
Discussion on the Absorption Mechanism of Aspergillus niger TLSF2 Strain to Copper and Cadmium in Water
LUO Peng-lina,ZHANG Ran-rana,SHI Li-shenga,ZHENG Jingb,KANG Weia,b
(a.Scholl of Environmental Science & Engineering;b.Hubei Key Laboratory of Mine Environmental Pollution Control & Remediation,
Hubei Polytechnic University, Huanggang 435003, Hubei, China)
Abstract:Aspergillus niger TLSF2 is a new-found fungus with heavy metal tolerance. In the study, in order to understand the adsorption mechanism of the strain to copper and cadmium in the water deeply, a series of experiments have done. The strains adsorption capacity was determined by AAS, the analysis of chemical form was done and content of the copper and cadmium in the A. niger TLSF2 was determined using the chemical reagent extraction step by step. Then the subcellular structure of the strain was separated by differential centrifugation, the copper and cadmium content of the component was determined,and the FTIR result of cell wall treated with copper and cadmium was discussed. The results showed that the largest copper adsorption of strain was up to 17.76 mg/g when the stress of cadmium and copper were 50 mg/L and 200 mg/L, the highest cadmium adsorption was 5.19 mg/g when the cadmium and copper were 50 mg/L and 200 mg/L respectively, which reflected the strains high tolerance and strong adsorption ability to copper and cadmium. In addition, the cell wall were verified the primary adsorbing position to heavy metal, the sodium chloride extraction condition were the main chemical form of the heavy metals combination. The carboxyl, amino and hydroxyl groups were proved the major perssad to integrate the copper, besides, the cadmium binding site included the amide and phosphate group.
Key words:Aspergillus niger TLSF2; copper; cadmium; adsorption mechanism
Cu是生命所必需的微量元素,但过量的Cu不仅会造成环境污染,而且会危及人和动物的生命安全。正常情况下环境中Cd的含量很低,但Cd是一种生命非必需的元素,一旦进入生物体内就会造成永久性的伤害。环境中的Cu、Cd污染源主要有三大类,一类是原生矿物经过自然矿化或者风化作用导致铜元素的过量存在,自然条件下Cu、Cd往往与其他重金属元素造成环境的复合污染;另一类是采矿或冶炼过程中产生的Cu、Cd污染;第三类是工业“三废”的排放、使用Cu、Cd制剂和城市垃圾处理不当等导致的污染[1]。Cu作为一种细胞代谢的抑制剂,在水体中时可毒害微生物群落,使其在水生生物中富集,人类通过食物链摄取过量的Cu会出现Cu中毒反应,严重影响健康甚至造成死亡[2]。
工程、生物、化学和农业治理是目前用于治理重金属污染环境的重要措施。其中生物治理措施利用植物、微生物的营养代谢活动来降低环境中重金属的毒性和重金属含量,实现修复受污染的生态环境,被国际上称作最有前途的重金属污染治理技术[3]。微生物种类繁多,具有分布广、繁殖快和对环境适应强等特点。近年来,微生物在修复重金属污染环境方面的成本低、操作简单和高效低耗等优势,受到研究人员普遍关注[4]。研究表明,微生物通过吸附作用可以降低环境中的重金属含量。微生物细胞壁上的羧基、羟基、琉基等活性基团能够与重金属离子发生配位结合或络合、离子交换等作用,达到吸收重金属离子的效果;微生物还可以通过物理性吸附使重金属离子在细胞壁上沉淀下来[5]。
黑曲霉(Aspergillus niger)是一类丝状真菌,在修复重金属污染土壤和处理含重金属离子、多环芳烃、纺织染料及农药等污染物的废水方面具有较高的研究应用价值。迄今,有关黑曲霉吸附重金属Cr、Pb、Zn、Ni、Hg的研究报道较多[6-15],黑曲霉处理Cu、Cd复合污染水体却很少报道。近年来,湖北理工学院矿区环境污染控制与修复湖北省重点实验室污染环境生物修复研究团队从大冶铜绿山铜铁矿洗矿池底泥中分离到1株Cu、Cd耐受能力较强的真菌菌株,经鉴定为黑曲霉(A. niger TLSF2)。为了深入了解黑曲霉菌株TLSF2修复重金属污染水体的潜力,本研究在实验室条件下探讨了TLSF2对水中Cu、Cd的吸附能力及其作用机制,为该菌株的工程应用提供理论依据。
1 材料与方法
1.1 试验材料
1.1.1 菌株来源 所用黑曲霉菌株TLSF2由湖北理工学院矿区环境污染控制与修复湖北省重点实验室污染环境生物修复研究团队从大冶铜绿山铜铁矿洗矿池底泥分离获得。
1.1.2 培养基 菌株活化使用PDA培养基,液体培养使用额外添加Cu、Cd的PD培养基。
1.2 试验方法
1.2.1 吸附试验 用PDA平板活化培养黑曲霉菌株。取容量为150 mL三角瓶,先加入一定量的黑曲霉菌丝,然后分别加入50 mL已知浓度的CuSO4、Cd(NO3)2贮备液,置于恒温振荡器上进行吸附反应(150 r/min)。待达到吸附平衡后,过滤取出菌丝,用浓HNO3进行消解,原子吸收分光光度法(Varian AA-240FS-GTA120)测菌丝Cu、Cd含量。用公式计算菌体对Cu2+、Cd2+的吸附量,计算公式:Q=C菌×V÷(m×1 000)。式中,Q为吸附量(mg/g);C菌为菌体Cu、Cd的含量(mg/L);m为菌体干重(g);V为反应液体积(mL)。
1.2.2 Cu、Cd在菌体细胞中的化学形态分析 用添加Cu、Cd的PD培养基振荡培养黑曲霉菌3~5 d(28 ℃、150 r/min,下同)。用化学试剂逐步提取法[16-18]分析Cu、Cd在黑曲霉菌体中的化学形态。依次使用乙醇(体积分数80%)、去离子水、氯化钠(1 mol/L)、醋酸(体积分数2%)及HCl(0.6 mol/L)5种提取剂进行逐步提取。上述5种提取剂提取的Cu、Cd化学形态分别为乙醇提取态(FE)、水提取态(FW)、氯化钠提取态(FNaCl)、醋酸提取态(FHAc)和盐酸提取态(FHCl),最后为残留态(Fr)。将上述收集的上清液分别加入烧杯,用浓HNO3消解充分,原子吸收分光光度法测定各组分Cu、Cd含量。
1.2.3 Cu、Cd在菌体的亚细胞分布 用添加Cu、Cd的PD培养基振荡培养黑曲霉。采用差速离心法分离黑曲霉菌体的亚细胞结构[19]。提取液组分为50 mmol/L pH 7.5的Tris-HCl缓冲液、0.25 mmol/L蔗糖和1.0 mmol/L DTT,置于冰箱中预冷。试验时,准确称取1.0 g菌丝鲜样,加入预冷的提取液20 mL,经充分研磨,将匀浆转入离心管中进行差速离心,分步分离细胞的亚细胞组分:①细胞壁组分(F1)。在4 000 r/min下离心10 min所得沉淀;②细胞器组分(F2)。将上一步离心所得上清液在16 000 r/min下离心45 min,所得沉淀;③可溶性组分(F3)。即第二步所得的上清液。此组分主要是细胞质及液泡内的有机物,大分子和无机离子。
收集F1、F2和F3,分别用浓HNO3消解充分,原子吸收分光光度法测各组分的Cu、Cd含量。
1.2.4 菌体亚细胞组分红外光谱分析 用添加Cu、Cd的PD培养基振荡培养黑曲霉3~5 d,过滤取菌丝体,分别置于80 ℃烘箱中烘至恒重研磨备用。采用KBr压片法对菌丝体进行红外光谱分析(FTIR)[20,21]。
2 结果与分析
2.1 黑曲霉对水中Cu、Cd的吸附能力
由图1可知,培养液Cd浓度为50 mg/L条件下,依次向培養基中添加浓度为0(对照)、50、100、150、200、250和300 mg/L CuSO4贮备液后,菌株TLSF2对Cu的吸附量分别为0、1.28、4.71、8.89、17.76、13.70和11.17 mg/g,并且在一定范围内随着Cu浓度的升高,菌株对Cu的吸附量相应增加,当初始Cu浓度为200 mg/L时,吸附达到最大值。试验同时观察到,Cd初始浓度保持50 mg/L不变时,对照中菌株对Cd的吸附量最大为13.91 mg/g,之后,随着Cu浓度的升高,菌株对Cd的吸附量依次降低, Cu浓度为300 mg/L时,菌株对Cd的吸附量最低,为1.17 mg/g。可见,随着共存Cu浓度的升高,菌株对Cd的吸附能力呈下降趋势,Cu的共存对菌株吸附Cd存在较强的抑制作用。
由图2可知,培养液中Cu浓度为200 mg/L条件下,依次向培养基中添加浓度为0(对照)、10、20、30、40、50和60 mg/L Cd(NO3)2贮备液后,黑曲霉对Cd的吸附量分别为0、0.64、1.45、2.52、3.91、5.19和4.99 mg/g,并且在一定范围内随着Cd浓度的升高,菌株对Cd的吸附量逐渐增加,初始Cd浓度为50 mg/L时,吸附量达到最大值。与此同时,Cu初始浓度保持200 mg/L不变时,随着加入Cd浓度的升高,菌株对Cu的吸附量依次降低,对照菌株对Cu的吸附量最大,为21.75 mg/g,Cd浓度为60 mg/L时,菌株对Cu的吸附量最低,为7.54 mg/g。可见,随着共存Cd浓度的升高,菌株对Cu的吸附能力呈下降趋势,Cd的加入对菌株吸附Cu有一定的抑制作用。
2.2 黑曲霉细胞内Cu、Cd的化学形态分析
由表1可知,采用初始浓度50 mg/L Cd分别与初始浓度50、100、150、200、250、300 mg/L Cu联合处理菌株后, Cu初始浓度低于250 mg/L时,菌体Cu总含量逐渐升高,Cu浓度达到250 mg/L以上时,菌体Cu总含量出现下降趋势,且黑曲霉细胞内各化学形态Cu含量随着Cu初始浓度变化也呈现出与吸附总量变化大致相同的趋势。经浓度为50、100、150、200、250、300 mg/L Cu处理后,氯化鈉提取的Cu含量分别为0.46、2.37、4.74、7.46、7.14和6.18 mg/g,分别占菌体Cu总含量的47.92%、48.17%、41.36%、42.53%、39.36%、40.55%。同样条件下,去离子水提取态Cu含量占总含量的比例分别为20.83%、23.58%、28.27%、27.34%、26.00%、27.23%。其他化学提取态的Cu含量明显低于氯化钠提取态和去离子水提取态。Cu含量最低的是残渣态,Cu初始浓度在100 mg/L以内时,残渣态Cu含量接近于0。菌株TLSF2对Cu的化学形态主要为氯化钠提取态。
由表2可知,用初始浓度200 mg/L Cu分别与初始浓度10、20、30、40、50、60 mg/L Cd联合处理黑曲霉,菌株吸附Cd的总含量依次为1.18、2.93、4.13、5.13、5.94、5.50 mg/g,且随着加入Cd初始浓度的升高,菌株对Cd的吸附量呈上升的趋势,其中,氯化钠提取态的Cd含量分别为0.64、1.66、2.20、2.33、2.50、2.20 mg/g,分别占菌体Cd总含量的34.44%、56.66%、53.27%、45.42%、42.09%和40.00%;醋酸提取态Cd含量占总含量的比例分别为13.56%、18.77%、23.97%、27.88%、26.09%和24.18%,去离子水提取态Cd含量略低于醋酸提取态Cd含量。此外,乙醇提取态和残渣态Cd含量均远小于氯化钠提取态Cd含量,特别是残渣态Cd含量,随着Cd浓度升高,其变化值并不显著,且接近于0。结果显示,菌株TLSF2对Cd的化学形态也主要为氯化钠提取态。
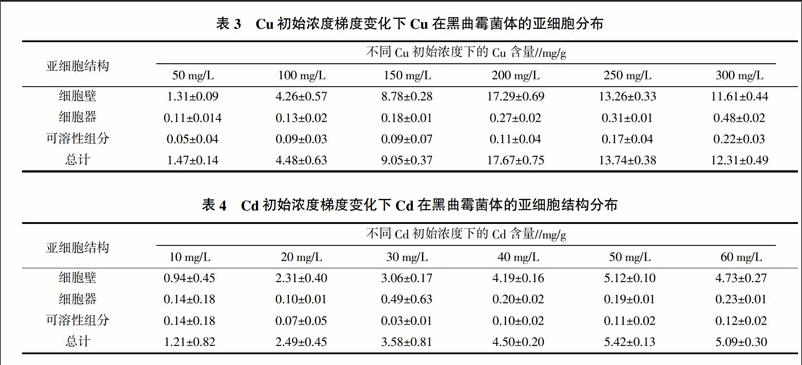
2.3 Cu、Cd在黑曲霉菌体内的亚细胞分布
由表3可以看出,Cu在黑曲霉菌体内主要分布在细胞壁上。细胞壁组分的Cu含量远远高于其余两部分的Cu含量,且随着Cu处理浓度的增加,细胞壁上的Cu含量占菌丝体Cu含量的比例呈先增加后减少趋势,用浓度为50 mg/L的Cd分别与浓度为 50、100、150、200、250、300 mg/L的Cu 联合处理菌体,分布在细胞壁上的Cu含量分别占总含量的89.12%、95.09%、97.02%、97.85%、96.51%和94.31%,并且随着Cu处理浓度的增加,在细胞器组分和可溶性组分上的Cu含量及其占菌株Cu总含量的百分比变化不明显,只有小幅度的差异变化。这表明,A. niger TLSF2菌株吸附Cu时,Cu大部分富集在菌体的细胞壁上。这可能是细胞壁作为一道天然屏障,挡住了培养液中的Cu,使得进入到菌丝细胞内的Cu极少,维持了菌丝细胞内较稳定和较低的Cu含量,增强了菌丝对Cu的耐性。
由表4可知,细胞壁也是黑曲霉富集重金属Cd的主要结构。当采用初始浓度为200 mg/L Cu分别与初始浓度分别为10、20、30、40、50、60 mg/L Cd联合处理菌株,富集在细胞壁上的Cd含量依次为0.94、2.31、3.06、4.19、5.12和4.73 mg/g,分别占菌株Cd总含量的77.69%、92.77%、85.47%、93.11%、94.46%和92.93%。与Cu在细胞壁上的富集率相比,菌株细胞壁对Cd的富集率略低,可能是由于Cd的生物毒性较大,更能破坏菌体细胞壁结构从而进入细胞内部。
2.4 Cu、Cd共存下黑曲霉菌体红外光谱分析
利用傅立叶红外光谱对经Cu、Cd处理的菌株TLSF2细胞组分进行解析,在4 000~500 cm-1波段下,比较对照和处理细胞官能团的红外表征。通过分析对照未经Cu处理菌株细胞组分红外光谱图(图3)可知,在3 500~3 200 cm-1处存在强宽峰,为碳水化合物O-H的对称伸缩振动峰和蛋白质的 -NH的不对称伸缩振动峰共同作用的结果;2 924 cm-1处的峰代表亚甲基,是烷基链中C-H伸缩振动峰;1 622 cm-1附近处的吸收峰是蛋白质酰胺Ⅰ带C=O伸缩振动峰;1 383 cm-1处可能为羧酸盐中 -C(=O)O-的对称与反对称伸缩振动峰;1 314 cm-1处的吸收峰代表胺类中的C-N 伸缩振动;而1 200~900 cm-1之间的吸收峰主要是细胞壁多糖结构中 -C-O-C和-C-O-P伸缩振动峰,1 149 cm-1处的峰主要来自P=O的伸缩振动以及-(=O)O-与-OH的叠加;1 081 cm-1处主要为糖类的C-H和P-O伸缩振动;1 036 cm-1附近处为磷酸基团、糖羟基中的C-O伸缩振动和S=O的伸缩振动共同作用的结果。与无Cu对照相比,经不同Cu浓度(50~300 mg/L)处理72 h的菌株细胞组分红外光谱图并无新的特征峰出现,表明经Cu处理后,菌株的细胞结构并没有改变,只是已有特征峰随着Cu浓度增加发生变化。
由图4可知,低浓度Cd(10~30 mg/L)处理下,特征峰的吸收峰无明显变化;随着Cd浓度的增加,代表-OH和-NH伸缩振动吸收峰发生明显红移(60 mg/L Cd,No.1处),代表-COOH的伸缩振动吸收峰发生蓝移,代表磷酸基团、糖羟基中的 C-O 伸缩振动峰出现红移(50、60 mg/L Cd下),代表酰胺Ⅰ带C=O伸缩振动峰发现蓝移(40、50 mg/L Cd下),代表糖类的C-H和P-O伸缩振动峰峰位變化很小,出现小幅度的红移(40~60 mg/L Cd下)。
综上分析可知,黑曲霉菌株TLSF2在吸附Cu的过程中,主要的结合位点为羟基、羧基和氨基等;在吸附Cd的过程中,结合位点涉及羧基、羟基、氨基、磷酸基、酰胺基等。
3 小结
通过研究黑曲霉菌株TLSF2对水中Cu、Cd的吸附作用,初步得出以下结论。
1)在Cu、Cd共存条件下,随着Cu浓度的升高,菌株对Cd的吸附能力呈下降趋势,Cu的共存对黑曲霉菌株吸附Cd存在较强的抑制作用。同样,随着共存Cd浓度的升高,菌株对Cu的吸附能力呈下降趋势,Cd的加入对菌株吸附Cu有一定的抑制作用。
2)在Cu、Cd共存条件下,随着培养液中Cu、Cd浓度的增加,细胞壁上Cu、Cd含量占总含量的比例随之增多,而进入细胞内的Cu、Cd含量变化不显著。这可能是由于几丁质、蛋白质和多糖等对于丝状真菌吸附与富集重金属起着关键作用的组分主要存在于真菌细胞壁,这些组分能配位结合一些进入到真菌体内的金属离子,通过有效抑制重金属离子的活度来抑制重金属离子在真菌细胞中的跨膜运输。这有利于细胞内重金属离子浓度维持在较低的水平,因此增强了真菌对重金属的耐性和抗性[22]。细胞壁作为一道重要屏障,能够有效阻止重金属进入丝状真菌细胞体。
3)Cu、Cd在黑曲霉菌株TLSF2细胞内的化学形态都以氯化钠提取态为主。
4)羟基、羧基和氨基等是黑曲霉菌株TLSF2细胞与Cu发生作用的主要官能团,菌株吸附Cd结合位点涉及羧基、羟基、氨基、磷酸基、酰胺基等多个官能团。
参考文献:
[1] 彭红云,杨肖娥.香薷植物修复铜污染土壤的研究进展[J].水土保持学报,2005,19(5):197-201.
[2] 向 华,于晓英.铜污染对水体—水生植物的毒害效应研究进展[J].湖南农业科学,2009(11):54-56.
[3] CASTILLO O S,DASGUPTA-SCHUBERT N,ALVARADO C J, et al. The effect of the symbiosis between Tagetes erecta L. (marigold) and Glomus intraradices in the uptake of copper(Ⅱ) and its implications for phytoremediation[J].New Biotechnology,2011,29(1):156-164.
[4] ADKI V S,JADHAV P J,BAPAT V A. At the cross roads of environmental pollutants and phytoremediation:A promising bio remedial approach[J].Journal of Plant Biochemistry and Biotechnology,2013,23(2):125-140.
[5] 张玉刚,龙新宪,陈雪梅.微生物处理重金属废水的研究进展[J]. 环境科学与技术,2008,31(6):58-63.
[6] 李盼盼,臧淑艳,袁 博,等.黑曲霉对模拟废水中汞的吸附[J]. 沈阳化工大学学报,2011,25(1):1-5.
[7] 毕文龙,崔雨琪,方 迪,等.嗜酸硫杆菌和黑曲霉对电镀污泥重金属浸出效果[J].环境工程学报,2014,8(10):4402-4408.
[8] 任婉侠,李培军,李晓军.黑曲霉产酸淋滤去除污染土壤中的重金属[J].中国环境科学,2008,28(8):736-741.
[9] 任婉侠,李培军,耿 涌,等.土壤重金属生物淋滤修复过程中廉价碳源的筛选研究[J].农业环境科学学报,2009,28(9):1837-1843.
[10] 辛 馨.改性黑曲霉吸附水中低浓度U(Ⅵ)的研究[D].湖南衡阳:南华大学,2015.
[11] 陈莹莹.黑曲霉对重金属离子的生物吸附研究[D].沈阳:沈阳理工大学,2013.
[12] 刘 雪.丝状真菌Aspergillus niger A80胞外多聚物的生成特征及其对重金属(Zn、Cd和Pb)的络合效能[D].山东青岛:中国海洋大学,2013.
[13] 崔雨琪,方 迪,毕文龙,等.一株黑曲霉的分离鉴定及其对土壤重金属的生物浸出效果[J].应用与环境生物学报,2014, 20(3):420-425.
[14] 刘 娜.用于去除水中重金属离子的Fe3O4/黑曲霉复合材料的研究[D].长春:吉林大学,2014.
[15] 刘云国,冯宝莹,樊 霆,等.真菌吸附重金属离子的研究[J].湖南大学学报(自然科学版),2008,35(1):71-74.
[16] 于方明,汤叶涛,周小勇,等.镉在圆锥南芥(Arabis paniculata Franch.)中的亚细胞分布及其化学形态[J].中山大学学报(自然科学版),2008,46(6):88-92.
[17] 周守标,徐礼生,吴龙华,等.镉和锌在皖景天细胞内的分布及化学形态[J].应用生态学报,2008,19(11):2515-2520.
[18] 李会合,杨肖娥.硫对超积累东南景天镉累积、亚细胞分布和化学形态的影响[J].植物营养与肥料学报,2009,15(2):395-402.
[19] EREN E. Removal of lead ions by Unye(Turkey) bentonite in iron and magnesium oxide-coated forms[J].Hazard Mater,2009, 165:63-70.
[20] 聂凌鸿,宁正祥.广东淮山药多糖的纯化及化学结构鉴定研究[J]. 林产化学与工业,2004,24(增刊):101-106.
[21] 韦革宏,李素俭,徐万里,等. 根瘤菌菌体及胞外多糖红外光谱特征[J]. 应用与环境生物学报,1999,5(3):310-314.
[22] FAVERO N,COSTA P,MASSIMINO M L. In vitro uptake of cadmium by basidiomycetes Pleurotus ostreatuslil[J]. Biotechnology Letters,1991,13(10):701-704.
猜你喜欢
中国环境科学(2023年9期)2023-09-25
生物技术通报(2018年4期)2018-05-07
中央民族大学学报(自然科学版)(2017年1期)2017-06-11
食品研究与开发(2017年5期)2017-04-11
中国酿造(2016年12期)2016-03-01
中国酿造(2016年12期)2016-03-01
大连工业大学学报(2015年4期)2015-12-11
食品工业科技(2014年7期)2014-03-11
华东理工大学学报(自然科学版)(2014年6期)2014-02-27
食品科学(2013年17期)2013-03-11
