
Structure and function of the contactin-associated protein family in myelinated axons and their relationship with nerve diseases
2017-10-11 02:21YanZouWeifengZhangHaiyingLiuXiaLiXingZhangXiaofangMaYangSunShiyiJiangQuanhongMaDeenXu
中国神经再生研究(英文版) 2017年9期
Yan Zou, Wei-feng Zhang, Hai-ying Liu, Xia Li, Xing Zhang, Xiao-fang Ma, Yang Sun, Shi-yi Jiang, Quan-hong Ma, De-en Xu
Department of Neurology, Te Second People’s Hospital of Wuxi, Wuxi, Jiangsu Province, China
REVIEW
Structure and function of the contactin-associated protein family in myelinated axons and their relationship with nerve diseases
Yan Zou, Wei-feng Zhang, Hai-ying Liu, Xia Li, Xing Zhang, Xiao-fang Ma, Yang Sun, Shi-yi Jiang, Quan-hong Ma, De-en Xu*
Department of Neurology, Te Second People’s Hospital of Wuxi, Wuxi, Jiangsu Province, China
Abstract
The contactin-associated protein (Caspr) family participates in nerve excitation and conduction, and neurotransmitter release in myelinated axons. We analyzed the structures and functions of the Caspr family–CNTNAP1 (Caspr1), CNTNAP2 (Caspr2), CNTNAP3 (Caspr3), CNTNAP4 (Caspr4) and CNTNAP5(Caspr5), Caspr1–5 is not only involved in the formation of myelinated axons, but also participates in maintaining the stability of adjacent connections. Caspr1 participates in the formation, differentiation, and proliferation of neurons and astrocytes, and in motor control and cognitive function. We also analyzed the relationship between the Caspr family and neurodegenerative diseases, multiple sclerosis, and autoimmune encephalitis. However, the effects of Caspr on disease course and prognosis remain poorly understood. Te effects of Caspr on disease diagnosis and treatment need further investigation.
Key Words:nerve regeneration; contactin-associated protein family; myelinated axon; structure; function;Alzheimer’s disease; multiple sclerosis; autism spectrum disorders; neurodegeneration; neural regeneration
Introduction
The main constituents of the brain are neuronal axons,which may be myelinated or unmyelinated. Myelinated neuronal axons offer distinct advantages over unmyelinated axons, such as increased transport speed and decreased axonal diameter (Snaidero and Simons, 2014). Te formation and stability of myelinated axons partly depend on members of the contactin-associated protein (Caspr) family, primarily comprising Caspr1–5 (Spiegel et al., 2002; Weichenhan et al.,2008; Bralten et al., 2010). Te Caspr family are transmembrane proteins with similar structures, though each exhibits functional specificity.
Caspr1, the first member discovered, is located at the paranodes. It forms a complex with contactin and NFASC-155 (NF-155), acts as a barrier between the nodes of Ranvier and internodes, and is involved in the propagation of action potentials and mediation of signal transport (Ullrich et al., 1995; Gollan et al., 2003; Sherman et al., 2005).Furthermore, a recent study indicated that Caspr1 may mediate the timing of neuron and astrocyte differentiation in neural progenitor cells (Wu et al., 2017). Te expression of Caspr1 has also been associated with certain neurodegenerative diseases and may represent a therapeutic target for treatment (Coman et al., 2006; Fan et al., 2013). Recent research has begun to focus on the clinical value of Caspr2, which functions to stabilize resting potential by forming a complex with K+channels (Lancaster et al., 2011). Caspr2 antibodies may be useful as serum markers for the early identification of many neurological conditions, though the clinical ef ficacy of these markers remains to be investigated (Dogan Onugoren et al., 2016). Although little is known regarding the precise functions of the recently identified Caspr3, Caspr4,and Caspr5 proteins, some evidence suggests that Caspr3 is involved in motor control and learning (Peñagarikano et al.,2011; Anderson et al., 2012; O’Roak et al., 2012; Karayannis et al., 2014). A lack of Caspr4 and Caspr5 may be a risk factor for autism spectrum disorders. Here, we briefly discuss the state of research concerning the structures and functions of Caspr family proteins in myelinated axons. This may be useful in the study of the pathology of neural degeneration and other diseases.
Special Structure of Caspr Family Proteins in Myelinated Axons
Structure of myelinated axons
Myelinated fibers are bundled into multiple fascicles by the perineurium and then encased by the epineurium, forming a complete nerve. Te perineurium and epineurium are generated by oligodendrocytes in the central nervous system and Schwann cells in the peripheral nervous system. Te cells are connected by tight junctions, which aid in protecting fibers from external pathogens (Peltonen et al., 2013).
Tough the surrounding cells are different, myelinated axons are compartmentalized into similar parts (Figure 1). Regions wrapped with segments of compact myelin are referred to as internodes. Gaps in the myelin sheath form the nodes of Ranvier. A septate-like junction called the paranode is located on each side of the gap, where surrounding cell membranes attach
to the axolemma. Contactin and Caspr1 are the main elements of paranodes. NF-155 is also located in this region and forms a complex with contactin and Caspr1, bridging the myelin loops to the axolemma. Te region adjacent to the paranode,containing the myelin and axolemma, is known as the juxtaparanode. Voltage-gated K+channels and Caspr2 are present at juxtaparanodes. Te precise structures of different members of the Caspr family are discussed below.
Special Structure of Caspr Family Proteins
Caspr1 is the primary protein of Caspr family
CNTNAP1 is a 190-kDa neuronal transmembrane protein encoded on chromosome 17q21 and highly concentrated at paranodes (Figure 2). As a member of the neurexin superfamily, Caspr1 contains a large extramembrane domain,a single transmembrane region, and a short intracellular region. The extramembrane region contains a factor VII/discoidin region, a laminin G domain, and a fibrinogen domain separated from other factors by another combination of laminin G and epidermal growth factor domains, followed by Pro-Gly-Tyr repeats and another laminin G domain. Te intracellular domain includes a proline-rich sequence capable of binding to a subclass of SH3 domains of specific molecular signals, including Src and Fyn tyrosine kinases (Poliak et al., 1999). The juxta-membrane region of paranodin has high similarity with the glycophorin C region located in the isovalent sequence position. In glycophorin C this domain participates in anchoring the cortical actin/spectrin cytoskeleton to the plasma membrane in red blood cells through its interaction with the N-terminal region of the protein band 4.1.A conserved band 4.1 binding motif is found in glycophorin C, neurexin IV, and paranodin, and hence termed GNP motif,and in several other proteins. Notably, domains similar to the N-terminal region of protein band 4.1 are also found in other proteins and are therefore referred to as four-point-one, ezrin,radixin, moesin (FERM) (Buttermore et al., 2011; Cifuentes-Diaz et al., 2011). FERM functions as a bridge between the cell surface and cytoskeleton, and further research has indicated that protein band 4.1 attaches Caspr to the actin-based cytoskeleton (Denisenko-Nehrbass et al., 2003; Horresh et al.,2010; Buttermore et al., 2011; Cifuentes-Diaz et al., 2011; Einheber et al., 2013). Above all, several domains of Caspr differ in structure and bind to specific receptor proteins.
Caspr2 is another member of Caspr family
Another prominent member of the Caspr family is Caspr2.The extracellular region of Caspr2 is also a mosaic of domains, including signal peptide, discoidin, laminin G, epidermal growth factor, and fibrinogen domains. At the amino acid level, Caspr2 is 45% identical to Caspr1, exhibiting many structural similarities, except that Caspr2 lacks the Pro-Gly-Tyr repeats found near the transmembrane domain of Caspr. Intracellularly, protein band 4.1, not only interacts with Caspr1, but also with Caspr2 (Horresh et al., 2010;Hivert et al., 2016). Both share a common protein band 4.1 binding site, although the intracellular region of Caspr2 is more like glycophorin C and Nrx IV than Caspr1. Caspr2 also contains a short amino acid sequence at its C-terminal,acting as a binding site for the type2 PDZ domain, which is not found in Caspr1. Caspr2 is also contained in a tripartite complex with Shaker-type voltage gated K+channels and the immunoglobulin superfamily molecule transient axonal glycoprotein-1, a glycosyl-phosphatidyl-inositol-anchored cell adhesion molecule, on both axons and in glial cells (Labasque and Faivre-Sarrailh, 2010). While Caspr1 is present at paranodal junctions, Caspr2 is located in juxtaparanodes and some isolated paranodal loops.
Tree additional members of the Caspr family, Caspr3–5,have also been identified (Spiegel et al., 2002; Traut et al.,2006). Researchers have known about the existence of these proteins for a number of years, but little has been uncovered regarding their location, physiological role, and function.Structurally, Caspr3 and Caspr4 are very similar to Caspr1 and Caspr2, as both have the hallmarks of type I transmembrane proteins and similar overall structures to the neurexins (Spiegel et al., 2002). Tese two proteins also have similar extracellular regions formed by specific domains.Te main difference between Caspr3 and Caspr4 lies in the intracellular region. Te cytoplasmic region is formed by 21 and 44 amino acids in Caspr3 and Caspr4, respectively. Both lack the sequence for a protein band 4.1 binding site, yet do contain a binding site for the PDZ domain, like Caspr2 and all neurexins. Caspr3 contains a type III PDZ domain, while,like Caspr2 and neurexins, Caspr4 contains a classical type II PDZ domain. Caspr3 and Caspr4 also differ from one another in distribution. Caspr3 can be found in a variety of cell types, with the exception of Schwann cells, while Caspr4 is expressed mainly in the olfactory bulb, hippocampus, deep cerebellar nuclei, and substantia nigra (Traut et al., 2006).
Functions of Caspr Family Proteins
Generation of myelinated axons
Myelination in the peripheral nervous system begins as a continuous membranous extension from Schwann cells.Instead of the outer myelin tongue moving around the axon, the inner tongue takes an active progression to move underneath the growing sheath. Much research has been dedicated to understanding the generation of myelination in the central nervous system. In the early stages of myelination, myelin is formed by the fusion of different glial processes from one or many oligodendrocytes. Once the initial contact has been made, the myelin membrane advances in a spiraling motion. While myelin acts as a single, flat extension of the membrane in the shape of triangle, its outermost layer maintains direct contact with the cell body, while the innermost layer exhibits the shortest lateral distance to the axon (Snaidero et al., 2014). Myelin growsviathe twining of the leading edge at the inner tongue around the axon(i.e., underneath the deposited membrane). Meanwhile, the membrane layers extend laterally towards the nodal region.Te cytoplasmic-rich pockets of each layer are in close contact with the axolemma, which eventually forms the paranodal loops in the shape of globular particles (Pedraza et al.,2009; Sobottka et al., 2011). As lateral extension proceeds,the glial membrane attaches to the axolemmaviathe axoglial adhesion complex formed by contactin and Caspr1 on axons and NF-155 on glia. Te formation of this complex is required not only for node formation but also for promoting the extension of the membrane layer to the node (Zonta et al., 2008; Susuki et al., 2013; Çolakoğlu et al., 2014). Several studies also have also revealed that the myelin generation process differs between animals and humans. In mice, the process begins at the spinal node at birth and continues until postnatal day 60 (Baumann and Pham-Dinh, 2001). In humans, the peak of myelination occurs in the first year after birth and persists into young childhood, especially in some cortical areas (Fields, 2008).
Participation of Caspr family proteins in axon myelination
Caspr1 is a significant component of the paranode, which participates in its formation and maintains the myelinated axon. While accumulation of Caspr1 at the membrane is detected only after myelination in the peripheral nervous system, the clustering of Caspr1 upon initial glial cell contact is completed prior to the initiation of myelination in the central nervous system (Eisenbach et al., 2009). Before myelination begins, Caspr1 appears as a line on the axon surface. During the initial stages of myelination, when glial cells establish the first contact with axons, Caspr1 appears to be enriched in several places. Caspr1 transforms into a loose coil that accompanies the leading edges of myelin. NF-155 has been observed to cooperate with Caspr1 during this process, eventually co-localizing at the paranode. In the central nervous system, once the contact between the axon and glial cells has been made, NF-155 accumulates at the site of contact, and the complex formed between Caspr1 and NF-155 begins to recruit myelin proteins (Pedraza et al., 2009).
Contactin, another component of this complex, contributes to the sorting of Caspr1. Both proteins assemble in the endoplasmic reticulum during biosynthesis, and the natural association between contactin and Caspr1 may target them to the cell surfacevialipid rafts (Figure 3). The paranodal distribution of these two proteins is interdependent. If contactin is not expressed, Caspr1 is blocked in the endoplasmic reticulum instead of being directed to the axolemma(Bonnon et al., 2003). Association with glycan phosphatidyl inositol-anchored contactin releases Caspr from the lectin-chaperone calnexin, allowing the complex to exit from the endoplasmic reticulum (Bonnon et al., 2003). When Caspr1 is deficient, contactin is not detected at paranodes but is abundant at nodes. Te intracellular domain of Caspr1 interacts with protein band 4.1, and the absence of protein band 4.1 accelerates the internalization of the binding of Caspr1 and contactin (Gollan et al., 2002). This indicates that Caspr1 serves as a transmembrane scaffold protein for the stabilization of the Caspr1/contactin adhesion complexviaconnections to cytoskeletal components. However,Caspr1 serves a number of additional roles as well. During myelination, NF-155 can binding directly to a high molecular weight contactin, though the presence of Caspr inhibits this interaction (Gollan et al., 2003). The resulting expression of low molecular weight contactin, an endoglycosidase H-sensitive isoform, may then bind to Caspr1 but not to NF-155 (Gollan et al., 2003; Sherman et al., 2005). Researchers have therefore suggested that NF-155 may bind directly to the oligosaccharide of contactin (Bonnon et al., 2007). In general, Caspr1 is vital for the construction of complexes required for the formation of myelin during development.
The formation of Caspr2 and K+channels is somewhat different to that for Caspr1 and its partner contactin. Compared to the co-recruitment of Caspr1 and contactin, a distinct mechanism underlies the axonal distribution of Kv1.2 and Caspr2 (Figure 4). The accurate location of K+channels is dependent on two factors: the precise formation of paranodes and the Caspr2/transient axonal glycoprotein-1 complex (Gordon et al., 2014). During myelination, the Kv1/Caspr2 complex is first recruited at paranodes together with Caspr1/protein band 4.1. Following this it is directed to the juxtaparanodesviatransient axonal glycoprotein-1 in an asymmetrical fashion (Hivert et al., 2016). Kv1.2 channels are more highly concentrated at distal juxtaparanodes, relative to the soma, whereas the doublets of paranodes are symmetrical. The lateral positioning of Kv1/Caspr2 may be induced by their clearance from the rows of septate-like junctions when the paranodal junction becomes compact(Hivert et al., 2016). The final position of K+channels is controlled by a multi-step process. First is dependent on the generation of the paranodal barrier. Lack of any component of the paranode, such as Caspr1 or contactin, may result in the movement of K+channels to the node. Second, the proper placement of K+channels requires Caspr2. In contrast to the accumulation of K+channels at paranodes lacking Caspr1, genetic ablation of Caspr2 redistributes K+channels at the mesaxonal line along the internode. In the absence of both Caspr proteins, Kv1 channels form large aggregates along the internodal axolemma. When the paranodal region is impacted by dysfunctional Caspr1, Caspr2 to some extent compensates for the barrier function at this site (Gordon et al., 2014). Terefore, Caspr2 also participates in the formation of myelinated axons and contributes somewhat to the stability of axons.
Other Function of Caspr Family Proteins
In addition to participating in myelination, Caspr1 also contributes to the formation and maintenance of the components responsible for the stability of myelin, acts as a biochemical and electrical barrier between internodes and the nodes of Ranvier, and participates in the generation of action potentials. Nodes of Ranvier are rich in Na+channels and responsible for the propagation of action potentialsviasaltatory conduction. Mutations resulting in Caspr1 deficiency in mice result in the components of nodes dispersing along axons and inhibiting the transition from Na1.2 to Na1.6 in the central nervous system. However, in the peripheral nervous system, the effects of Caspr1 deficiency are more modest:Te length of nodes increases and maturation is delayed, although all nodes eventually express Na1.6 (Rios et al., 2003).Additionally, myelinated axons lacking Caspr1 also exhibit aberrant mitochondrial function in nodal regions. These mitochondria are typically large and occupy prominent varicosities of the nodal axolemma (Einheber et al., 2006). In Caspr1-deficient mice, K+channels mislocate to the paranodes. Tese clusters persist in the peripheral nervous system,but not the central nervous system (Rios et al., 2003).

Figure 1 Structure of myelinated axons.
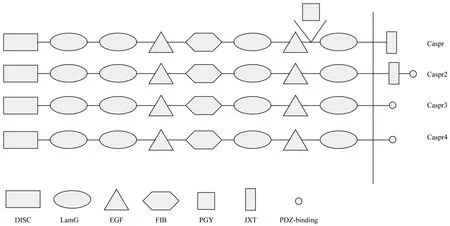
Figure 2 Mosaic structures of different members of the Caspr family.
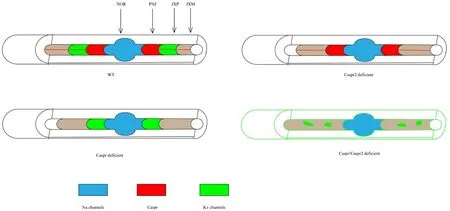
Figure 3 Distribution of ion channels and cell adhesion molecules in distinct axonal subdomains in the central nervous system.
In addition to the displacement of ion channels caused by the disruption of Caspr1, alterations to the paranode itself can also lead to changes in action potentials (Rosenbluth et al., 2012). Recent research has indicated that the mean width of the junctional gap in Caspr1-null mice is significantly larger than that in control mice, and that the range of lengths observed exhibits greater variation in Caspr1-null than control mice (Rosenbluth et al., 2012). Further studies have indicated that this difference can be attributed to the disruption of transverse bands formed by Caspr1, contactin, and NF-155. Overall, disruption of Caspr1 results in impaired saltatory conduction and mislocation of ion channels.
Caspr1 also plays an essential role in the development of the cerebral cortex, regulating the time sequence of fate determination in neural progenitor cells (Wu et al., 2017) (Figure 5). Caspr1 is highly expressed in retinal ganglion cells during the early neurogenic period, while it is mainly expressed in cortical neurons during the gliogenic period. Caspr1 acts to influence the fate of neurons and astrocytes by modulating the timing of the generation of neurons and astrocytes from progenitor cells. Caspr1 deficiency in neural progenitor cells leads to the delayed generation of upper- and deep-layer cortical neurons, and to the precocious formation of astrocytes. Precocious astrogenesis may lead to several neurodevelopmental conditions associated with mental impairment,including Costello syndrome (Paquin et al., 2009), Noonan syndrome (Gauthier et al., 2007), neurofibromatosis-1, andcardiofaciocutaneous syndrome (Urosevic et al., 2011). It is worthwhile to also note that astrocyte dysfunction may be a causative factor in the pathogenesis of autism spectrum disorders and other psychiatric disorders (Sloan and Barres, 2014).Caspr1 may mediate the development of such conditions by activating the Notch pathway. Receptor function in the Notch pathway depends on proteolytic cleavage events and the release of the intracellular domain into the nucleus (Wu et al.,2017). Above all, Caspr1 seems to play a decisive role in the timing of neuron and astrocyte developmentviarepression of the Notch signaling pathway, and may be a decisive factor in the pathogenesis of psychological diseases.
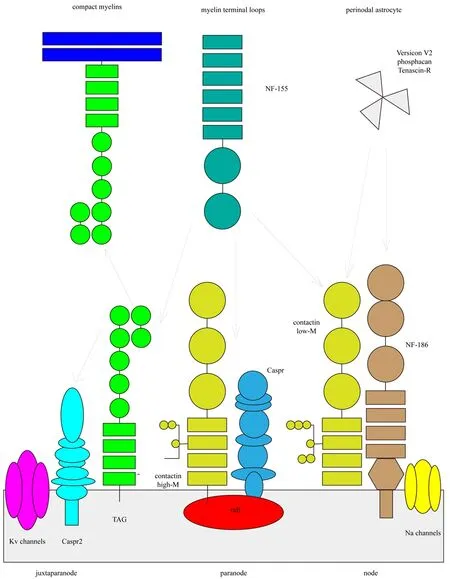
Figure 4 Caspr family proteins form myelinated axons.
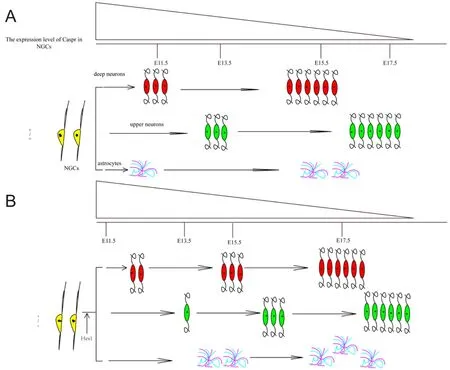
Figure 5 Schematic model for the role of Caspr in the timing of neuron and astrocyte generation.
In mice, Caspr3 is expressed abundantly in the basal ganglia between the first and second postnatal weeks (Hirata et al., 2016b). Considering that the basal ganglia is involved in motor control and learning, this indicates that Caspr3 may be responsible for the regulation of these processes.Recent research has revealed that alterations in Caspr3 may delay motor learning (Hirata et al., 2016a). However, no differences in spontaneous activity, anxiety-like behavior,and nociception have been observed between knock-out and wild-type animals (Hirata et al., 2016a). Furthermore,no significant differences in social behavior, grooming, or prepulse inhibition have been noted when compared with knock-down ofCaspr2orCaspr4(Peñagarikano et al., 2011;Karayannis et al., 2014). Caspr3 may also contribute to neuronal activity associated with motor learning, although the precise mechanisms by which this occurs remain unknown.
Caspr4 is expressed abundantly in neural progenitor cells,which may reflect its involvement in the self-renewal and differentiation of neural progenitor cells into neurons and glial cells. In neural progenitor cells, Caspr4 promotes neuronal differentiation and inhibits proliferationviabinding the ligand of Numb protein X2 (Yin et al., 2015). However,further research is required to elucidate the mechanisms underlying these effects.
Clinical Significance of Caspr Family Proteins
Caspr1 participates in the pathology of Alzheimer’s disease
Caspr family proteins have been implicated in several neurodegenerative disorders, such as Alzheimer’s disease and multiple sclerosis, as well as autism spectrum disorders. Alzheimer’s disease is a neurodegenerative disorder that results in loss of memory and cognitive function. Tere are two pathological markers of Alzheimer’s disease: amyloid plaques and neurofibrillary tangles (Fan et al., 2013). Amyloid plaques are formed by the aggregation of amyloid-β peptides (Kurz and Perneczky, 2011), which are generated by the cleavage of amyloid precursor proteins. Usually, amyloid precursor protein is cleared by α-secretase and γ-secretase, but in pathological cases it is differentially cleared by β-secretase and γ-secretase. Some research has suggested that Caspr1 may form a γ-secretase associated complex, indicative of its participation in the pathology of Alzheimer’s disease (Hur et al.,2012). Additional research has demonstrated that Caspr1 is distributed around amyloid plaques in the cerebral cortex of mice and interacts with amyloid precursor protein. Indeed,over-expression of Caspr1 inhibits that of amyloid precursor protein (Fan et al., 2013), although the precise mechanism underlying this process remains unknown.
Caspr1 acts as the marker of multiple sclerosis
Multiple sclerosis is a chronic, in flammatory, neurodegenerative disease of the central nervous system. Te characteristic neurological decline is attributed to neuronal and axonal loss caused by chronic local inflammation and generalized immune system activation in the central nervous system(Frischer et al., 2009; Weiner, 2009; Lassmann et al., 2012).Multiple sclerosis is further characterized by the presence of activated microglia, neurodegeneration, a compromised blood-brain barrier, and increased glutamate concentration(Howell et al., 2006, 2010). In multiple sclerosis affected areas, all nodal, paranodal, and juxtaparanodal domains are morphologically disorganized. In the early stages, the paranodal components Caspr1 and NF-155 are found in elongated clusters, while diffused voltage gated K+channels begin to overlap with the original paranodal region (Coman et al., 2006). As the disease progresses the paranodal clusters increase in length, while voltage gated K+channels become severely diffuse. Nav channels are the last to undergo elongation. Caspr1 may therefore serve as a marker indicating the specific stage of the disease and the prognosis. Some research suggests that demyelination downregulates the expression of Caspr1 in lesioned areas, and that Caspr1 reappears in parallel with the formation of new myelin sheaths(Coman et al., 2006; Howell et al., 2006). Te enlargement of Caspr1 is in line with the course of disease (Wolswijk and Balesar, 2003). Furthermore, the enlargement of Caspr1-positive structures may not be limited to the paranodes alone. In conclusion, the changes in expression levels of Caspr1 may indicate impairments in saltatory conduction and further myelin loss.
Caspr2 has been associated with autoimmune encephalitis antibodies as well as childhood epilepsy and tumors. Recent research has revealed that Caspr2 expression changes may occur secondary to epilepsy, and may contribute somewhat to certain comorbidities (Wright et al., 2016). Dogan Onugoren et al. (2016) reported that patients with encephalitis and testing positive for Caspr2 antibodies had accelerated recoveries following immunoadsorption therapy. Additional research suggests that Caspr2 may act as a tumor suppressor and an independent prognostic factor in patients with glioma (Bralten et al., 2010). In several gliomas, genetic alternations and methylation of theCNTNAP2gene are associated with reduced levels of Caspr2, and further analysis has indicated that such changes result in decreased proliferation and increased apoptosis (Bralten et al., 2010).
Ablation of Caspr2, Caspr4, or Caspr5 may be a risk factor for autism spectrum disorders. Caspr2-de ficient mice exhibit defects in cortical neuron migration, decreased numbers of GABAergic interneurons (Peñagarikano et al., 2011),abnormal dendritic arborization, reduced spine densities(Anderson et al., 2012), impaired GluA1 trafficking (Vareaet al., 2015), and autism-like behaviors (Gordon et al., 2016).In contrast, Caspr4-deficient mice exhibit decreased GABAergic neurotransmission and elevated release of dopamine,resulting in abnormal sensory-motor gating and grooming endophenotypes (Peñagarikano et al., 2011). However, the exact mechanisms underlying such alterations remain to be elucidated.
Conclusion
Caspr family proteins not only participate in the generation of myelinated axons, but are also critical for the stability of paranodal junctions. An abundance of research has elucidated the involvement of Caspr1 in neuron and astrocyte generation, differentiation, and proliferation. In addition, Caspr family proteins have been implicated in motor control and learning. In this review, we have discussed the relationship between Caspr family proteins and neurological diseases such as Alzheimer’s disease, multiple sclerosis, autoimmune encephalitis, and childhood epilepsy. Te contributions of these proteins to disease courses and prognoses remain unknown. Further research is required to elucidate the precise functions of Caspr family proteins and to identify potential novel techniques for diagnosis and treatment.
Acknowledgments:We are very grateful to Mrs. Quan-hong Ma for her guidance in preparation of this paper.
Author contributions:YZ, XL, YS, HYL, XZ and XFM contributed to literature retrieval. YZ, WFZ and SYJ participated in paper writing. DEX and QHM were in charge of paper modi?cation. All authors approved the?nal version of the paper.
Conflicts of interest:None declared.
Plagiarism check:Checked twice by iTenticate.
Peer review:Externally peer reviewed.
Open access statement:Tis is an open access article distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 3.0 License, which allows others to remix, tweak, and build upon the work non-commercially, as long as the author is credited and the new creations are licensed under the identical terms.
Anderson GR, Galfin T, Xu W, Aoto J, Malenka RC, Südhof TC (2012)Candidate autism gene screen identifies critical role for cell-adhesion molecule CASPR2 in dendritic arborization and spine development.Proc Natl Acad Sci U S A 109:18120-18125.
Baumann N, Pham-Dinh D (2001) Biology of oligodendrocyte and myelin in the mammalian central nervous system. Physiol Rev 81:871-927.
Bonnon C, Goutebroze L, Denisenko-Nehrbass N, Girault JA,Faivre-Sarrailh C (2003) Te paranodal complex of F3/contactin and caspr/paranodin traffics to the cell surface via a non-conventional pathway. J Biol Chem 278:48339-48347.
Bonnon C, Bel C, Goutebroze L, Maigret B, Girault JA, Faivre-Sarrailh C (2007) PGY repeats and N-glycans govern the traf ficking of paranodin and its selective association with contactin and NF-155. Mol Biol Cell 18:229-241.
Bralten LB, Gravendeel AM, Kloosterhof NK, Sacchetti A, Vrijenhoek T, Veltman JA, van den Bent MJ, Kros JM, Hoogenraad CC, Sillevis Smitt PA, French PJ (2010) The CASPR2 cell adhesion molecule functions as a tumor suppressor gene in glioma. Oncogene 29:6138-6148.
Buttermore ED, Dupree JL, Cheng J, An X, Tessarollo L, Bhat MA (2011)Te cytoskeletal adaptor protein band 4.1B is required for the maintenance of paranodal axoglial septate junctions in myelinated axons.J Neurosci 31:8013-8024.
Cifuentes-Diaz C, Chareyre F, Garcia M, Devaux J, Carnaud M, Levasseur G, Niwa-Kawakita M, Harroch S, Girault JA, Giovannini M,Goutebroze L (2011) Protein 4.1B contributes to the organization of peripheral myelinated axons. PLoS One 6:e25043.
Çolakoğlu G, Bergstrom-Tyrberg U, Berglund EO, Ranscht B (2014)Contactin-1 regulates myelination and nodal/paranodal domain organization in the central nervous system. Proc Natl Acad Sci U S A 111:E394-E403.
Coman I, Aigrot MS, Seilhean D, Reynolds R, Girault JA, Zalc B,Lubetzki C (2006) Nodal, paranodal and juxtaparanodal axonal proteins during demyelination and remyelination in multiple sclerosis.Brain 129:3186-3195.
Denisenko-Nehrbass N, Oguievetskaia K, Goutebroze L, Galvez T,Yamakawa H, Ohara O, Carnaud M, Girault JA (2003) Protein 4.1B associates with both Caspr/paranodin and Caspr2 at paranodes and juxtaparanodes of myelinated fibres. Eur J Neurosci 17:411-416.
Dogan Onugoren M, Golombeck KS, Bien C, Abu-Tair M, Brand M,Bulla-Hellwig M, Lohmann H, Munstermann D, Pavenstadt H,Tolking G, Valentin R, Wiendl H, Melzer N, Bien CG (2016) Immunoadsorption therapy in autoimmune encephalitides. Neurol Neuroimmunol Neuroinflamm 3:e207.
Einheber S, Bhat MA, Salzer JL (2006) Disrupted axo-glial junctions result in accumulation of abnormal mitochondria at nodes of ranvier.Neuron Glia Biol 2:165-174.
Einheber S, Meng X, Rubin M, Lam I, Mohandas N, An X, Shrager P,Kissil J, Maurel P, Salzer JL (2013) Te 4.1B cytoskeletal protein regulates the domain organization and sheath thickness of myelinated axons. Glia 61:240-253.
Eisenbach M, Kartvelishvily E, Eshed-Eisenbach Y, Watkins T, Sorensen A, Tomson C, Ranscht B, Barnett SC, Brophy P, Peles E (2009) Differential clustering of Caspr by oligodendrocytes and Schwann cells.J Neurosci Res 87:3492-3501.
Fan LF, Xu DE, Wang WH, Yan K, Wu H, Yao XQ, Xu RX, Liu CF, Ma QH (2013) Caspr interaction with Amyloid Precursor Protein reduces amyloid-beta generation in vitro. Neurosci Lett 548:255-260.
Fields RD (2008) White matter in learning, cognition and psychiatric disorders. Trends Neurosci 31:361-370.
Frischer JM, Bramow S, Dal-Bianco A, Lucchinetti CF, Rauschka H,Schmidbauer M, Laursen H, Sorensen PS, Lassmann H (2009) Te relation between inflammation and neurodegeneration in multiple sclerosis brains. Brain 132:1175-1189.
Gauthier AS, Furstoss O, Araki T, Chan R, Neel BG, Kaplan DR, Miller FD (2007) Control of CNS cell-fate decisions by SHP-2 and its dysregulation in Noonan syndrome. Neuron 54:245-262.
Gollan L, Salomon D, Salzer JL, Peles E (2003) Caspr regulates the processing of contactin and inhibits its binding to neurofascin. J Cell Biol 163:1213-1218.
Gollan L, Sabanay H, Poliak S, Berglund EO, Ranscht B, Peles E (2002)Retention of a cell adhesion complex at the paranodal junction requires the cytoplasmic region of Caspr. J Cell Biol 157:1247-1256.
Gordon A, Adamsky K, Vainshtein A, Frechter S, Dupree JL, Rosenbluth J, Peles E (2014) Caspr and caspr2 are required for both radial and longitudinal organization of myelinated axons. J Neurosci 34:14820-14826.
Gordon A, Salomon D, Barak N, Pen Y, Tsoory M, Kimchi T, Peles E(2016) Expression of Cntnap2 (Caspr2) in multiple levels of sensory systems. Mol Cell Neurosci 70:42-53.
Hirata H, Takahashi A, Shimoda Y, Koide T (2016a) Caspr3-deficient mice exhibit low motor learning during the early phase of the accelerated rotarod task. PLoS One 11:e0147887.
Hirata H, Umemori J, Yoshioka H, Koide T, Watanabe K, Shimoda Y(2016b) Cell adhesion molecule contactin-associated protein 3 is expressed in the mouse basal ganglia during early postnatal stages. J Neurosci Res 94:74-89.
Hivert B, Pinatel D, Labasque M, Tricaud N, Goutebroze L, Faivre-Sarrailh C (2016) Assembly of juxtaparanodes in myelinating DRG culture: Differential clustering of the Kv1/Caspr2 complex and scaffolding protein 4.1B. Glia 64:840-852.
Horresh I, Bar V, Kissil JL, Peles E (2010) Organization of myelinated axons by Caspr and Caspr2 requires the cytoskeletal adapter protein 4.1B. J Neurosci 30:2480-2489.
Howell OW, Rundle JL, Garg A, Komada M, Brophy PJ, Reynolds R(2010) Activated microglia mediate axoglial disruption that contributes to axonal injury in multiple sclerosis. J Neuropathol Exp Neurol 69:1017-1033.
Howell OW, Palser A, Polito A, Melrose S, Zonta B, Scheiermann C,Vora AJ, Brophy PJ, Reynolds R (2006) Disruption of neurofascin localization reveals early changes preceding demyelination and remyelination in multiple sclerosis. Brain 129:3173-3185.
Hur JY, Teranishi Y, Kihara T, Yamamoto NG, Inoue M, Hosia W,Hashimoto M, Winblad B, Frykman S, Tjernberg LO (2012) Identif ication of novel gamma-secretase-associated proteins in detergent-resistant membranes from brain. J Biol Chem 287:11991-12005.
Karayannis T, Au E, Patel JC, Kruglikov I, Markx S, Delorme R, Heron D,Salomon D, Glessner J, Restituito S, Gordon A, Rodriguez-Murillo L,Roy NC, Gogos JA, Rudy B, Rice ME, Karayiorgou M, Hakonarson H, Keren B, Huguet G, et al. (2014) Cntnap4 differentially contributes to GABAergic and dopaminergic synaptic transmission. Nature 511:236-240.
Kurz A, Perneczky R (2011) Amyloid clearance as a treatment target against Alzheimer’s disease. J Alzheimers Dis 24 Suppl 2:61-73.
Labasque M, Faivre-Sarrailh C (2010) GPI-anchored proteins at the node of Ranvier. FEBS Lett 584:1787-1792.
Lassmann H, van Horssen J, Mahad D (2012) Progressive multiple sclerosis: pathology and pathogenesis. Nat Rev Neurol 8:647-656.
Lancaster E, Huijbers MG, Bar V, Boronat A, Wong A, Martinez-Hernandez E, Wilson C, Jacobs D, Lai M, Walker RW, Graus F, Bataller L, Illa I, Markx S, Strauss KA, Peles E, Scherer SS, Dalmau J (2011)Investigations of caspr2, an autoantigen of encephalitis and neuromyotonia. Ann Neurol 69:303-311.
O’Roak BJ, Vives L, Girirajan S, Karakoc E, Krumm N, Coe BP, Levy R,Ko A, Lee C, Smith JD, Turner EH, Stanaway IB, Vernot B, Malig M,Baker C, Reilly B, Akey JM, Borenstein E, Rieder MJ, Nickerson DA,et al. (2012) Sporadic autism exomes reveal a highly interconnected protein network of de novo mutations. Nature 485:246-250.
Paquin A, Hordo C, Kaplan DR, Miller FD (2009) Costello syndrome H-Ras alleles regulate cortical development. Dev Biol 330:440-451.
Peñagarikano O, Abrahams BS, Herman EI, Winden KC, Gdalyahu A,Dong H, Sonnenblick LI, Gruver R, Almajano J, Bragin A, Golshani P, Trachtenberg JT, Peles E, Geschwind DH (2011) Absence of CNTNAP2 leads to epilepsy, neuronal migration abnormalities and core autism-related deficits. Cell 147:235-246.
Pedraza L, Huang JK, Colman D (2009) Disposition of axonal caspr with respect to glial cell membranes: Implications for the process of myelination. J Neurosci Res 87:3480-3491.
Peltonen S, Alanne M, Peltonen J (2013) Barriers of the peripheral nerve. Tissue Barriers 1:e24956.
Poliak S, Gollan L, Martinez R, Custer A, Einheber S, Salzer JL, Trimmer JS, Shrager P, Peles E (1999) Caspr2, a new member of the neurexin superfamily, is localized at the juxtaparanodes of myelinated axons and associates with K+ channels. Neuron 24:1037-1047.
Rios JC, Rubin M, St Martin M, Downey RT, Einheber S, Rosenbluth J,Levinson SR, Bhat M, Salzer JL (2003) Paranodal interactions regulate expression of sodium channel subtypes and provide a diffusion barrier for the node of Ranvier. J Neurosci 23:7001-7011.
Rosenbluth J, Petzold C, Peles E (2012) Dependence of paranodal junctional gap width on transverse bands. J Comp Neurol 520:2774-2784.
Sherman DL, Tait S, Melrose S, Johnson R, Zonta B, Court FA, Macklin WB, Meek S, Smith AJ, Cottrell DF, Brophy PJ (2005) Neurofascins are required to establish axonal domains for saltatory conduction.Neuron 48:737-742.
Sloan SA, Barres BA (2014) Mechanisms of astrocyte development and their contributions to neurodevelopmental disorders. Curr Opin Neurobiol 27:75-81.
Snaidero N, Simons M (2014) Myelination at a glance. J Cell Sci 127:2999-3004.
Snaidero N, Mobius W, Czopka T, Hekking LH, Mathisen C, Verkleij D, Goebbels S, Edgar J, Merkler D, Lyons DA, Nave KA, Simons M(2014) Myelin membrane wrapping of CNS axons by PI(3,4,5)P3-dependent polarized growth at the inner tongue. Cell 156:277-290.
Sobottka B, Ziegler U, Kaech A, Becher B, Goebels N (2011) CNS live imaging reveals a new mechanism of myelination: the liquid croissant model. Glia 59:1841-1849.
Spiegel I, Salomon D, Erne B, Schaeren-Wiemers N, Peles E (2002)Caspr3 and caspr4, two novel members of the caspr family are expressed in the nervous system and interact with PDZ domains. Mol Cell Neurosci 20:283-297.
Susuki K, Chang KJ, Zollinger DR, Liu Y, Ogawa Y, Eshed-Eisenbach Y,Dours-Zimmermann MT, Oses-Prieto JA, Burlingame AL, Seidenbecher CI, Zimmermann DR, Oohashi T, Peles E, Rasband MN (2013)Tree mechanisms assemble central nervous system nodes of Ranvier. Neuron 78:469-482.
Traut W, Weichenhan D, Himmelbauer H, Winking H (2006) New members of the neurexin superfamily: multiple rodent homologues of the human CASPR5 gene. Mamm Genome 17:723-731.
Ullrich B, Ushkaryov YA, Südhof TC (1995) Cartography of neurexins:more than 1000 isoforms generated by alternative splicing and expressed in distinct subsets of neurons. Neuron 14:497-507.
Urosevic J, Sauzeau V, Soto-Montenegro ML, Reig S, Desco M, Wright EM, Canamero M, Mulero F, Ortega S, Bustelo XR, Barbacid M(2011) Constitutive activation of B-Raf in the mouse germ line provides a model for human cardio-facio-cutaneous syndrome. Proc Natl Acad Sci U S A 108:5015-5020.
Varea O, Martin-de-Saavedra MD, Kopeikina KJ, Schurmann B, Fleming HJ, Fawcett-Patel JM, Bach A, Jang S, Peles E, Kim E, Penzes P(2015) Synaptic abnormalities and cytoplasmic glutamate receptor aggregates in contactin associated protein-like 2/Caspr2 knockout neurons. Proc Natl Acad Sci U S A 112:6176-6181.
Weichenhan D, Traut W, Gongrich C, Himmelbauer H, Busch L,Monyer H, Winking H (2008) A mouse translocation associated with Caspr5-2 disruption and perinatal lethality. Mamm Genome 19:675-686.
Weiner HL (2009) Te challenge of multiple sclerosis: how do we cure a chronic heterogeneous disease? Ann Neurol 65:239-248.
Wolswijk G, Balesar R (2003) Changes in the expression and localization of the paranodal protein Caspr on axons in chronic multiple sclerosis. Brain 126:1638-1649.
Wright S, Geerts AT, Jol-van der Zijde CM, Jacobson L, Lang B, Waters P,van Tol MJ, Stroink H, Neuteboom RF, Brouwer OF, Vincent A (2016)Neuronal antibodies in pediatric epilepsy: Clinical features and longterm outcomes of a historical cohort not treated with immunotherapy. Epilepsia 57:823-831.
Wu ZQ, Li D, Huang Y, Chen XP, Huang W, Liu CF, Zhao HQ, Xu RX,Cheng M, Schachner M, Ma QH (2017) Caspr controls the temporal specification of neural progenitor cells through notch signaling in the developing mouse cerebral cortex. Cereb Cortex 27:1369-1385.
Yin FT, Futagawa T, Li D, Ma YX, Lu MH, Lu L, Li S, Chen Y, Cao YJ,Yang ZZ, Oiso S, Nishida K, Kuchiiwa S, Watanabe K, Yamada K,Takeda Y, Xiao ZC, Ma QH (2015) Caspr4 interaction with LNX2 modulates the proliferation and neuronal differentiation of mouse neural progenitor cells. Stem Cells Dev 24:640-652.
Zonta B, Tait S, Melrose S, Anderson H, Harroch S, Higginson J, Sherman DL, Brophy PJ (2008) Glial and neuronal isoforms of Neurofascin have distinct roles in the assembly of nodes of Ranvier in the central nervous system. J Cell Biol 181:1169-1177.
Copyedited by Yu J, Li CH, Qiu Y, Song LP, and Zhao M.
How to cite this article: Zou Y, Zhang WF, Liu HY, Li X, Zhang X, Ma XF, Sun Y, Jiang SY, Ma QH, Xu DE (2017) Structure and function of the contactin-associated protein family in myelinated axons and their relationship with nerve diseases. Neural Regen Res 12(9):1551-1558.
Funding: Tis study was supported by a grant from the Instructional Program of Science and Technology Bureau of Wuxi City of China, No.0302B010507130039PB.
*Correspondence to:De-en Xu, Ph.D.,xudeen@126.com.
orcid:0000-0001-9718-8457(De-en Xu)
10.4103/1673-5374.215268
Accepted: 2017-06-10

- 中国神经再生研究(英文版)的其它文章
- Tp53 gene mediates distinct dopaminergic neuronal damage in different dopaminergic neurotoxicant models
- Unusual neural tract between injured fornix and pedunculopontine nucleus in a patient with traumatic brain injury
- Promises and pitfalls of immune-based strategies for Huntington’s disease
- Post electrical or lightning injury syndrome: a proposal for an American Psychiatric Association’s Diagnostic and Statistical Manual formulation with implications for treatment
- Odorants could elicit repair processes in melanized neuronal and skin cells
- Neuroprotective effects of erythropoietin on neurodegenerative and ischemic brain diseases: the role of erythropoietin receptor