
蔊菜光合-光响应曲线及模型拟合
2017-11-06 02:53文予陌范增丽黎云祥严鑫艺权秋梅
浙江农业科学 2017年10期
文予陌,范增丽,黎云祥,严鑫艺,权秋梅
(西华师范大学 生命科学学院,四川 南充 637002)
蔊菜光合-光响应曲线及模型拟合
文予陌,范增丽,黎云祥,严鑫艺,权秋梅*
(西华师范大学 生命科学学院,四川 南充 637002)
为给蔊菜生理生态研究及栽培提供基础理论依据,以药用植物蔊菜为研究材料,采用Li-6400便携式光合仪,测定了其光响应曲线、CO2响应曲线及相关生理参数,探讨了拟合蔊菜光响应曲线的最适模型,同时研究了光合参数对净光合速率的影响。结果表明,改进指数模型是拟合蔊菜光响应曲线的最适模型,其R2为0.987;蔊菜的光饱和点为1 239.380 μmol·m-2·s-1,光补偿点为37.834 μmol·m-2·s-1,最大净光合速率(CO2)为18.404 μmol·m-2·s-1,暗呼吸速率(CO2)为2.679 μmol·m-2·s-1,表观量子效率为0.075。
蔊菜; 光响应曲线; 模型
蔊菜(RorippaindicaL. Hiem)为十字花科(Cruciferae)蔊菜属(Rorippa)的一年生草本植物[1],又名野油菜。在我国南方地区有较广泛的分布,民间常药用,其主要有效成分为蔊菜素和蔊菜酰胺[2]。在《本草纲目》中记载,“利胸膈,豁冷痰,心腹痛”,有清热下火、祛痰止咳、祛湿的作用,可治疗伤风发烧[3]、老年支气管炎[4]等。目前,蔊菜的研究主要集中在与油菜的远缘杂交[5]、抗旱、抗菌性[6]、形态特性及种子培育等方面[7],未有蔊菜生理方面的相关研究。
由于自然界中不同种类植物甚至同一植物不同器官对环境因子(主要是CO2浓度、光照强度和温度)的响应程度不同[8],而且各个模型的光响应参数差别较大,因此,在进行光响应模型拟合时须考虑模型的适用性。光响应曲线模型数量甚多[9],广泛应用于植物生理生态等方面研究的有指数曲线模型、正切函数曲线模型、二项式回归模型、非直角双曲线模型、直角双曲线模型、Michealis-Menten模型、Farquhar生化模型和直角双曲线改进模型[10]。
本文采用其中最普遍的非直角双曲线模型、直角双曲线模型、直角双曲线改进模型、改进指数模型和指数模型,分别拟合蔊菜的光响应曲线,对比模型估算的生理参数和实测值,分析和讨论模型的适用性。
1 材料与方法
1.1 试验地概况
该试验在西华师范大学生命科学学院试验基地进行,地处四川省南充市(106°04′E,30°49′N),年均温度15.8~17.8 ℃,年降水量980~1 150 mm,海拔300 m,紫色土壤[10],属亚热带湿润性季风气候[11]。
1.2 测定
参照陈根云等[12]的方法,在测量光响应曲线之前,选9株长势良好且一致的野生蔊菜植株,每株选1片健康完整叶测量。在5月份天气晴朗时间段,从早上8点到下午6点,每隔2 h使用便携式光合仪Li-6400(Li-COR,Lincoln, USA)进行光响应测量。具体方法如下:用CO2钢瓶提供碳源,CO2浓度为(395±4.14)μmol·mol-1,叶温(25±0.95)℃,流速500 μmol·s-1,样本室相对湿度65%±5%。光合有效辐射强度用内置红蓝光源设定,光强梯度是:2 000、1 800、1 600、1 400、1 200、1 000、800、600、400、200、150、120、100、80、60、20、0 μmol·m-2·s-1。在光响应曲线测定之前,如光照强度未达到1 000 μmol·m-2·s-1,则使用1 000 μmol·m-2·s-1的红蓝光对被测叶片诱导20 min,再启用光合仪光响应曲线的自动测量程序,测量中要及时进行仪器的自动匹配。测量野生蔊菜叶片的净光合速率(net photosynthetic rate,Pn)。
1.3 数据处理
将光合仪中的数据导出,在Excel表格中初步将光响应测量数据求平均值,用SPSS 23.0软件中的非线性回归模块完成模型公式设定,分别用5个模型对采集的数据进行拟合,得出拟合曲线,光补偿点(light compensation point,LCP)、光饱和点(light saturation point,LSP)、Pn值和决定系数(R2)等参数均能由光响应曲线计算得到[13-14],根据以上参数来衡量各个模型与数据的总体适宜性。
1.4 模型
非直角双曲线模型[15-16]:

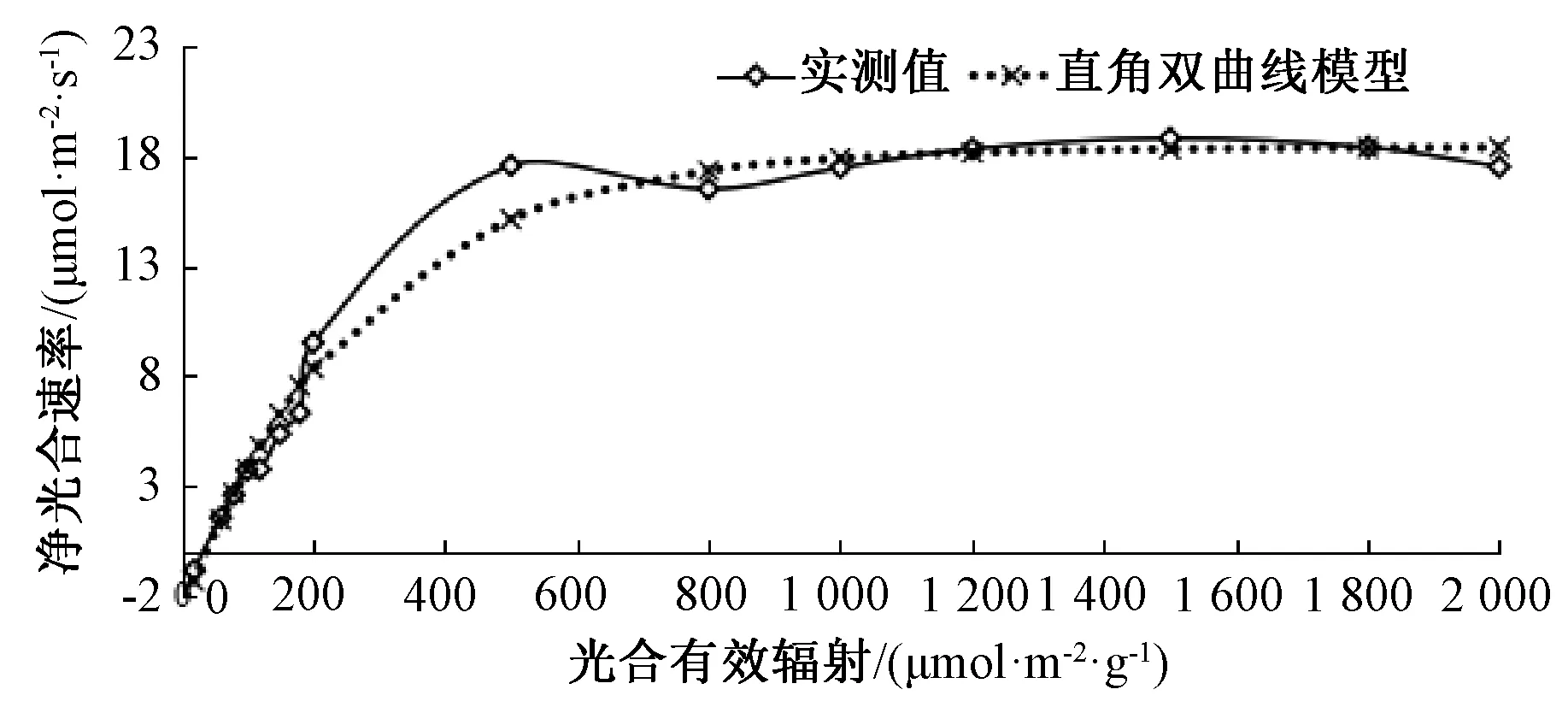
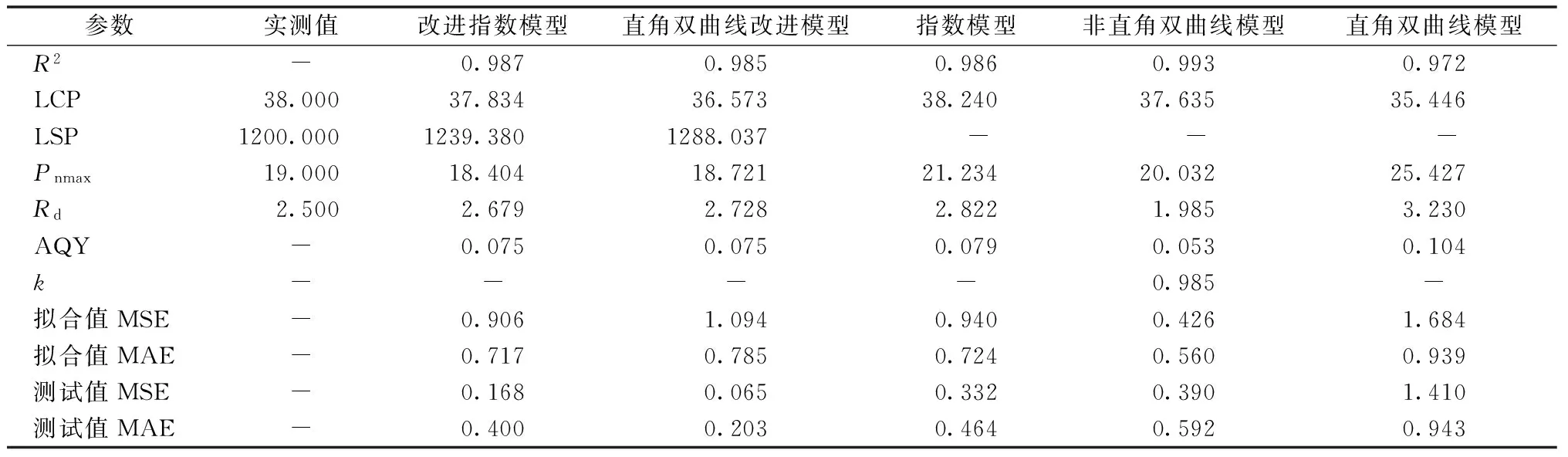
式中:φ为表观量子效率;Pnmax为最大净光合速率;I为光合有效辐射;k为光响应曲角(0 直角双曲线模型[17]: Pn=φIPnmax÷(φI+Pnmax)-Rd。 直角双曲线改进模型[18-19]: Pn=α(1-βI)(I-Ic)÷(1-γI)。 式中:Ic为光补偿点;α是当I=0和I=Ic时,相连两点直线的斜率;β是修正系数;γ是光响应曲线初始斜率与最大净光合速率之比,则γ=α/Pnmax。 指数模型[20]: Pn=Pnmax[1-eφ(I-Ic)]。 改进指数模型[21-22]: Pn=αe-βI-γe-eI。 2.1 改进指数模型的拟合 由图1可知,蔊菜的改进指数模型拟合的决定系数R2=0.987,拟合曲线和实测曲线的变化趋向相同,Pn值均先随着光强的增强快速上升,接着保持平稳,在超过饱和光强后光合效率反而下降,整体拟合效果较好。 图1 蔊菜光响应曲线的改进指数模型 2.2 直角双曲线改进模型的拟合 由图2可知,蔊菜的直角双曲线改进模型拟合的决定系数R2=0.985,拟合结果同改进指数模型,Pn值先随着光强的增加快速上升,接着保持平稳,超过光饱和点后光合效率下降,拟合水平较好。 图2 蔊菜光响应曲线的直角双曲线改进模型 2.3 非直角双曲线模型的拟合 由图3可知,蔊菜的非直角双曲线模型拟合决定系数R2=0.993,虽然决定系数值在5个模型中最大,但与实测值对比,预测值在光强高于1 500 μmol·m-2·s-1后曲线呈持续上升趋势,然而实测值曲线呈下降变化趋向,两者不相符。 图3 蔊菜光响应曲线的非直角双曲线模型 2.4 指数模型的拟合 由图4可知,蔊菜指数模型拟合的决定系数R2=0.986,决定系数比其他模型数值偏小,和非直角双曲线模型一样,光强超过1 500 μmol·m-2·s-1后,预测值曲线呈持续上升趋式,和实测值曲线的下降趋势不符。 图4 蔊菜光响应曲线的指数模型 2.5 直角双曲线模型的拟合 由图5可知,蔊菜的直角双曲线模型拟合的决定系数R2=0.972,决定系数值在5个模型中最小。光强80~200 μmol·m-2·s-1时,预测值与实测值相差较大;光强超过1 500 μmol·m-2·s-1后呈持续上升趋式,与实测值的变化趋势不符合。 2.6 模型适用性分析 表1中的各项生理参数是用蔊菜实测数据分别和5种模型拟合得到的。5种模型的R2值均达到了0.900以上,虽然非直角双曲线模型的R2值最大(0.993),但是该模型在强光下是一条没有极点的渐近线,其拟合的光合速率呈现不断上升的趋势,不存在光饱和点。决定系数R2值的大小只能表明模型的拟合程度高低,并不能保证拟合结果和实际测量值相符合[27],所以不能仅依据R2值的大小判定一个模型的拟合优劣情况。同样,指数模型、直角双曲线模型也是一条没有极点的渐近线,不能解决植株光合效率在高光强时下降的问题。 图5 蔊菜光响应曲线的直角双曲线模型 表1 5个模型的各项生理参数与实测值的比对 注:AQY,内禀量子效率;k,曲角;LCP,光补偿点;LSP,光饱和点;Pn max,最大净光合速率;Rd,暗呼吸速率;MSE,均方误差;MAE,平均绝对误差。 对比表1中实际测量值和改进指数模型、直角双曲线改进模型的预测值:改进指数模型的R2值更大(0.987),并且由改进指数模型估算的LCP值、LSP值、Rd值更接近实际测量值;直角双曲线改进模型估算的Pn max值更接近实际测量值。综合分析以上的生理参数,可以初步推断出拟合值最接近实测值的模型是改进指数模型。 净光合速率是研究植物光合作用的关键指标,光响应曲线是研究光合作用影响因素(CO2浓度、光照强度和温度)的重要手段。若能选择合适的光合光响应模型,则能更准确地反映出植物的光反应过程。本试验中,在用于蔊菜光响应曲线拟合的几个模型中,在光合有效辐射(photosynthetically active radiation,PAR)超过LSP时,表现出Pn值随着PAR的上升而下降的趋势,唯有改进指数和直角双曲线改进模型,符合植物的实际生理生长规律,且与童贯和[28]、朱文旭等[29]研究植物自然条件下的植物生理状态,应当避免使用大于2 000 μmol·m-2·s-1的PAR测量植物的光响应曲线的结论一致[28-31]。其他3个模型的拟合曲线,不能准确解决光饱和区域中净光合速率与光强的相关性问题。 此外,还可以根据均方误差(MSE)和平均绝对误差(MAE)值来比较拟合值和实测值的差异,拟合和测试的MSE和MAE值越小,表明其拟合值越接近实测值,模型的精确度越高。综合MES、MAE结果表明:非直角双曲线模型的拟合值最小,但其计算预测值的MSE、MAE却很大;直角双曲线改进模型的测试值最小,然而其拟合值的MSE、MAE却很大。这种拟合的结果比较好,但预测值和实测值数值极端的情况,表明该模型出现了过拟合现象。陈卫英等[21]的研究表明,不同光合途径下C3、C4植物的光响应曲线用改进指数模型分析描述更准确。 由蔊菜光响应的拟合结果可知,由改进指数模型计算得到的LSP为1 239.380 μmol·m-2·s-1,Pn max(CO2)为18.404 μmol·m-2·s-1,LCP为37.834 μmol·m-2·s-1,Rd(CO2)为2.679 μmol·m-2·s-1。蒋高明[32]判定阴生植物的LCP<20 μmol·m-2·s-1或更低,LSP为500~1 000 μmol·m-2·s-1;阳生植物的LCP为50~100 μmol·m-2·s-1,LSP为1 500~2 000 μmol·m-2·s-1或更高。一般认为,LCP、LSP和Rd值都较高的植物属于阳生植物,在强光环境下才能生长且发育健壮[31]。由此推断,蔊菜是阳生植物,这正好印证蔊菜一般喜欢生长在阳光充足的田野的特性。但是阳生植物在超过自身光饱和点的高光照条件下,其光合速率反而下降,可能是因为色素系统在强光下受到一定程度的破坏。所以,在蔊菜的栽种试验中,为避免植物发生光抑制现象,应尽量种植在适宜的光强环境中,应该控制PAR不超过LSP,在需要的时候可以采取遮荫措施来确保植物正常的光合作用。 [1] 中国科学院中国植物志编辑委员会.中国植物志:第三十三卷[M]. 北京: 科学出版杜, 1987: 300-304. [2] 唐宗俭, 陈嬿, 奚国良. 蔊菜有效成分的研究[J]. 中国科学, 1974 (1): 17-22. [3] 谌立巍, 陈思敏, 吴小唯, 等. 蔊菜提取物体外抗菌活性研究[J]. 内蒙古中医药, 2008, 27(19): 31-32. [4] 佚名. 蔊菜素的实验研究和临床疗效观察[J]. 医学研究杂志, 1972 (3): 19-20. [5] 汤洁, 戴兴临, 张技,等. 甘蓝型油菜与蔊菜远缘杂交选育新品系的主要农艺及产量性状研究[J]. 江西农业学报, 2008, 20(9): 35-37. [6] 涂玉琴, 戴兴临, 涂伟凤, 等. 蔊菜幼苗抗菌核病及抗旱和耐湿特性的鉴定[J]. 植物资源与环境学报, 2011, 20(3): 9-15. [7] 申娟. 秦岭地区藤本植物多样性及园林应用研究[D]. 杨凌: 西北农林科技大学, 2013. [8] DAMESIN C. Respiration and photosynthesis characteristics of current-year stems ofFagussylvatica: from the seasonal pattern to an annual balance [J]. New Phytologist, 2003, 158(3): 465-475. [9] 席晓雪. 大青山针阔叶树种光合特征及其数字模拟[D]. 呼和浩特: 内蒙古农业大学, 2012. [10] 陈兰英, 黎云祥, 钱一凡, 等. 改进指数模型对紫茉莉光合-光响应及CO2响应适用性研究[J]. 广西植物, 2013(6): 839-845. [11] 罗培, 谌柯, 刘辉, 等. 城郊农业区土地利用变化及动因分析:以四川南充市高坪区为例[J]. 资源科学, 2007, 29(4): 179-186. [12] 陈根云, 俞冠路, 陈悦,等. 光合作用对光和二氧化碳响应的观测方法探讨[J]. 植物生理与分子生物学学报, 2006, 32(6): 691-696. [13] 叶子飘, 高峻. 光响应和CO2响应新模型在丹参中的应用[J]. 西北农林科技大学学报(自然科学版), 2009, 37(1): 129-134. [14] 梁文斌, 聂东伶, 吴思政, 等. 短梗大参光合作用光响应曲线及模型拟合[J]. 经济林研究, 2014, 32(4): 38-44. [15] MARSHALL B, BISCOE P V. A model for C3 leaves describing the dependence of net photosynthesis on irradiance. 2. Application to the analysis of flag leaf photosynthesis [J]. Journal of Experimental Botany, 1980, 31(120): 41-48. [16] DIAS-FILHO M B. Photosynthetic light response of the C4 grassesBrachiariabrizanthaandB.humidicolaunder shade [J]. Scientia Agricola, 2002, 59(1): 65-68. [17] CANNELL M G R, THORNLEY J H M. Temperature and CO2responses of leaf and canopy photosynthesis: a clarification using the non-rectangular hyperbola model of photosynthesis[J]. Annals of Botany, 1998, 82(6): 883-892. [18] 叶子飘, 于强. 一个光合作用光响应新模型与传统模型的比较[J]. 沈阳农业大学学报, 2007, 38(6): 771-775. [19] 叶子飘. 光响应模型在超级杂交稻组合-Ⅱ优明86中的应用[J]. 生态学杂志, 2007, 26(8): 1323-1326. [20] POTVIN C, LECHOWICZ M J, TARDIF S. The statistical analysis of ecophysiological response curves obtained from experiments involving repeated measures[J]. Ecology, 1990, 71(4): 1389-1400. [21] 陈卫英, 陈真勇, 罗辅燕, 等. 光响应曲线的指数改进模型与常用模型比较[J]. 植物生态学报, 2012, 36(12): 1277-1285. [22] CHEN Z Y, PENG Z S, YANG J, et al. A mathematical model for describing light-response curves inNicotianatabacum, L [J]. Photosynthetica, 2011, 49(3): 467-471. [23] 李伟成, 王树东, 钟哲科, 等. 几种经验模型在丛生竹光响应曲线拟合中的应用[J]. 竹子研究汇刊, 2009, 28(3): 20-28. [24] 王卓. 叶绿体与蓝藻代谢网络的比较分析及进化研究[D]. 上海: 上海交通大学, 2006. [25] 占杰. 基于MRF先验的PET图像重建与动力学参数估计[D]. 广州: 南方医科大学, 2009. [26] 何绍雄, 曹鉴萍, 黄团华. 非线性房室模型参数计算法[J]. 数值计算与计算机应用, 1986, 7(3): 147-152. [27] 廖小锋, 刘济明, 张东凯, 等. 野生小蓬竹的光合光响应曲线及其模型拟合[J]. 中南林业科技大学学报, 2012, 32(3): 124-128. [28] 童贯和. 不同供钾水平对小麦旗叶光合速率日变化的影响[J]. 植物生态学报, 2004, 28(4): 547-553. [29] 朱文旭, 张会慧, 许楠, 等. 间作对桑树和谷子生长和光合日变化的影响[J]. 应用生态学报, 2012, 23(7): 1817-1824. [30] 蒋高明. 植物生理生态学[M]. 北京: 高等教育出版社, 2004. [31] 韦友欢, 黄秋婵, 王慧珏, 等. 阴生植物与阳生植物色素含量的比较分析[J]. 湖北农业科学, 2010, 49(5): 1126-1129. 收入日期:2017-06-26 文予陌(1994—),女,重庆人,硕士研究生,研究方向为学科教学(生物),E-mail: 1094096394@qq.com。 权秋梅,副教授,E-mail: meimeiq@163.com。 文献著录格式:文予陌,范增丽,黎云祥,等. 蔊菜光合-光响应曲线及模型拟合[J].浙江农业科学,2017,58(10):1779-1783. 10.16178/j.issn.0528-9017.20171034 S634.9 A 0528-9017(2017)10-1779-04 (责任编辑侯春晓)
2 结果与分析




3 讨论
猜你喜欢
环境保护与循环经济(2021年7期)2021-11-02
水产养殖(2021年2期)2021-02-24
哈尔滨轴承(2020年1期)2020-11-03
南方农业学报(2020年4期)2020-06-04
南方农业学报(2020年10期)2020-01-21
中国奶牛(2019年10期)2019-10-28
小学生学习指导(低年级)(2019年9期)2019-09-25
中学生数理化·中考版(2019年8期)2019-07-13
中学生数理化·七年级数学人教版(2019年4期)2019-05-20
电子制作(2018年23期)2018-12-26
