
响应面优化长白落叶松体胚形成早期培养条件的研究
2017-11-11 06:24李淑娟白晓明张含国
植物研究 2017年4期
宋 跃 李淑娟 白晓明 张含国
(东北林业大学林木遗传育种国家重点实验室,哈尔滨 150040)
响应面优化长白落叶松体胚形成早期培养条件的研究
宋 跃 李淑娟*白晓明 张含国
(东北林业大学林木遗传育种国家重点实验室,哈尔滨 150040)
以长白落叶松胚性愈伤组织为材料,通过Box-Behnken Design设计试验,建立以体胚发生量为响应值的响应面模型,优化长白落叶松体胚形成早期的培养条件。结果表明:相对离子浓度(P=0.007)、肌醇浓度(P=0.000)、培养周期(P=0.000)、肌醇浓度与培养周期的交互作用(P=0.007)对长白落叶松体细胞胚胎发生量的影响极为显著。长白落叶松体胚形成早期的最适培养条件为:相对离子浓度26.767 7% BM、肌醇浓度10.454 5 g·L-1、培养周期12.656 6 d,预测体细胞胚胎的发生量为337.04 个·g-1。为便于实际操作,验证试验将培养条件调整为:离子浓度25% BM、肌醇浓度10 g·L-1、培养周期13 d,体胚平均发生量为336.46 个·g-1,与预测值336.29 个·g-1基本一致,体胚萌发率与生根率分别达100%和83.33±5.78%,与前期试验相比,长白落叶松体胚发生的数量和质量均有显著提高。
落叶松属;长白落叶松;响应面优化;体胚发生;胚胎形成早期
落叶松(LarixMill.)具早期速生、抗逆性强、轮伐期短等特点[1],是我国东北地区三大针叶用材树种之一。长白落叶松(Larixolgensis)作为东北地区的优良乡土树种,其种源试验开展较早,自上世纪70年代末以来已有一批速生丰产、抗逆性强的优良长白落叶松家系被陆续选育出来,其栽培区不断向北延伸,推广面积超过60余万hm2[2]。但长白落叶松的种子产量不稳定[3],有性生殖的子代变异大,许多优良性状难以维持,而扦插等无性繁殖也存在生根率较低等问题[2~4],限制了落叶松良种的大规模繁育利用,难以满足林业生产的实际需求。
植物的体细胞胚胎发生具有数量多、繁殖快、结构完整、植株再生率高以及不受季节影响等特点,被认为是一种可实现针叶树大规模繁殖的重要技术手段[4]。通过几十年来的不断努力,落叶松的体细胞胚胎发生技术获得了突破性进展。其中一些种,如欧洲落叶松(Larixdecidua)[5]、日本落叶松(Larixkaempferi)[5~7]、华北落叶松(Larixprincipis-rupprechtii)[8]、杂种落叶松(L.decidua×principis-rupprechtii)(L.decidua×kaempferi)(L.kaempferi×decidua)(L.kaempferi×olgensis)[9~11]等均已通过体胚发生途径获得了再生植株。但就目前而言,想要利用落叶松的无性繁殖来代替种子繁殖依然存在许多技术上的难题[2]。尤其是在体胚发生体系中,优质体胚的大量获得是落叶松体胚规模化生产的重要环节,而高质量的体胚能否大量获得受早期原胚发育状态的直接影响[12~13]。
因此,有必要优化体细胞胚胎形成早期的培养条件,以改善原胚团在体胚成熟前的生理和发育状态。但对落叶松属植物而言,有关该方面的研究报道较少,且尚未见长白落叶松体胚形成早期影响因子研究的详细报道。
响应面法作为一种多因素实验数据回归分析方法[14],具有试验次数较少、可分析实验因素间的交互作用、求得的回归方程精度高等优点,被广泛应用于农业、化学、食品及制造等领域[15]。本研究以长白落叶松胚性愈伤组织为材料,通过Box-Behnken Design设计试验,建立以体胚发生量为响应值的响应面模型,优化长白落叶松体胚形成早期的培养条件,为其良种的规模化繁育和利用奠定基础。
1 材料与方法
1.1 试验材料
以林木遗传育种国家重点实验室(东北林业大学)诱导并保存的长白落叶松胚性愈伤组织为研究对象[4]。供试的胚性愈伤组织避光继代于含2,4-D 0.15 mg·L-1、BA 0.05 mg·L-1、KT 0.05 mg·L-1、肌醇1.0 g·L-1、水解酪蛋白CH 0.5 g·L-1、谷氨酰胺1.0 g·L-1、琼脂6 g·L-1,pH值为(6.00±0.02)的BM培养基上,培养温度为(25±1)℃。取增殖培养约14 d的新鲜组织作为试材。
1.2 体细胞胚胎的成熟
称取约0.2 g长白落叶松胚性愈伤组织,接种到含不同相对离子(无机盐)浓度及肌醇浓度、且不含植物生长调节剂的培养基上进行过渡培养。一定时间后,将上述组织转接于含有ABA 20 mg·L-1、PEG400080 g·L-1、AgNO35 mg·L-1、肌醇0.5 g·L-1、谷氨酰胺0.5 g·L-1、水解酪蛋白0.25 g·L-1、琼脂6 g·L-1,pH值为(6.00±0.02)的BM培养基上,(25±1)℃暗培养,完成体细胞胚胎的成熟,8周后统计获得子叶胚的数量。各处理均设置3次重复,每次重复接种3团胚性愈伤组织。
1.3 响应面法优化体胚形成早期培养条件
前期研究结果[16]的方差分析表明,在长白落叶松体胚形成早期培养阶段,培养周期(P=0.000)、相对离子浓度(P=0.000)及肌醇浓度(P=0.002)对体胚发生量的影响极为显著,而蔗糖(P=0.321)、水解酪蛋白(P=0.600)及谷氨酰胺浓度(P=0.122)的影响并不显著。
因此,本研究着重对培养介质的相对离子浓度、肌醇浓度及培养周期等3个因素进行优化,以每克胚性愈伤组织的体细胞胚胎发生量(个·g-1)为响应值建立响应面模型,试验因素和水平见表1。
表1试验因素和水平
Table1Experimentalfactorsandlevels
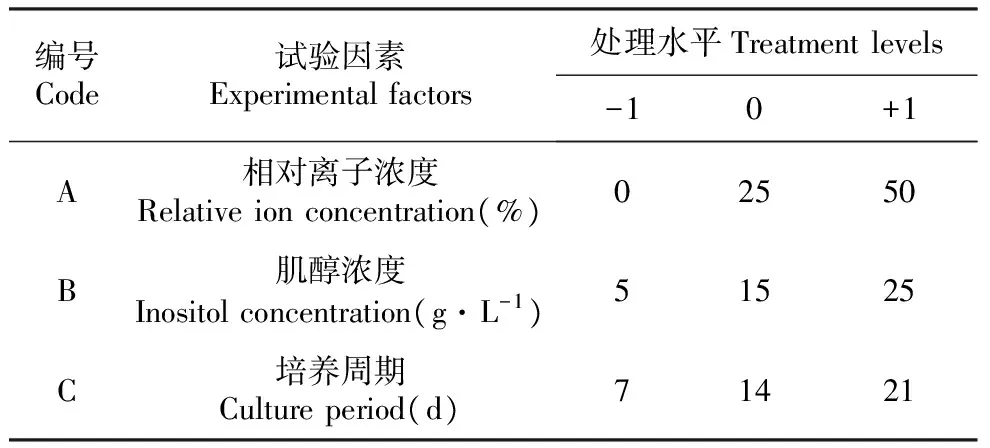
编号Code试验因素Experimentalfactors处理水平Treatmentlevels-10+1A相对离子浓度Relativeionconcentration(%)02550B肌醇浓度Inositolconcentration(g·L-1)51525C培养周期Cultureperiod(d)71421
注:以BM培养基所含有的相对离子浓度为100%,以去离子水所含有的相对离子浓度为0%。
Note:The relative ion concentration in the BM medium was 100%,and the relative ion concentration in deionized water was about 0%.
1.4 验证试验
为验证优化结果的可靠性,同时考虑到试验的可行性,选取各条件优化值的临近整数进行验证,待体胚成熟后,统计子叶胚的数量,并与模型预测结果进行比较。验证共进行3次平行试验,每次试验重复接种3团鲜质量约0.2 g的胚性愈伤组织。
1.5 体细胞胚胎的萌发
选取发育正常的子叶胚,放置在含有蔗糖20 g·L-1,琼脂4.0 g·L-1,VB13.0 mg·L-1,AC 2.0 g·L-1的WPM培养基上进行萌发。每个培养皿接种10个体胚,重复3次。光照时间为16 h·d-1,光照强度约为50 μmol·m-2·s-1。7 d后统计体胚的萌发率,8周后统计生根率。
1.6 统计与分析
每克胚性愈伤组织的体细胞胚胎发生量(个·g-1)=体细胞胚胎发生数/体胚成熟培养前胚性愈伤组织的鲜质量;体细胞胚胎的萌发率(%)=萌发的体胚数/接种的体胚数×100%;体细胞胚胎的生根率(%)=生根的体胚数/接种的体胚数×100%
文中数据使用PASW Statistics 18、Minitab 17软件进行分析,Duncan法进行多重比较。
2 结果与分析
2.1 体胚成熟初期培养条件的响应面优化结果2.1.1 模型的建立及显著性检验
基于BBD(Box-Behnken Design)的试验设计及结果见表2。采用响应面分析法分析试验结果,由此建立以体胚发生量为响应值Y的回归方程:
Y=-71.2+819.6A+12.41B+36.94C-1 474.1A2-0.428 2B2-1.31C2+1.08A·B-2.83A·C-0.299 9B·C
(1)
式中:A为培养介质的相对离子浓度(% BM);B为肌醇浓度(g·L-1);C为培养周期(d)。
方差分析结果(表3)可知,以体细胞胚胎发生量为响应值时,上述二次方程模型极显著(P=0.000),而该模型的失拟不显著(P=0.320),说明该模型的拟合良好,可进行下一步的优化研究[17]。本实验中的3个因素A(P=0.007)、B(P=0.000)、C(P=0.000),二次项A2(P=0.000)、B2(P=0.000)、C2(P=0.000)及B·C(P=0.007)对响应值的影响极显著,而因素A与B(P=0.601)、C间交互作用(P=0.352)的影响不显著。这表明各个具体的试验因素对响应值的影响并不是简单的线性关系。
表2响应面设计及试验结果
Table2Experimentaldesignandresultsofresponsesurfaceoptimizationoftransitioncultureconditions
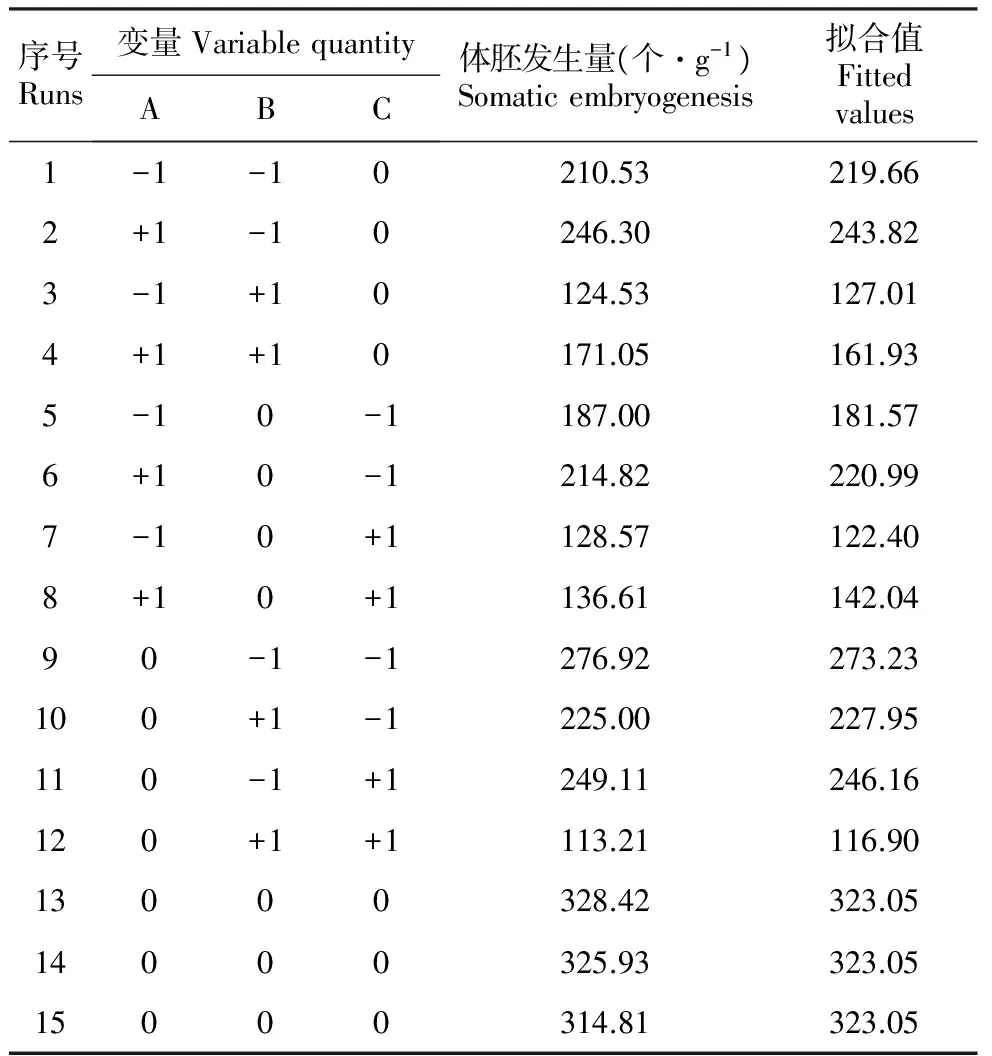
序号Runs变量VariablequantityABC体胚发生量(个·g-1)Somaticembryogenesis拟合值Fittedvalues1-1-10210.53219.662+1-10246.30243.823-1+10124.53127.014+1+10171.05161.935-10-1187.00181.576+10-1214.82220.997-10+1128.57122.408+10+1136.61142.0490-1-1276.92273.23100+1-1225.00227.95110-1+1249.11246.16120+1+1113.21116.9013000328.42323.0514000325.93323.0515000314.81323.05
表3离子浓度、肌醇浓度及培养周期对体胚发生量影响的方差分析
Table3Varianceanalysisoftheeffectofionic,inositolcontentandtheculturedayontheoccurrenceofsomaticembryogenesis
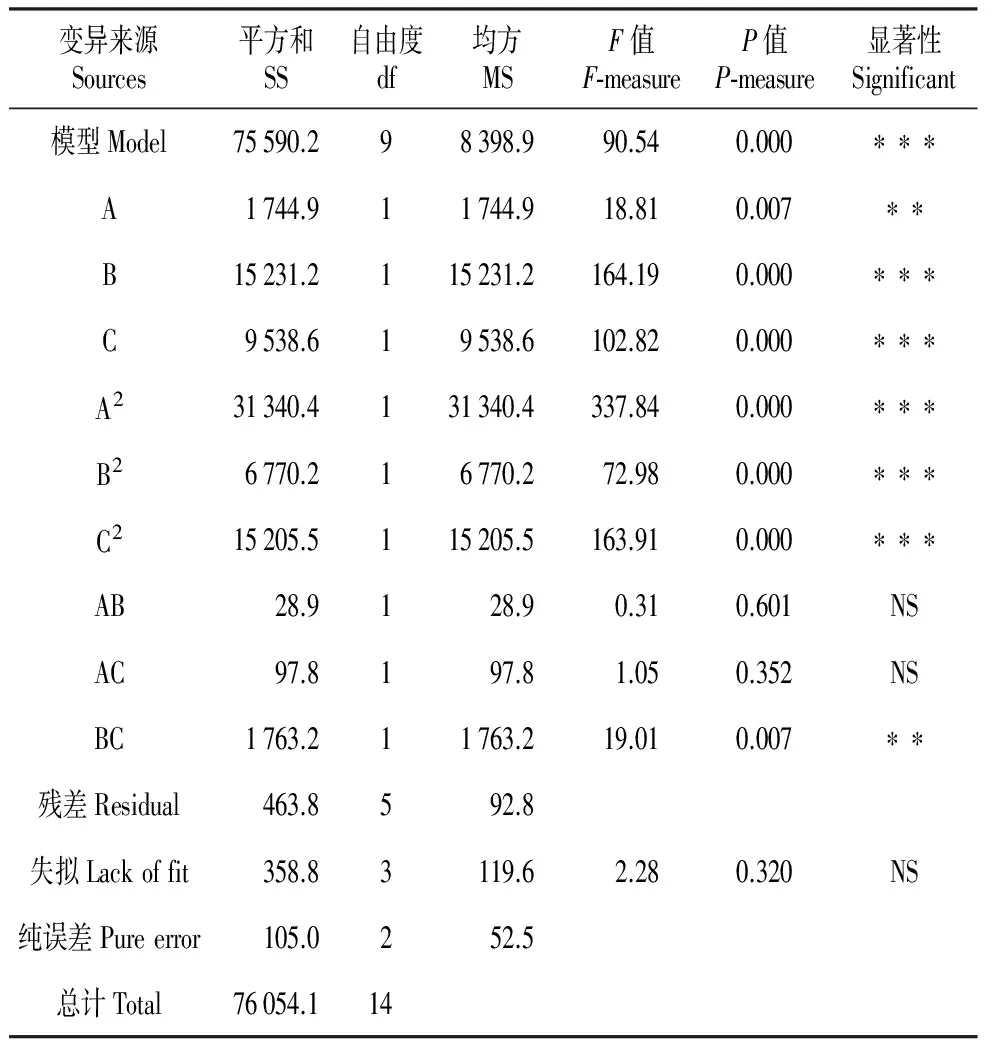
变异来源Sources平方和SS自由度df均方MSF值F⁃measureP值P⁃measure显著性Significant模型Model75590.298398.990.540.000∗∗∗A1744.911744.918.810.007∗∗B15231.2115231.2164.190.000∗∗∗C9538.619538.6102.820.000∗∗∗A231340.4131340.4337.840.000∗∗∗B26770.216770.272.980.000∗∗∗C215205.5115205.5163.910.000∗∗∗AB28.9128.90.310.601NSAC97.8197.81.050.352NSBC1763.211763.219.010.007∗∗残差Residual463.8592.8失拟Lackoffit358.83119.62.280.320NS纯误差Pureerror105.0252.5总计Total76054.114
** .P<0.010; *** .P<0.001; NS.P>0.050
该模型的相关系数R-sq=0.993 9、R-sq(调整)=0.982 9、R-sq(预测)=0.921 4,表明此方程可以解释98.29%的数据,且使用该模型推测的结果较为可靠。此外,变异系数(CV)可以反映模型的置信度,CV值越高,模型的置信度越低,反之相反[18]。本试验的CV=4.44%,说明该模型的置信度较高,模型方程能够真实地反映试验值,可以较好地分析响应值的变化。
2.1.2 响应面交互作用分析与优化
通过Minitab 17软件对各因素之间的交互作用进行响应面分析,并绘制响应面曲线图及等高线图。
从图1~3A可知,每个响应面均为开口向下的凸形曲面,这表明响应值存在最大值[19~21]。在一定范围内调整体胚形成早期阶段的培养条件,体胚发生量会随介质所含的相对离子浓度、肌醇浓度以及培养周期的增加或延长而呈现出先提高后降低的变化趋势。等高线图可以直观地反映出各因素间交互作用对响应值的影响[19]。由图1~3的图B可知,相对离子浓度、肌醇浓度、培养周期间的交互作用均对体胚发生量有一定的影响,其交互作用的影响力排序为:肌醇浓度与培养周期>相对离子浓度与培养周期>相对离子浓度与肌醇浓度。
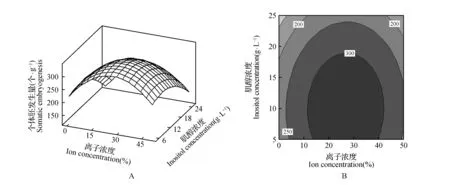
图1 相对离子浓度与肌醇浓度对体胚发生量影响的响应面及等高线Fig.1 Response surface and contour lines effects of ion and inositol content on the occurrence of somatic embryos
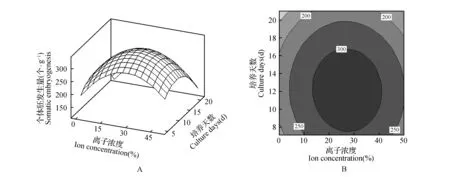
图2 相对离子浓度与培养周期对体胚发生量影响的响应面及等高线Fig.2 Response surface and contour lines effects of ion concentration and synchronized culture period on the occurrence of somatic embryos
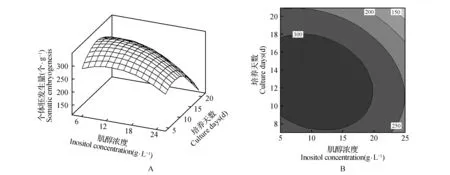
图3 肌醇浓度与培养周期对体胚发生量影响的响应面图及等高线Fig.3 Response surface and contour lines effects of inositol content and synchronized culture period on the occurrence of somatic embryos

图4 长白落叶松胚性愈伤组织及早期原胚显微结构A.长白落叶松早期原胚的显微结构;B.体胚形成早期阶段的长白落叶松胚性愈伤组织Fig.4 L.olgensis embryogenic callus and early proembryo microstructure A. Microstructure of early proembryos of L.olgensis; B. Surface structure of embryogenic callus in early stage of somatic embryogenesis of L.olgensis

图5 长白落叶松体细胞胚胎的成熟Fig.5 The mature somatic embryos of L.olgensis
2.2 最佳培养条件的验证
通过模型优化获得的最适培养条件为:无机盐浓度26.767 7% BM、肌醇浓度10.454 5 g·L-1、培养周期12.656 6 d,体细胞胚胎的发生量为337.04 个·g-1。为便于实际操作,选取各条件的临近整数值,具体为:无机盐浓度25% BM、肌醇浓度10 g·L-1、培养周期13 d,重复试验3次,体胚平均发生量为336.46 个·g-1,与预测值336.29 个·g-1基本一致,表明该方程与实际情况的拟合良好。
2.3体胚成熟早期培养对体胚形态发生及萌发的影响
形态学观察发现,在体胚形成早期培养阶段,胚性愈伤组织逐渐由半透明转变为乳白色,组织表面结构也随之干燥、最终呈绳索状或颗粒状(图4B);镜检原胚的发育状态相对较好,胚柄类似结构较为粗壮、胚头饱满(图4A)。将这些原胚转接于含ABA的成熟培养基上约15 d,即有数量可观、子叶整齐、大小适中、形态修长、呈子弹状的子叶胚出现(图5),并且这些胚胎发生的同步性较高、畸形率低。
相对而言,未经过渡培养而直接进行成熟培养的体胚,其发生量较低、成熟周期相对较长、同步性差、形态大小不一且畸形胚所占比率较高。通常,原胚团接种45 d后才可观察到零星的子叶胚出现,这些胚胎的苗端虽已分化并形成子叶,但仍有部分胚胎根端的胚柄类似结构未能完全消失,导致下胚轴膨大、愈伤化、根原基发育异常,这些畸形胚的萌发较为困难且成苗率极低。
综上所述,在体胚形成的早期阶段,适当改变培养基成份进行一定时间的过渡培养可以促进长白落叶松体胚的发生与形态建成,并能明显提高体胚的数量与质量。最优培养条件下获得的体胚,其萌发率高达100%,生根率也达80%以上(表4);与原有的长白落叶松体胚发生体系[4](子叶胚发生频率相对较低,每克愈伤组织上只发生67.60~179.89个子叶胚,体胚的萌发率仅为75%,而体胚的生根率不足70%)相比有较大改善。
表4体胚成熟早期培养对体胚萌发的影响
Table4Effectsofearlyembryocultureonsomaticembryogermination
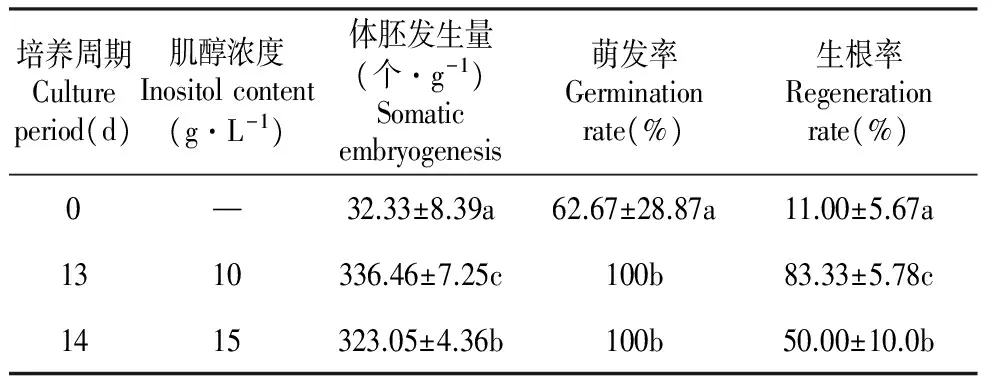
培养周期Cultureperiod(d)肌醇浓度Inositolcontent(g·L-1)体胚发生量(个·g-1)Somaticembryogenesis萌发率Germinationrate(%)生根率Regenerationrate(%)0—32.33±8.39a62.67±28.87a11.00±5.67a1310336.46±7.25c100b83.33±5.78c1415323.05±4.36b100b50.00±10.0b
注:不同字母表示在0.05水平上差异显著。
Note:Different letters indicate significant differences at 0.05 level.
3 讨论
植物的体细胞胚胎发生是一个极为复杂的生物学过程,在体胚发生的不同阶段,原胚团及体细胞胚胎的生理状态有所不同[22]。因此需根据胚胎发育的需求,适当调整培养基的成份。松杉类植物的体细胞胚胎发育可分为三个阶段,即原胚团时期、胚胎形成早期及胚胎形成晚期[23~24]。根据原胚团形态的差异,又可将原胚团时期分为原胚团Ⅰ、原胚团Ⅱ及原胚团Ⅲ等3个阶段[25]。而在这3个阶段的原胚团中,仅有处于原胚团Ⅲ阶段的原胚团才能进一步发育形成成熟的体细胞胚,而增殖培养基中添加的细胞分裂素(如6-BA、KT)与生长素(如2,4-D、NAA)会促使细胞的无序分裂而破坏体胚的极性,导致原胚团Ⅲ破裂[26]。因此在胚胎形成的早期适当改变培养条件,即在体胚成熟早期进行过渡培养利于针叶树体细胞胚胎的形态建成。
培养介质中添加的各类元素在细胞的生命活动过程中均起着不同的重要作用。植物体细胞胚胎发生的不同阶段,胚性细胞对各种无机离子的吸收程度也有所不同[22]。有研究表明,处于不同时期的胚性悬浮细胞中氮元素及矿质元素的含量变化明显,随着培养时间的延长,钙、铜、铁的含量逐渐降低;钾、氮、镁的含量有所增加;而磷的含量则呈现出先增加后急剧降低的趋势[27]。本研究表明,在长白落叶松体胚成熟的早期阶段,适当降低培养基中无机盐的浓度可以显著提高成熟体胚的获得量。推测这种现象的原因可能是:某些无机盐(如KCl、KH2PO4、MgSO4)的浓度过高,影响或抑制了原胚团细胞对其他无机离子(如Zn2+、Cu2+、Co2+、Mn2+等与体胚形成至关重要的离子)的吸收与利用[22],其内在的分子机制尚待进一步研究。
肌醇作为一种小分子有机化合物可以参与渗透压的调节,并为植物的生长提供碳源,其衍生物还参与如:细胞内信号转导、生长素的合成及运输及植物细胞的抗逆调节等多种理化过程[28~33]。有研究表明,肌醇作为合成磷脂酰肌醇以及磷脂酰肌醇磷酸的底物,参与维持内膜系统的结构与功能,从而影响生长素调控植物胚胎的发育过程[33]。日本落叶松及日本×长白落叶松的原胚团经9 g·L-1肌醇高渗处理42 d,胚头的质量与密度较对照组有明显的改善[34]。本文的研究结果表明:肌醇浓度对长白落叶松体胚发生的影响极为显著。在体细胞胚胎成熟的早期阶段,适当提高肌醇浓度可以促进体细胞胚的成熟。
4 结论
本研究对体胚成熟早期阶段培养基中无机盐离子浓度、肌醇浓度及培养周期等因素对长白落叶松体胚发生数量的影响进行了研究,采用响应面法进行培养条件的优化。试验结果表明:相对离子浓度(P=0.007)、肌醇浓度(P=0.000)以及培养周期(P=0.000)对长白落叶松体胚发生量的影响极为显著。模型拟合的最优培养条件为:相对离子浓度26.767 7% BM、肌醇浓度10.454 5 g·L-1、培养周期12.656 6 d,体胚发生量为337.04 个·g-1。实际操作时调整培养条件为:相对离子浓度25% BM、肌醇浓度10 g·L-1、培养周期13 d,体胚的发生量为336.46 个·g-1,其萌发率与生根率分别可达100%、83.33±5.78%,为实现长白落叶松良种的规模化繁育奠定了基础。
1.王伟达,李成浩,杨静莉,等.杂种落叶松未成熟胚的体细胞胚发生和植株再生[J].林业科学,2009,45(8):34-38.
Wang W D,Li C H,Yang J L,et al.Somatic embryogenesis and plantlet regeneration from immature zygotic embryos of hybrid larch[J].Scientia Silvae Sinicae,2009,45(8):34-38.
2.杨书文.落叶松的遗传改良[M].哈尔滨:东北林业大学出版社,1994.
Yang S W.Genetic improvement of larch[M].Harbin:Northeast Forestry University Press,1994.
3.刘足根,朱教君,袁小兰,等.辽东山区长白落叶松(Larixolgensis)种子雨和种子库[J].生态学报,2007,27(2):579-587.
Liu Z G,Zhu J J,Yuan X L,et al.On seed rain and soil seed bank ofLarixolgensisin montane regions of eastern Liaoning Province,China[J].Acta Ecologica Sinica,2007,27(2):579-587.
4.宋跃,甄诚,张含国,等.长白落叶松胚性愈伤组织诱导及体细胞胚胎发生[J].林业科学,2016,52(10):45-54.
Song Y,Zhen C,Zhang H G,et al.Embryogenic callus induction and somatic embryogenesis from immature zygotic embryos ofLarixolgensis[J].Scientia Silvae Sinicae,2016,52(10):45-54.
5.Von Aderkas P,Klimaszewska K,Bonga J M.Diploid and haploid embryogenesis inLarixleptolepis,L.decidua,and their reciprocal hybrids[J].Canadian Journal of Forest Research,1990,20(1):9-14.
6.吕守芳,张守攻,齐力旺,等.日本落叶松体细胞胚胎发生的研究[J].林业科学,2005,41(2):48-52.
Lü S F,Zhang S G,Qi L W,et al.Somatic embryogenesis from immature embryos ofLarixkaempferi[J].Scientia Silvae Sinicae,2005,41(2):48-52.
7.汪小雄,杨映根.日本落叶松体细胞胚胎发生的研究[J].安徽农业科学,2010,38(4):2118-2121.
Wang X X,Yang Y G.Study on the somatic embryogenesis ofLarixleptolepis[J].Journal of Anhui Agricultural Sciences,2010,38(4):2118-2121.
8.齐力旺.华北落叶松体细胞胚胎发生与遗传转化系统建立的研究[D].北京:中国林业科学研究院,2000.
Qi L W.Studies on the somatic embryogenesis and establishment of experimental system inLarixprincipis-Rupprechtii[D].Beijing:Chinese Academy of Forestry,2000.
9.Klimaszewska K.Plantlet development from immature zygotic embryos of hybrid larch through somatic embryogenesis[J].Plant Science,1989,63(1):95-103.
10.Lelu M A,Bastien C,Klimaszewska K,et al.An improved method for somatic plantlet production in hybrid larch(Larix×leptoeuropaea):Part 1.Somatic embryo maturation[J].Plant Cell,Tissue and Organ Culture,1994,36(1):107-115.
11.王伟达,李成浩,杨静莉,等.不同植物生长调节物质对杂种落叶松胚性愈伤组织增殖的影响[J].东北林业大学学报,2008,36(9):5-7.
Wang W D,Li C H,Yang J L,et al.Effect of different plant growth regulators on embryogenic callus proliferation of hybrid larix[J].Journal of Northeast Forestry University,2008,36(9):5-7.
12.中国林业科学研究院林业研究所.一种提高针叶树种体细胞胚胎发生数量与质量的培养基:中国,CN1733906A[P].2006-02-15.
INST Forestry CAF.Medium for improving quantity and quality of conifer somatic embryogenesis:Chinese,CN1733906A[P].2006-02-15.
13.中国林业科学研究院林业研究所.一种改良针叶树种胚性愈伤组织质量的培养基:中国,CN1733905A[P].2006-02-15.
INST Forestry CAF.Culture medium for improving conifer embryogenic callus quality:Chinese,CN1733905A[P].2006-02-15.
14.罗猛,宋卓悦,胡娇阳,等.超声法提取山里红叶总黄酮及其抗氧化活性研究[J].植物研究,2015,35(4):632-637.
Luo M,Song Z Y,Hu J Y,et al.Optimization of ultrasonic-assisted extraction of total flavonoids fromCrataeguspinnatifidaleaves and its antioxidant activities[J].Bulletin of Botanical Research,2015,35(4):632-637.
15.郝学财,余晓斌,刘志钰,等.响应面方法在优化微生物培养基中的应用[J].食品研究与开发,2006,27(1):38-41.
Hao X C,Yu X B,Liu Z Y,et al.The application of response surface methodology in optimization of microbial media[J].Food Research and Development,2006,27(1):38-41.
16.东北林业大学.一种提高长白落叶松体细胞胚发生和植株再生效率的过渡培养基及培养方法:中国,CN105409767A[P].2016-03-23.
Northeast Forestry University.Transitional culture medium improving somatic embryogenesis and plant regeneration efficiency ofLarixolgensisand culture method:Chinese,CN105409767A[P].2016-03-23.
17.李淑娟,詹亚光,杨传平,等.基于响应面法的白蜡属花粉离体萌发培养基优化[J].植物学报,2009,44(2):223-229.
Li S J,Zhan Y G,Yang C P,et al.Response surface methodology to determine the optimal medium for pollen germination for ash[J].Chinese Bulletin of Botany,2009,44(2):223-229.
18.Amenaghawon N A,Nwaru K I,Aisien F A,et al.Application of box-behnken design for the optimization of citric acid production from corn starch usingAspergillusniger[J].British Biotechnology Journal,2013,3(3):236-245.
19.王化,周丽萍,李梦莎,等.微波辅助提取暴马丁香中紫丁香苷和橄榄苦苷的工艺研究[J].植物研究,2016,36(1):141-145.
Wang H,Zhou L P,Li M S,et al.Optimization of microwave-assisted extraction of syringin and oleuropein fromSyringareticulata[J].Bulletin of Botanical Research,2016,36(1):141-145.
20.刘燕,陈小银,杨丽丽,等.响应面优化菊芋菊糖的提取工艺研究[J].植物研究,2016,36(4):627-633.
Liu Y,Chen X Y,Yang L L,et al.Extraction process optimization of inulin fromHelianthustuberosusL. by response surface methodology[J].Bulletin of Botanical Research,2016,36(4):627-633.
21.林建原,季丽红.响应面优化银杏叶中黄酮的提取工艺[J].中国食品学报,2013,13(2):83-90.
Lin J Y,Ji L H.Optimization of flavonoids fromGinkgobilobausing respone surface analysis[J].Journal of Chinese Institute of Food Science and Technology,2013,13(2):83-90.
22.崔凯荣,戴若兰.植物体细胞胚发生的分子生物学[M].北京:科学出版社,2000.
Cui K R,Dai R L.Molecular biology of plant somatic embryogenesis[M].Beijing:Science Press,2000.
23.汪小雄,卢龙斗,郝怀庆,等.松杉类植物体细胞胚发育机理的研究进展[J].西北植物学报,2006,26(9):1965-1972.
Wang X X,Lu L D,Hao H Q,et al.Research advances about developmental mechanism of somatic embryos in conifers[J].Acta Botanica Boreali-Occidentalia Sinica,2006,26(9):1965-1972.
24.Singh H.Embryology of gymnosperms[M].Berlin:Borntrager,1978.
25.Filonova L H,Bozhkov P V,Von Arnold S.Developmental pathway of somatic embryogenesis inPiceaabiesas revealed by time-lapse tracking[J].Journal of Experimental Botany,2000,51(343):249-264.
26.Filonova L H,Bozhkov P V,Brukhin V B,et al.Two waves of programmed cell death occur during formation and development of somatic embryos in the gymnosperm,Norway spruce[J].Journal of Cell Science,2000,113(24):4399-4411.
27.向太和,钟华鑫,梁海曼,等.水稻胚性悬浮细胞系建立过程中的生理生化变化[J].作物学报,1995,21(2):223-119.
Xiang T H,Zhong H X,Liang H M,et al.Physiological and biochemical changes during the establishment of embryogenic suspension cell lines of rice(OryzasativaL.)[J].Acta Agronomica Sinica,1995,21(2):223-119.
28.Irvine R F,Schell M J.Back in the water:the return of the inositol phosphates[J].Nature Reviews Molecular Cell Biology,2001,2(5):327-338.
29.Hall P J,Bandurski R S.[3H]Indole-3-acetyl-myo-inositol hydrolysis by extracts ofZeamaysL. vegetative tissue[J].Plant Physiology,1986,80(2):374-377.
30.Peskan T,Westermann M,Oelmüller R.Identification of low-density Triton X-100-insoluble plasma membrane microdomains in higher plants[J].European Journal of Biochemistry,2000,267(24):6989-6995.
31.Tiede A,Bastisch I,Schubert J P,et al.Biosynthesis of glycosylphosphatidylinositols in mammals and unicellular microbes[J].Biological Chemistry,1999,380(5):503-523.
32.Borner G H H,Sherrier D J,Weimar T,et al.Analysis of detergent-resistant membranes in Arabidopsis.Evidence for plasma membrane lipid rafts[J].Plant Physiology,2005,137(1):104-116.
33.Luo Y,Qin G,Zhang J,et al.D-myo-inositol-3-phosphate affects phosphatidylinositol-mediated endomembrane function inArabidopsisand is essential for auxin-regulated embryogenesis[J].The Plant Cell,2011,23(4):1352-1372.
34.王静.利用胚性愈伤组织进行落叶松遗传转化体系的研究[D].北京:中国林业科学研究院,2007.
Wang J.Studies on the transformation ofLarixbased on the system of somatic embryogenesis[D].Beijing:Chinese Academy of Forestry,2007.
National High-tech R&D Program(“863” Program 2013AA102704-04-03)
introduction:SONG Yue(1991—),male,master,Mainly engaged in larch genetic breeding research.
date:2017-01-06
ResponseSurfaceOptimizationofCultureConditionsfortheFormationofEarlyEmbryosinLarixolgensis
SONG Yue LI Shu-Juan*BAI Xiao-Ming ZHANG Han-Guo
(State Key Laboratory of Tree Genetics and Breeding,Northeast Forestry University,Harbin 150040)
With the larch embryogenic callus, through the Design Box-Behnken design test, we established a response surface model based on the somatic embryogenesis. The culture conditions of somatic embryo early maturation was optimized inLarixolgensis. The relative ion concentration(P=0.007), the inositol concentration(P=0.000), the culture period(P=0.000) and the interaction between inositol content and culture period(P=0.007) effect on the number of somatic embryos ofL.olgensiswere extremely significant. Cultivation of Larix somatic embryo maturation was 337.04 ind·g-1at the early stage of the most optimum conditions for relative ion content of 26.767 7% BM basic culture medium, inositol 10.454 5 g·L-1, culture time 12.656 6 d, and the number of somatic embryos average. In order to facilitate the practical operation, the culture condition was changed to the relative ion concentration of 25% BM, inositol 10 g·L-1, incubation period of 13 d, then the average number of somatic embryos was 336.46 ind·g-1, that was consistent with the predicted value 336.29 ind·g-1. Compared with previous test, the quantity and quality ofL.olgensissomatic embryos were increased significantly.
Larix;Larixolgensis;response surface optimization;somatic embryogenesis;early embryo formation
国家高技术研究发展计划(“863”计划2013AA102704-04-03)
宋跃(1991—),男,硕士研究生,主要从事落叶松遗传育种方面的研究。
* 通信作者:E-mail:lishujuan@126.com
2017-01-06
* Corresponding author:E-mail:lishujuan@126.com
S791.22;Q943.1
A
10.7525/j.issn.1673-5102.2017.04.018
猜你喜欢
新疆农业科学(2022年7期)2022-08-10
中国饲料(2022年6期)2022-04-22
西南农业学报(2021年11期)2021-12-31
饲料工业(2021年21期)2021-12-24
南方农业·中旬(2021年2期)2021-06-24
中南林业科技大学学报(2020年11期)2020-12-08
中南林业科技大学学报(2020年10期)2020-11-03
天津农学院学报(2020年3期)2020-10-14
中国饲料(2019年12期)2019-07-12
现代农业科技(2018年22期)2018-01-15
