
色季拉山不同海拔梯度冷杉幼苗的基本特征及生理指标变化
2017-11-14 09:46刘济铭辛福梅杨小林赵垦田贾黎明
浙江大学学报(农业与生命科学版) 2017年5期
刘济铭,辛福梅*,杨小林,赵垦田,贾黎明
色季拉山不同海拔梯度冷杉幼苗的基本特征及生理指标变化
刘济铭1,2,辛福梅1,2*,杨小林2,赵垦田2,贾黎明11
(1.北京林业大学林学院,北京100083;2.西藏农牧学院资源与环境学院,西藏林芝860000)
对藏东南色季拉山不同海拔梯度(3 800~4 400 m)急尖长苞冷杉(Abies georgei var.smithii)幼苗的基本特征、根系活力、可溶性还原糖、可溶性蛋白质、丙二醛含量、超氧化物歧化酶活性的变化规律进行研究,以阐明急尖长苞冷杉幼苗在不同海拔梯度的变化特征,为研究藏东南林线树种对环境的响应机制提供理论依据。结果表明:1)色季拉山在海拔3 900 m处的环境条件适宜急尖长苞冷杉幼苗生长;2)色季拉山在海拔3 800~4 100 m范围内,急尖长苞冷杉幼苗可溶性蛋白质、还原糖和根系丙二醛含量基本稳定,环境胁迫对其生长无明显影响;3)色季拉山从海拔4 200 m处开始向林线、树线过渡,在海拔4 400 m处外界环境恶劣,急尖长苞冷杉幼苗的苗高、根系活力、叶绿素含量等指标均急剧下降,受环境胁迫影响较大。
色季拉山;海拔;急尖长苞冷杉幼苗;生理指标
植物与环境的关系主要体现在植物的生活史、形态、物候及生理等多个方面,不同植物种类有着不同的适应对策[1]。国内外先后开展了诸多关于遗传基因和环境因子对植物性状作用模式的探索,在所研究的植物表型性状中,叶片对植物营养的获得、利用及利用效率的关系最为密切,能够反映植物适应环境变化所形成的生存对策[2-3]。同样,根系是植物与土壤直接接触的器官,具有支持、吸收、合成和储存有机物等功能,其活性与形态分布直接或间接影响着植物对营养和水分的吸收,进而影响植物地上部分的生长和生态功能的发挥[4]。随着全球碳循环研究的深入,国内外学者对根系构型、生产和周转、空间分布以及影响因素方面的研究热情高涨。研究发现,环境中的温度、水分、盐分及养分状况等都影响着植物根系的生长发育[5],在不同环境条件下植物根系会产生相应的调节机制来适应环境。研究显示:随着海拔升高,许多植物叶面积减小[6],但也有增大或先增大后减小的现象[7];叶绿素含量随海拔升高呈下降趋势[8],但有的变化不明显[9]。植物幼苗的早期生长是决定其将来植被组成和植被类型的重要因素,幼苗对环境变化十分敏感,因而幼苗常用作监测土壤、气候等环境异质性对植物群落组成影响的探针[10]。
青藏高原由于其海拔和气候环境的多变性已成为我国一个独特的地理单元,其东南部分布有全球海拔最高的高山林线,具有以冷杉、云杉和柏木等为主的林线树种[11-12]。由此可见,青藏高原东南部既是世界上最高的林线分布区域,又是气候变化最为明显的区域之一,因而了解该地区林线的生态气候指标可以为分析未来气候情景下林线的可能变化提供基础[13]。然而青藏高原地形复杂,交通不便,海拔高低起伏巨大,人口稀疏,导致对青藏高原的研究长期处于初级阶段,同时,藏东南色季拉山也因此长期处于近自然状态,受人为因素干扰极少,是良好的野外科学研究基地。本研究沿色季拉山海拔梯度(3 800~4 400 m)选择生长良好的7—8年生急尖长苞冷杉(Abies georgei var.smithii)幼苗,通过对其幼苗叶片、根系基本特征及生理指标的研究,以期阐明色季拉山急尖长苞冷杉在不同海拔梯度的变化特征,为进一步解释色季拉山冷杉林对海拔的响应机制提供理论依据。
1 材料与方法
1.1 研究区概况
研究区为自然环境特殊多样、气候类型复杂的藏东南色季拉山(E 93°12′~95°35′,N 29°10′~30°15′),位于西藏雅鲁藏布江大拐弯西北侧,念青唐古拉山与喜马拉雅山脉接合处的工布自然保护区林芝县内。该区气候属于典型的亚高山温带半湿润性气候,干湿季节分明。林芝生态站多年观测显示:该地带年平均气温3.5℃,最暖月(7月)平均气温11.15℃,最冷月(1月)平均气温4.1℃;年降水量1 095.4 mm,主要集中于6—9月;年平均相对湿度83%[14]。该区土壤以酸性棕壤为主,土层较厚,腐殖质化过程明显,土壤pH值在4~6之间[15]。受西南季风及地形的影响,色季拉山阴坡与阳坡的平均温度、极端温度、降雨量等存在较大差异,导致植被群落的反应和垂直带变化较明显,研究区内急尖长苞冷杉(Abies georgei var.smithii)为色季拉山阴坡乔木层的优势种群。
急尖长苞冷杉为松科冷杉亚科冷杉属植物,是青藏高原东南缘亚高山暗针叶林的主要树种之一,主要分布在滇西北、川西南、藏东南的高山地带,特别是在西藏林芝市的米林、察隅、波密等地[13]。它是冷杉属在西藏分布面积最广的树种,为湿润山地亚高山地带的显域植被和顶级群落,是构成藏东南阴坡林线群落的优势树种。在海拔4 300 m左右为色季拉山的森林线,海拔4 300 m以上乔木单株可以成活但不能成林分布,海拔4 400 m左右为树线。色季拉山阴坡为急变型林线,海拔4 400 m为乔木树种分布的上限,阴坡林线群落交错区垂直宽度为70 m。本研究区域植被群落类型的基本特征[16]见表1。
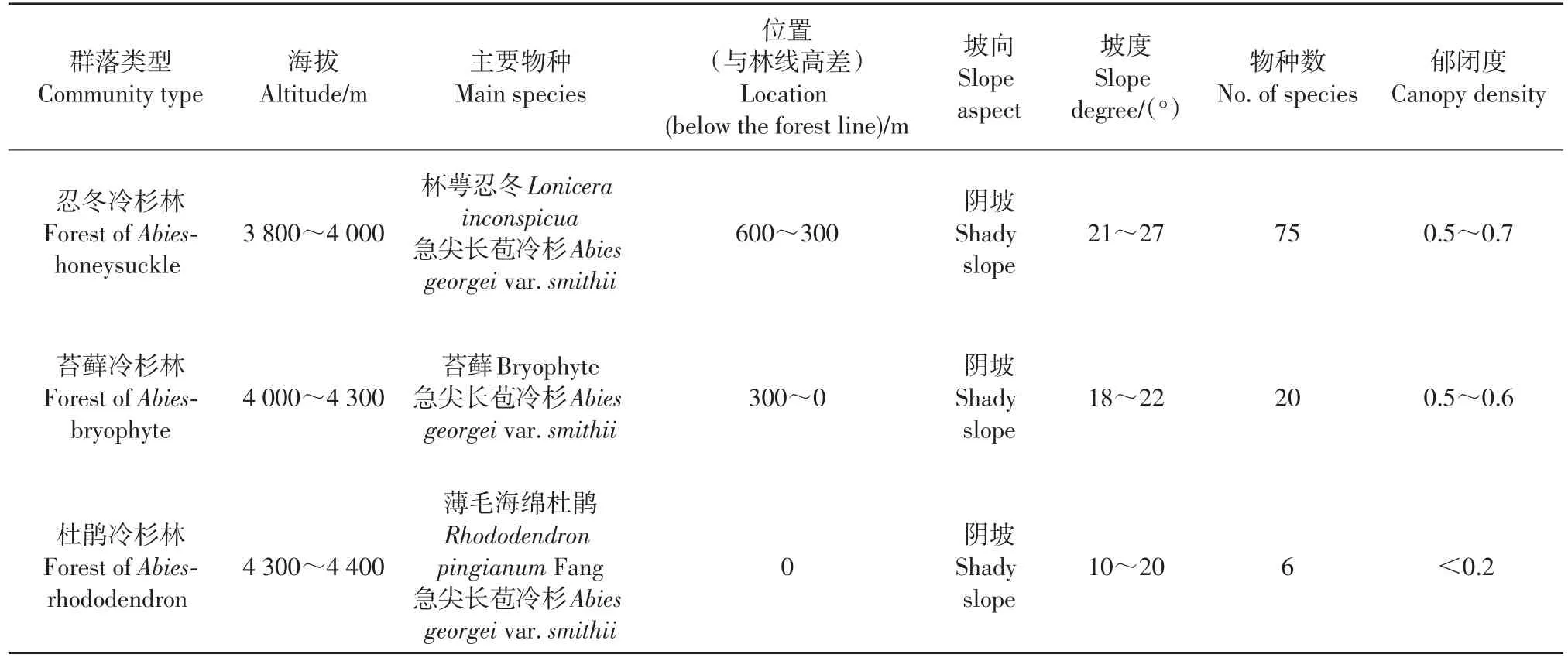
表1 色季拉山林线研究区域植被群落类型基本特征[16]Table 1 Basic characteristics of timberline communities at different altitudes in the Mt.Shergyla[16]
1.2 样品采集及处理
于2014年4月在色季拉山阴坡海拔3 800~4 400 m范围内,以垂直梯度100 m为间隔,设置7个20 m×20 m的研究样地,按年龄结构分别采集50株急尖长苞冷杉幼苗,标记,清水浸泡,流水冲洗、分离后供测定。本研究采用色季拉山不同海拔梯度生长状况良好的7—8年生急尖长苞冷杉幼苗为样本测定分析。
1.3 指标测定方法
1.3.1 基本生长指标测定
采用精度为0.02 mm的游标卡尺对急尖长苞冷杉幼苗的苗高、地径进行统计,用分析天平分别测量样品鲜质量、饱和质量。
1.3.2 根系活力测定
急尖长苞冷杉幼苗根系活力测定采用氯化三苯基四氮䂳(TTC)法。将各海拔梯度样本根系混匀,各称取0.1 g样品,浸没于混合液中,37℃保温3 h后,加入1 mol/L硫酸2 mL终止反应,各取5 mL样品液置于刻度试管中,加入5 mL乙酸乙酯和少量连二亚硫酸钠(Na2S2O4),保证各管中量一致,充分振荡提取红色有色液层,以空白作参比,用分光光度计测定在485 nm处的吸光度值,测定根系活力。
1.3.3 叶绿素含量测定
用丙酮提取急尖长苞冷杉幼苗叶片,采用紫外分光光度计(UV5200)测定其叶绿素含量。
1.3.4 抗逆性指标测定
可溶性还原糖含量采用蒽酮法测定,可溶性蛋白质含量采用考马斯亮蓝染色法测定,丙二醛含量采用双组分分光光度法测定,超氧化物歧化酶活性采用氮蓝四唑(NBT)法测定。
1.3.5 数据处理
采用Excel 2013、R3.3.1、SPSS 21.0等软件对试验数据和各项指标进行分析、处理,以及表格、图表的绘制。
2 结果与分析
2.1 不同海拔梯度的急尖长苞冷杉幼苗基本生长指标差异
在不同海拔梯度下急尖长苞冷杉幼苗的苗高存在显著差异(图1)。其中,在3 900~4 400 m海拔范围内,苗高呈现显著下降趋势,但在3 900和4 100 m处有明显峰值,且在海拔3 900 m处冷杉幼苗苗高达到(81.0±1.3)mm,较海拔4 400 m处高出66.4%。不同海拔梯度的地径差异不显著,基本维持在1.4~2.4 mm之间,但在海拔4 400 m处跌落至最低值(图2)。
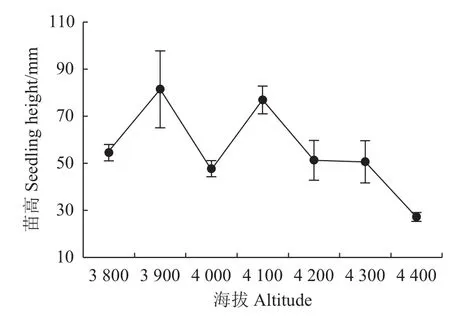
图1 不同海拔的急尖长苞冷杉幼苗苗高变化Fig.1 Height changes of Abies georgei var.smithii seedlings at different altitudes

图2 不同海拔的急尖长苞冷杉幼苗地径变化Fig.2 Ground diameter changes of Abies georgei var.smithii seedlings at different altitudes
2.2 不同海拔梯度的急尖长苞冷杉幼苗叶绿素含量差异
叶片是植物与外界环境接触面积最大的器官,在特定的环境中,植物叶片的形态特征、生理生化指标都会随之发生相应的适应性变化,这些变化在植物对碳的同化、水分蒸腾和能量平衡方面发挥着重要作用[9]。在本研究中,不同海拔梯度的急尖长苞冷杉幼苗叶片的叶绿素含量差异显著,且在不同海拔梯度下叶绿素a、叶绿素b和叶绿素总量均呈现出一致的变化趋势(图3)。在海拔3 900、4 100和4 300 m处冷杉幼苗叶片的叶绿素含量出现3次峰值,并在4300m处叶绿素总量达到最大值(12.76mg/L),比海拔4 000 m处超出51.1%。叶绿素含量是光反应进行的基础,长期以来叶绿素含量被作为衡量叶片光能吸收和利用能力的指标[17]。冷杉幼苗分别在海拔3 900、4 100和4 300 m处达到了峰值,根据样地地形分析,认为在海拔4 300 m处已接近林线范围,高大乔木分布稀疏,光照强度高,而在海拔4 400 m处为急尖长苞冷杉树线范围,极端低温,土壤瘠薄,光合效率低,幼苗生长状况差。
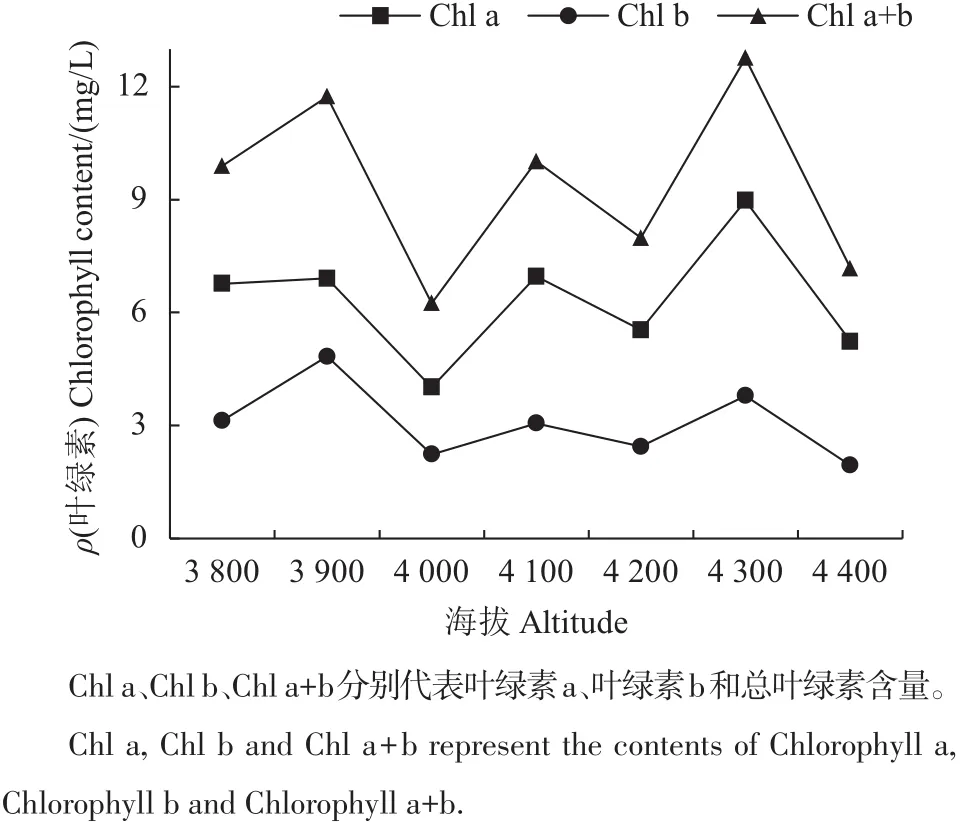
图3 不同海拔的急尖长苞冷杉幼苗叶片叶绿素含量变化Fig.3 Chlorophyll content changes of Abies georgei var.smithii seedlings at different altitudes
2.3 不同海拔梯度的急尖长苞冷杉幼苗根系活力差异
根系是植物活跃的吸收、合成器官,大量研究证实,根的生长状况、形态分布和活跃水平直接或间接地影响地上部分的营养分配及生长状况,根系活力是表征根系吸收养分功能的主要指标。相关性分析发现,急尖长苞冷杉幼苗根系活力与海拔呈显著负相关。色季拉山不同海拔急尖长苞冷杉幼苗根系活力随海拔上升总体呈现下降趋势,并在海拔3 800 m处达到峰值[7.86 μg/(g·h)],比海拔4 400 m处的最低值高出32.9%(图4)。这说明冷杉幼苗根系在海拔3 800 m处活力最强。
2.4 不同海拔梯度的急尖长苞冷杉幼苗丙二醛含量差异
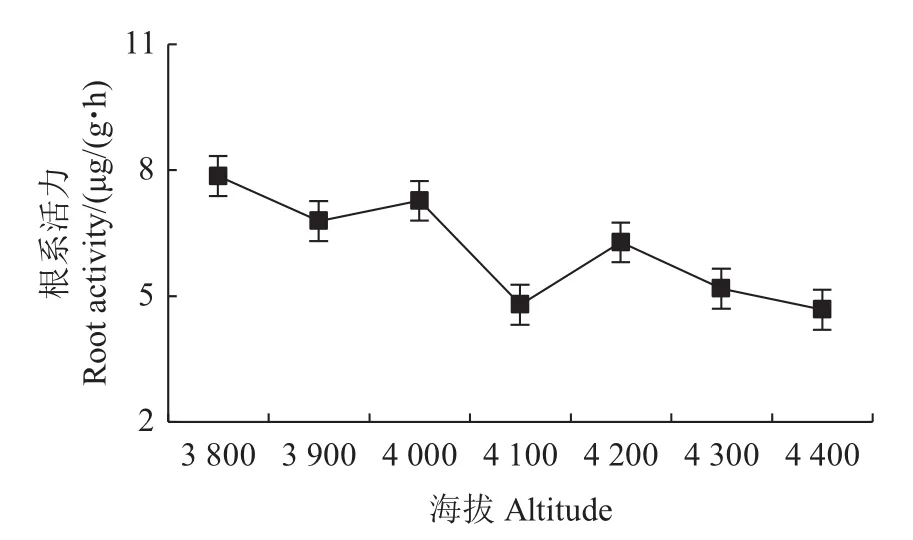
图4 不同海拔的急尖长苞冷杉幼苗根系活力变化Fig.4 Root activity changes of Abies georgei var.smithii seedlings at different altitudes
植物暴露在有氧环境中或多或少都会受到超氧化物、过氧化氢等活性氧的影响,这会引起膜脂类过氧化和蛋白质变性从而导致膜脂类的损伤。细胞膜通透性变化能反映细胞的损失程度。丙二醛(MDA)是膜脂过氧化作用的主要产物,它的产生会加剧膜的损伤,因此丙二醛含量可以有效反映膜脂过氧化程度。由图5可知,色季拉山不同海拔急尖长苞冷杉幼苗叶片和根系的丙二醛含量存在显著差异。其中:根系丙二醛含量始终高于叶片含量,且随海拔的升高而升高;在海拔4 400 m处,急尖长苞冷杉幼苗根系的丙二醛含量达到峰值(5.05 μmol/L),比海拔3 800 m处提高了25.4%。
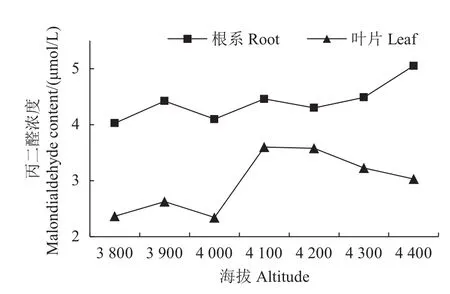
图5 不同海拔的急尖长苞冷杉幼苗丙二醛浓度变化Fig.5 Malondiadehyde content changes of Abies georgei var.smithiiseedlings at different altitudes
2.5 不同海拔梯度的急尖长苞冷杉幼苗可溶性蛋白质含量差异
可溶性蛋白质大多是参与各种代谢的酶类物质,在受到环境胁迫时会发生一定的变化,因此可溶性蛋白质含量是衡量植物抗逆性的重要指标[18]。由图6可知:色季拉山不同海拔梯度的急尖长苞冷杉幼苗叶片、根系的可溶性蛋白质含量存在极显著差异,可溶性蛋白质含量基本随海拔上升而升高;在色季拉山海拔3 800~4 100 m范围内,可溶性蛋白质含量基本稳定,从海拔4 200 m之后,可溶性蛋白质含量显著上升,在海拔4 300 m处叶片可溶性蛋白质达到最大值(13.30 mg/g)。在海拔4 400 m处,冷杉幼苗叶片、根系的可溶性蛋白质含量相比于3 800 m处分别提高了194%和820%,说明在海拔4 300和4 400 m处急尖长苞冷杉幼苗受到了较强的环境胁迫,导致可溶性蛋白质含量显著升高。
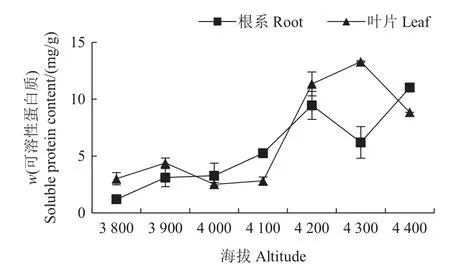
图6 不同海拔的急尖长苞冷杉幼苗可溶性蛋白质含量变化Fig.6 Soluble protein content changes of Abies georgei var.smithii seedlings at different altitudes
2.6 不同海拔梯度的急尖长苞冷杉幼苗可溶性还原糖含量差异
可溶性还原糖是一种重要的渗透调节物质,在干旱胁迫下,植物体内的可溶性糖含量在一定程度上能反映植物对不良环境的适应能力[19]。由图7可知:色季拉山不同海拔急尖长苞冷杉幼苗叶片的可溶性还原糖差异不显著,而根系之间的可溶性还原糖含量差异显著;在海拔3 800~4 100 m范围内,急尖长苞冷杉幼苗根系的可溶性还原糖含量持续降低,但在4 100~4 300 m范围内显著升高,并在海拔4 300 m处达到峰值19.13%,在海拔4 400 m处急剧下降到15.80%,波动明显。
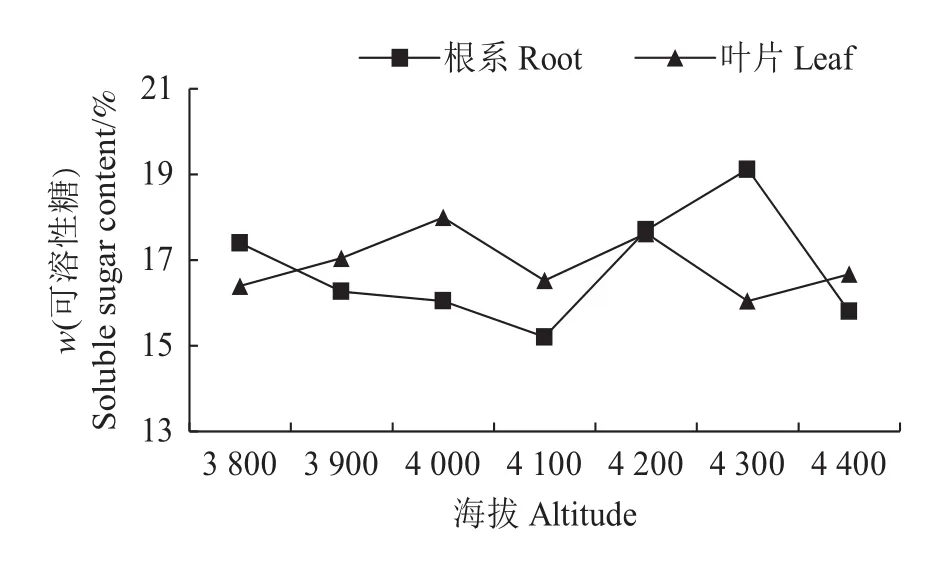
图7 不同海拔的急尖长苞冷杉幼苗可溶性还原糖含量变化Fig.7 Soluble sugar content changes of Abies georgei var.smithii seedlings at different altitudes
2.7 不同海拔梯度的急尖长苞冷杉幼苗超氧化物歧化酶活性差异
超氧化物歧化酶(superoxide dismutase,SOD)是一种广泛存在的生物体金属酶[20],它通过与过氧化物酶、过氧化氢酶等发生协同作用,来防御活性氧或其他过氧化物自由基对细胞膜系统的伤害。在正常情况下,SOD可清除植物体内不断产生的超氧自由基,使植物体内SOD活性和超氧自由基含量达成一定的平衡关系[21]。SOD活性高低可反映细胞膜透性变化和膜脂过氧化产物的含量变化。由图8可知:色季拉山急尖长苞冷杉幼苗根系SOD活性基本随海拔的升高而降低,但在海拔3 800和3 900 m之间呈先升高后降低的趋势,在海拔4 300~4 400 m处急剧下降,达到最低值;在色季拉山海拔3 900 m处,冷杉幼苗SOD活性达到最高值[54.35 μg/(g·h)],在海拔4 400 m处由于外界环境恶劣,已经超过冷杉幼苗承受范围,冷杉幼苗SOD活性急剧减弱,无法有效抵御逆境带来的损害。
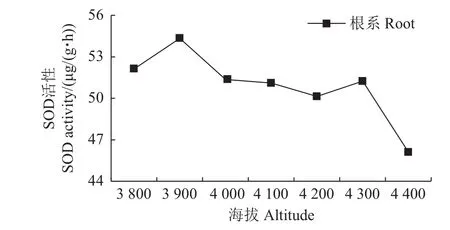
图8 不同海拔的急尖长苞冷杉幼苗根系超氧化物歧化酶(SOD)活性变化Fig.8 Root superoxide dismutase(SOD)activity of Abies georgei var.smithii seedlings at different altitudes
2.8 不同海拔梯度的急尖长苞冷杉幼苗相关性分析
采用SPSS 21.0软件对色季拉山不同海拔梯度急尖长苞冷杉幼苗各项基本指标进行相关性分析,结果如表2、表3所示。急尖长苞冷杉幼苗的苗高、根系活力和海拔均呈显著负相关,幼苗叶片、根系的可溶性蛋白质含量与海拔呈极显著正相关,根系的丙二醛含量与海拔呈显著正相关,地径、叶绿素含量、可溶性还原糖含量、超氧化物歧化酶活性与海拔间无显著相关性。

表2 不同海拔的急尖长苞冷杉幼苗基本指标的相关性分析Table 2 Correlation analysis of basic characteristics of Abies georgei var.smithii seedlings at different altitudes
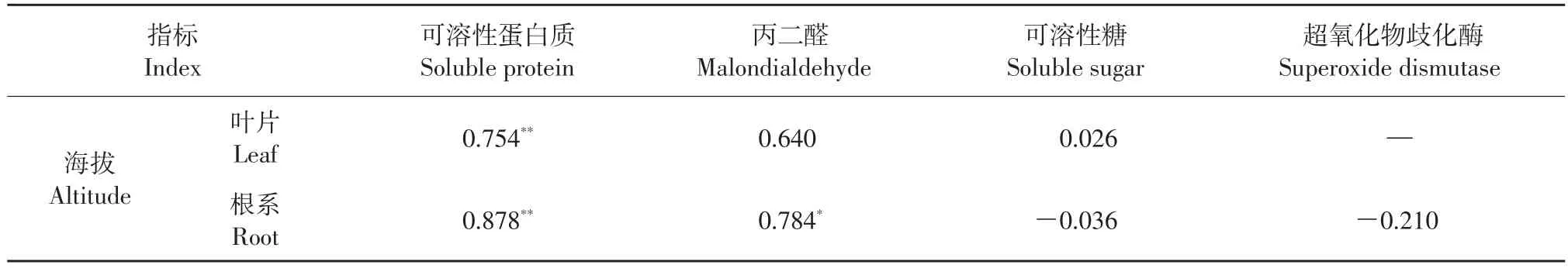
表3 不同海拔的急尖长苞冷杉幼苗抗逆性指标的相关性分析Table 3 Correlation analysis of resistant indexes of Abies georgei var.smithii seedlings at different altitudes
3 讨论与结论
青藏高原具有显著区别于其他各地的气候类型和地形因子,其中海拔是重要的地形因子之一。各环境因子在海拔梯度上也表现出梯度性变化,例如气温、气压等随着海拔的升高而降低,但也有诸如极端温度、光照、降雨量等因子出现不规律性变化[9],加之微地形的差异,相同海拔的环境因子也有不同,这对当地植被的生长分布产生了复杂影响,因此急尖长苞冷杉对不同海拔环境的响应也具有多样性。苗高、地径等基本生长指标可以直观体现出急尖长苞冷杉幼苗在不同海拔梯度的基本生长状况。在色季拉山不同海拔梯度上,冷杉幼苗的苗高、地径在海拔3 900、4 100 m处较大,幼苗生长状况良好,光合效率高,而丙二醛、可溶性蛋白质和可溶性还原糖含量在海拔3 900 m处相对较低,说明在海拔3 900 m处环境胁迫和土壤渗透势对冷杉幼苗的影响较小。且在海拔3 800~4 100 m范围内土壤温度、湿度相对较高,腐殖质层可达15~20 cm,土壤含砂石较少,有利于急尖长苞冷杉幼苗根系吸收水分、养分。综上表明,在色季拉山海拔3 900与4 100 m处适宜急尖长苞冷杉幼苗生长。
急尖长苞冷杉在生长过程中会受到各种环境胁迫诸如干旱、水害、热害、冷害、病虫害等的影响,这些环境胁迫都将导致器官细胞的膜脂类过氧化、蛋白质变性等,最终引起植株体内各项生理指标发生规律性变化。对急尖长苞冷杉幼苗的抗逆性指标丙二醛含量、超氧化物歧化酶活性、可溶性蛋白质和可溶性还原糖含量变化分析表明,在海拔3 800~4 100 m范围内,急尖长苞冷杉幼苗各项指标都趋于稳定,幼苗生长良好,环境胁迫并未对冷杉幼苗产生较大影响。在海拔4 200~4 400 m范围内,急尖长苞冷杉幼苗表现出显著的抗逆性指标变化,冷杉幼苗根系丙二醛、可溶性蛋白质含量显著上升,超氧化物歧化酶活性显著降低。结合根系活力指标发现,在海拔4 200~4 400 m范围内,急尖长苞冷杉幼苗遭受环境胁迫影响较大,导致冷杉幼苗根系活力衰弱,苗高、地径等基本生长指标下滑,苗木生长不良。
在海拔4 200~4 400 m范围内,林分向树线过渡,郁闭度急剧下降,灌木、草丛发达,群落结构为杜鹃冷杉林,光照强度高,在海拔4 300 m处冷杉幼苗叶片叶绿素含量达到峰值。但随着海拔升高,温度显著下降,昼夜极端温差超过冷杉幼苗所能承受的临界值,土壤层稀薄、砂石较多,冷杉根系无法从土壤中汲取足够的水分和养分。在海拔4 300和4 400 m处冷杉幼苗的各项生理指标显示,冷杉幼苗丙二醛含量达到峰值,环境胁迫对其影响严重,因此,该海拔范围的冷杉幼苗根系活力不足,苗木生长较差,在海拔4 400 m处表现尤为显著。
综上所述,急尖长苞冷杉幼苗生长受海拔影响显著。在海拔3 800~4 100 m范围内急尖长苞冷杉幼苗生长较好,尤其在3 900 m处冷杉幼苗生长最旺盛;在海拔4 200~4 400 m处,外界环境变化剧烈,冷杉幼苗无法抵御该范围环境胁迫带来的影响,表现为幼苗矮小,活力衰弱,生长缓慢。结合试验地光照、温度、水分、林分条件和微地形等因素分析认为,在色季拉山海拔3 900 m处最适宜急尖长苞冷杉幼苗生长。
本研究由于受西藏特殊地理环境影响,交通运输、采样分析不便,在同一海拔只设置了1块典型样地,无法完全排除试验地差异带来的误差。在今后的研究中,需进一步加大采样密度和广度,缩小海拔跨度,以更加精确地分析急尖长苞冷杉幼苗对藏东南色季拉山海拔梯度的响应。
[1] 刘金环,曾德慧,DON K L.科尔沁沙地东南部地区主要植物叶片性状及其相互关系.生态学杂志,2006,25(8):921-925.LIU J H,ZENG D H,DON K L.Leaf traits and their interrelationships of main plant species in southeast Horqin sandy land.Chinese Journal of Ecology,2006,25(8):921-925.(in Chinese with English abstract)
[2] VENDRAMINI F,DÍAZ S,GURVICH D E,et al.Leaf traits as indicators of resource-use strategy in floras with succulent species.New Phytologist,2002,154(1):147-157.
[3] VORONIN P Y,IVANOVA L A,RONZHINA D A,et al.Structural and functional changes in the leaves of plants from steppe communities as affected by aridization of the Eurasian climate.Russian Journal of Plant Physiology,2003,50(5):604-611.
[4] 荣秀连,王波,宋采博,等.土壤某些物理性状对植物根系生长的影响.草业与畜牧,2009(10):1-3.RONG X L,WANG B,SONG C B,et al.Effects of some physical properties of soil on plant root growth.Prataculture&Animal Husbandry,2009(10):1-3.(in Chinese)
[5] 未晓巍,吕杰,武慧,等.植物根系研究进展.北方园艺,2012(18):206-209.WEI X W,LÜ J,WU H,et al.Research advances of plant roots.Northern Horticulture,2012(18):206-209.(in Chinese)
[6] JORDAN D N,SMITH W K.Radiation frost susceptibility and the association between sky exposure and leaf size.Oecologia,1995,103(1):43-48.
[7] 席本野,王烨,贾黎明,等.宽窄行栽植模式下三倍体毛白杨根系分布特征及其与根系吸水的关系.生态学报,2011,31(1):47-57.XI B Y,WANG Y,JIA L M,et al.Property of root distribution of triploid Populus tomentosa and its relation to root water uptake under the wide-and-narrow row spacing scheme.Acta Ecologica Sinica,2011,31(1):47-57.(in Chinese with English abstract)
[8] 韩发,贲桂英,师生波.青藏高原不同海拔矮嵩草抗逆性的比较研究.生态学报,1998,18(6):654-659.HAN F,BEN G Y,SHI S B.Comparative study on the resistance of Kobresia humilis grown at different altitudes in Qinghai-Xizang Plateau.Acta Ecologica Sinica,1998,18(6):654-659.(in Chinese with English abstract)
[9] 祁建,马克明,张育新.辽东栎(Quercus liaotungensis)叶特性沿海拔梯度的变化及其环境解释.生态学报,2007,27(3):930-937.QI J,MA K M,ZHANG Y X.The altitudinal variation of leaf traits of Quercus liaotungensis and associated environmental explanations.Acta Ecologica Sinica,2007,27(3):930-937.(in Chinese with English abstract)
[10]KULLMAN L.Rapid recent range-margin rise of tree and shrub species in the Swedish Scandes.Journal of Ecology,2002,90(1):68-77.
[11]石培礼.亚高山林线生态交错带的植被生态学研究.北京:中国科学院,1999.SHI P L.A study on vegetation ecology of subalpine timberline ecotone.Beijing:Chinese Academy of Sciences,1999.(in Chinese)
[12]WANG X P,ZHANG L,FANG J Y.Geographical differences in alpine timberline and its climatic interpretation in China.Acta Geographica Sinica,2004,59(6):871-879.
[13]何吉成,罗天祥,徐雨晴.藏东南色季拉山急尖长苞冷杉(Abies georgei var.smithii)林线的生态气候特征.生态学报,2009,29(1):37-46.HE J C,LUO T X,XU Y Q.Characteristics of eco-climate at Smith fir timberline in the Sergyemla Mountains,Southeast Tibetan Plateau.Acta Ecologica Sinica,2009,29(1):37-46.(in Chinese with English abstract)
[14]罗大庆,张晓娟,任德智.藏东南色季拉山冷杉林林隙与非林隙小气候比较.北京林业大学学报,2014,36(6):48-53.LUO D Q,ZHANG X J,REN D Z.Comparative research on microclimate between forest gaps and non-gaps of Smith fir forests in the Sejila Mountains,southeastern Tibet.Journal of Beijing Forestry University,2014,36(6):48-53.(in Chinese with English abstract)
[15]王景升,王文波,普琼.西藏色季拉山主要林型土壤的水文功能.东北林业大学学报,2005,33(2):48-51.WANG J S,WANG W B,PU Q.Soil hydrological functions of the main forests on Sejila Hills in Tibet.Journal of Northeast Forestry University,2005,33(2):48-51.(in Chinese with English abstract)
[16]杨小林,崔国发,任青山,等.西藏色季拉山林线植物群落多样性格局及林线的稳定性.北京林业大学学报,2008,30(1):14-20.YANG X L,CUI G F,REN Q S,et al.Plant diversity pattern and stability of the timberline community on Sejila Mountain in Tibet,western China.Journal of Beijing Forestry University,2008,30(1):14-20.(in Chinese with English abstract)
[17]LIN Z H,CHEN L S,CHEN R B,et al.CO2assimilation,ribulose-1,5-bisphosphate carboxylase/oxygenase,carbohydrates and photosynthetic electron transport probed by the JIP-test,of tea leaves in response to phosphorus supply.BMC Plant Biology,2009,9:43.
[18]史玉炜,王燕凌,李文兵,等.水分胁迫对刚毛柽柳可溶性蛋白、可溶性糖和脯氨酸含量变化的影响.新疆农业大学学报,2007,30(2):5-8.SHI Y W,WANG Y L,LI W B,et al.Effects of water stress on soluble protein,soluble sugar and proline content in Tamarix hispida.Journal of Xinjiang Agricultural University,2007,30(2):5-8.(in Chinese with English abstract)
[19]吉增宝,王进鑫,李继文,等.不同季节干旱及复水对刺槐幼苗可溶性糖含量的影响.西北植物学报,2009,29(7):1358-1363.JI Z B,WANG J X,LI J W,et al.Dynamic changes of soluble sugar in the seedlings of Robinia pseudoacacia under drought stress and rewatering in different seasons.Acta Botanica Boreali-Occidentalia Sinica,2009,29(7):1358-1363.(in Chinese with English abstract)
[20]马伟荣,童军茂,单春会.超氧化物歧化酶(SOD)的特征及在植物抗逆性方面的研究进展.食品工业,2013,34(9):154-158.MA W R,TONG J M,SHAN C H.The characteristics of superoxide dismutase and its research in plant resistance.The Food Industry,2013,34(9):154-158.(in Chinese with English abstract)
[21]窦俊辉,喻树迅,范术丽,等.SOD与植物胁迫抗性.分子植物育种,2010,8(2):359-364.DOU J H,YU S X,FAN S L,et al.SOD and plant stress resistance.Molecular Plant Breeding,2010,8(2):359-364.(in Chinese with English abstract)
Basic characteristics and physiological index changes of Abies georgei var.smithii seedlings at different altitudes of Shergyla Mountain.Journal of Zhejiang University(Agric.&Life Sci.),2017,43(5):623-630
LIU Jiming1,2,XIN Fumei1,2*,YANG Xiaolin2,ZHAO Kentian2,JIA Liming1
(1.College of Forestry,Beijing Forestry University,Beijing 100083,China;2.College of Resource and Environment,Xizang Agriculture and Animal Husbandry College,Linzhi 860000,Xizang,China)
Shergyla Mountain;altitude;Abies georgei var.smithii seedlings;physiological indexes
S 718.43
A
10.3785/j.issn.1008-9209.2016.08.242
Summary The relationship between plants and environment is mainly reflected in the life history,morphology,phenology,physiology of plants.Different kinds of plants have different adaptation strategies.The southeastern of Qinghai-Tibet Plateau is among the highest forest line distribution areas and one of the most obvious climate change areas in the world.Understanding ecological and climatic indicators of the forest line in this area can offer a foundation for analyzing the possible changes in the future climate scenario.By means of research on the basic characteristics and physiological indicators of Abies georgei var.smithii seedlings on Mt.Shergyla in southeastern Tibet,we hope to discover the response of Abies georgei var.smithii seedlings to the environment with different altitude gradients,and provide basic evidence for further explaining the response of Abies georgei var.smithii seedlings to altitude.
In this paper,we collected 350 seedlings of 7-8 years with great growth condition in the range of 3 800-4 400 m above sea level in April 2014.We measured their basic growth indexes,root activity,chlorophyll content,etc.,and employed Excel 2013,R3.3.1,SPSS 21.0 and other software to analyze the data.
国家自然科学基金(31460192);西藏高原林业生态工程重点实验室建设项目;西藏农牧学院生态学学科高原生态恢复学术团队项目。
辛福梅(http://orcid.org/0000-0001-6470-5200),E-mail:xzxinfumei@163.com
(First author):刘济铭(http://orcid.org/0000-0003-2926-8038),E-mail:18289143108@163.com
2016-08-24;接受日期(Accepted):2017-09-11
The results showed that:1)The optimal growth of Abies georgei var.smithii seedlings was observed at the altitude of 3 900 m.The basic characteristics and chlorophyll content were significantly higher than the other altitudes.2)At the altitude of 3 800 m to 4 100 m,the soluble protein content,soluble sugar content and root activity of Abies georgei var.smithii seedlings were basically stable,indicating that the environmental stress had no significant effect on the growth of Abies georgei var.smithii seedlings.3)From the altitude of 4 200 m in the Mt.Shergyla,transition began with the forest line and tree line,and the external environment at the altitude of 4 400 m was harsh.The indexes such as seedling height,root activity and chlorophyll content declined sharply which were greatly affected by environmental stress.
In sum,the growth of Abies georgei var.smithii seedlings is largely affected by the altitude of Mt.Shergyla.The seedlings could not resist the effects of environmental stress in the range of the forest line,which resulted in short seedlings with weak energy and slow growth.
猜你喜欢
数学物理学报(2022年5期)2022-10-09
军事文摘(2022年14期)2022-08-26
散文诗(2021年22期)2022-01-12
林业与生态(2019年9期)2019-10-07
华东师范大学学报(自然科学版)(2019年3期)2019-06-24
环球时报(2019-05-23)2019-05-23
花火A(2019年1期)2019-03-22
电子制作(2018年1期)2018-04-04
现代养生·上半月(2017年7期)2017-07-07
发明与创新·大科技(2017年6期)2017-06-20
