
黑龙江省水稻品种抗性与稻瘟病病菌致病性年际变化趋势分析
2017-11-30 21:13马军韬张国民张丽艳邓凌韦王永力王英宫秀杰任洋
江苏农业科学 2017年20期
马军韬+张国民+张丽艳+邓凌韦+王永力+王英+宫秀杰+任洋

摘要:为分析黑龙江省水稻品种抗性及稻瘟病病菌致病性在年际间的波动及变异趋势,以8个水稻品种及24个抗瘟基因为试验材料,以180株稻瘟病病菌为选择压力,应用喷雾接种方式进行试验。结果表明,以2009—2011年的病菌为选择压力,水稻品种的抗性频率分别介于43.33%~93.33%、28.33%~73.33%、25.00%~78.33%之间,龙盾104抗性较好;联合抗病性方式下,各水稻组合的联合抗病性系数平均值分别为0.57、0.33、0.35,绥粳4与龙盾104组合抗性较好。以2009—2011年的病菌为选择压力,各基因的抗性频率分别介于6.67%~95.00%、5.00%~8667%、6.67%~91.67%之间,Pi-z5、Pi-9基因抗性较好;聚合Pi-z5与Pi-9基因条件下,品种抗性频率平均值分别为10000%、90.83%、96.46%。2009—2011年病菌的致病性相似系数分别介于0.35~0.97、0.19~1.00、0.20~1.00之间,致病率分别介于8.33%~87.50%、8.33%~95.83%、12.50%~95.83%之间,无毒基因Pi-9和Pi-z5出现的频率较高且相对稳定。
关键词:黑龙江省;水稻品种;抗瘟基因;稻瘟病菌;致病性;变化趋势
中图分类号: S435.111.4+1 文献标志码: A 文章编号:1002-1302(2017)20-0109-05
黑龙江省是我国水稻生产大省,同时也是稻瘟病为害的重灾区,常年减产在5%以上[1-2]。目前,当地生产上对该病害的控制以化学防控为主,农业防控为辅,但前者污染环境,后者技术水平要求高,效果均不理想。进一步分析发现,水稻种质由于自身抗瘟基因的存在,对稻瘟病具有较高的抗性权重,应用潜力较大。但是,稻瘟病病菌受内在遗传机制及外界环境影响,致病性存在分化,对不同类型种质侵染力差别显著。因此,要想实现水稻种质的生态控病,首要条件是明确稻瘟病病菌致病性的分化情况,获得水稻种质抗性信息,然后通过合理布局或抗性育种等生态方式实现病害高效控制。
目前,国内外相关研究较多。马辉刚等以江西省67个稻瘟病病菌菌株为选择压力,对35个水稻品种的抗性进行了评价,明确了两优培九的高抗源地位;应用联合抗病性方式,明确了两优培九与汕优晚3的优势组合地位[3]。张国民等以12个日本鉴别品种和24个水稻单基因系为鑒别体系,对黑龙江省178个稻瘟病病菌菌株的致病性进行了分析,将其划分为104个生理小种,明确了Pi-9的高抗源地位,同时获得了垦稻10、上育397等品种的抗性改良信息[4]。杨秀娟等以41个已知抗性基因的水稻品种为鉴别体系,对福建省87个稻瘟病病菌菌株的致病性进行分析,明确了无毒基因AvrPid-2、AvrPik的高频率地位[5]。此外,张亚玲等也相继进行了类似研究[6-8],对科研及生产起到了一定的促进作用。但是,多数研究由于缺乏种质抗性或病菌致病性在年际间变化及波动趋势方面的分析,从而难以明确种质抗性或病菌致病性的未来发展趋势,不利于抗性布局及育种策略的适时调整,有待改进。
本研究以黑龙江省主栽水稻品种和单基因系作为水稻种质,以连续3年的稻瘟病病菌菌株作为选择压力,在获得水稻种质抗性及稻瘟病病菌致病性准确的信息前提下,明确其年际间变异及波动趋势,为相关的持续研究提供基础信息,同时,应用联合抗病性分析及基因聚合分析的方式为抗性布局及抗瘟育种工作提供优化方案,从而生态控制病害。
1 材料与方法
1.1 试验材料
8个水稻品种分别为松粳9、松粳6、龙稻7、垦稻12、龙盾104、绥粳4、龙稻4、空育131,均由黑龙江省农业科学院耕作栽培研究所提供。
24个水稻单基因系各含有1个已知抗瘟基因,感病对照品种为丽江新团黑谷,不含任何抗瘟基因,均由黑龙江省农业科学院耕作栽培研究所提供。
稻瘟病病菌:总计180株单孢菌株,来源于黑龙江省第一、第二、第三积温带的水稻品种。其中,2009年在24个水稻品种上分离获得60株单孢菌株,2010年在21个水稻品种上分离获得60株单孢菌株,2011年在22个水稻品种上分离获得60株单孢菌株,均由黑龙江省农业科学院耕作栽培研究所分离并保存。
培养基:菌株分离与活化使用燕麦片番茄汁培养基,配方为30 g/L燕麦片、150 mL/L番茄汁、17~20 g/L 琼脂;菌株产孢使用高粱粒培养基,将约200 g高粱粒煮熟后置于三角瓶中,121 ℃湿热灭菌30 min后备用。
1.2 试验方法
1.2.1 稻瘟病病菌单孢子的分离及培养 将病叶用无菌水清洗后,浸泡4~6 h,放入恒温箱培养至产生霉层。镜检挑取单个分生孢子点接于燕麦片番茄汁培养基中,25 ℃培养3 d,将菌丝块挑入装有高粱粒培养基的三角瓶中,25 ℃培养20~25 d,待高粱粒表面长满菌丝后,无菌水洗脱菌丝,将高粱粒平铺于已灭菌铁托盘中,盖上无菌纱布,待高粱粒表面形成大量分生孢子后,用含0.025%吐温-20的自来水洗脱孢子,采用血球计数板计数,将孢子悬浮液浓度调至5×105 个/mL,供接种使用[9]。
1.2.2 水稻品种抗性分析方法 水稻品种抗性分析通过幼苗3叶1心期喷雾接种方式进行。幼苗以草碳土、黑土为基质,用育秧盘育苗,个体保苗15株,3次重复。待幼苗长至3叶1心期时,接种孢子悬浮液,剂量为100 mL/盘,然后在 26 ℃、相对湿度为100%条件下暗培养30 h,光照培养96~144 h,待发病完全后按表1中的标准调查发病情况[10]。调查后,按Hayashi等的标准[11],将0~3级归为抗病反应型,4~5级归为感病反应型。各重复间接种结果若抗、感病表现不一致,则按感病表现进行统计。
用抗性频率表示水稻品种抗性强弱,抗性频率=(非致病病菌菌株数/总菌株数)×100%,用联合抗病性[3,12]评价水稻品种搭配种植后抗性效果,它包括联合抗病性系数和联合致病性系数2个指标,以水稻品种组合既同时高抗稻瘟病病菌(联合抗病性系数最大)又不会同时被稻瘟病病菌侵染(联合致病性系数最小)为优劣评价标准,联合抗病性系数=水稻品种组合共同非致病病菌菌株数/总菌株数;联合致病性系数=水稻品种组合共同致病病菌菌株数/总菌株数。endprint
1.2.3 水稻抗瘟基因抗性分析方法 水稻抗瘟基因抗性分析通过幼苗3叶1心期喷雾接种方式进行,具体方法同“1.2.2”节。用聚合后的抗性频率表示抗瘟基因与水稻品种聚合后可能获得的最佳抗性,通过常规杂交技术完成,聚合后抗性频率=[(总菌株数-聚合后共同致病的菌株数)/总菌株数]×100%。
1.2.4 稻瘟病病菌致病性分析方法 稻瘟病病菌致病性分析通过幼苗3叶1心期喷雾接种方式进行,具体方法同“1.2.2”节。
用致病率表示稻瘟病病菌对水稻品种或抗瘟基因的致病力强弱,致病率=(致病菌株数/总菌株数)×100%。
用稻瘟病病菌致病性聚类分析方式评价菌株间致病性的分化情况,菌株间致病性相似系数差值越大,表示其致病性分化越剧烈。将水稻品种与稻瘟病病菌的反应型作为性状,抗病反应型记为1,感病反应型记为0,构成二态性状。相似系数=1-相异系数。
1.3 数据分析
应用Excel 2010对数据进行统计与计算,利用DPS v7.05分析软件,以不加权算术平均法(UMPGA)完成分析。
2 结果与分析
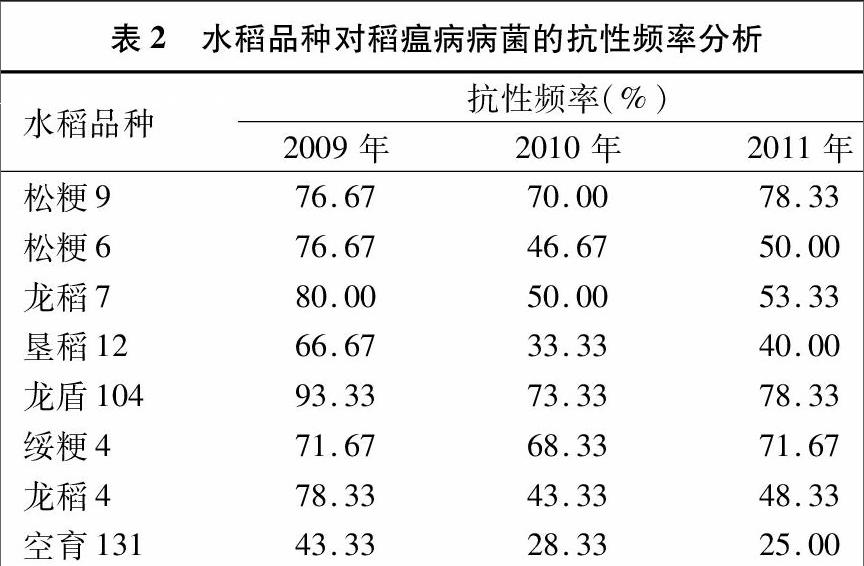
2.1 水稻品种对不同年际稻瘟病病菌抗性的分析
如表2所示,各水稻品种的抗性在稳定的前提下,年际间存在较大波动。以2009—2011年的病菌为选择压力,水稻品种的抗性频率分别介于43.33%~93.33%、28.33%~7333%、25.00%~78.33%之间,平均值分别为73.33%、5167%、5562%。其中,空育131的抗性在年际间呈持续下降的趋势,其他7个品种的抗性在年际间均呈先下降后上升的趋势,龙盾104与空育131一直分别为各年份抗性最好与最差的水稻品种。2011年与2009年相比,松粳9的抗性略有上升,上升了2.17%,其他7个品种的抗性均呈持平或下降趋势,龙稻4的抗性降幅最大,为38.30%。
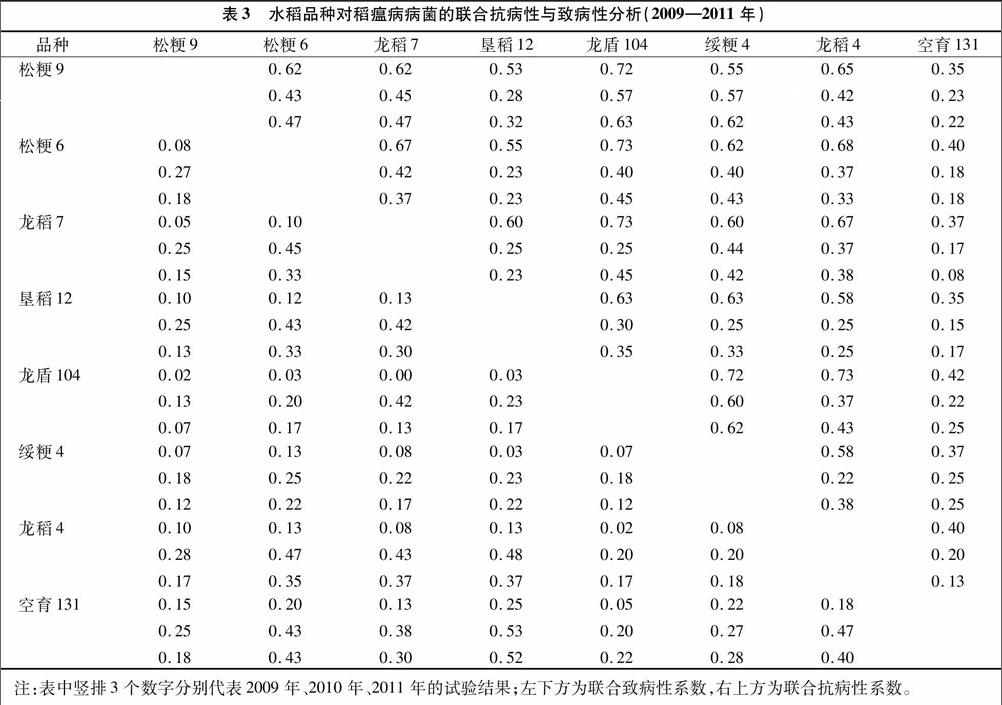
由表3可知,以2009—2011年病菌为选择压力,各水稻组合的联合抗病性系数均值分别为0.57、0.33、0.35,联合致病性系数均值分别为0.10、0.31、0.24。其中,各年际间相对较好的组合分别为龙稻7与龙盾104、龙稻4与龙盾104、绥粳4与龙盾104、松粳9与龙盾104。综合比较可知,绥粳4与龙盾104为联合抗病性较好、年际间波动较小的水稻组合,可以广泛推广应用。
2.2 抗瘟基因对不同年际稻瘟病病菌抗性分析
由表4可知,24个抗瘟基因的抗性在年际间存在较大的波动,以2009—2011年的病菌为选择压力,抗瘟基因的抗性频率分别介于6.67%~95.00%、5.00%~86.67%、6.67%~9167%之间,平均值分别为44.17%、30.42%、3049%。其中,2009—2011年抗性频率≥70%的基因分别有5、3、2个,依次为Pi-9、Pi-z5、Pi-ta2、Pi-12、Pi-20,Pi-zt、Pi-z5、Pi-9,Pi-z5、Pi-9;2009—2011年抗性频率≤30%的基因分别有11、14、14个,Pi-19、Pi-a、Pi-t分别为各年抗性最差的基因。综合评价可知,Pi-z5、Pi-9基因的抗性较好,年际间表现稳定,为优质抗源。依据抗性的变化趋势,可将24个抗瘟基因划分为5种类型:Pi-kp等6个基因的抗性呈持续下降趋势;Pi-k等3个基因的抗性呈持续上升趋势;Pi-zt、Pi-t基因的抗性呈先上升后下降趋势;Pi-ta2基因的抗性呈先下降后持平趋势;Pi-a等12个基因的抗性呈先下降后上升趋势。这间接说明了黑龙江省稻瘟病病菌的致病性变异趋势复杂。此外,2011年与2009年相比,Pi-k基因的抗性升幅最大,上升了109.11%,Pi-ta2基因的抗性降幅最大,下降了81.63%。
依据抗性较好且相对稳定的原则,选取Pi-z5、Pi-9基因通过基因聚合方式对水稻品种进行抗性改良分析。由表5可知,聚合Pi-z5与Pi-9基因组合前提下,水稻品种的抗性提升效应明显。以2009—2011年的病菌为选择压力,各品种的聚合后抗性频率平均值分别为100.00%、90.83%、9646%,其平均值分别比聚合前提高26.67%、39.16%、4084%。聚合单个基因前提下,以2009—2011年的病菌为选择压力,各年份抗性聚合的首选基因分别是Pi-9、Pi-z5、Pi-z5,龙盾104、龙盾104、绥粳4分别为各年份聚合后抗性頻率较高的品种,依次为100.00%、95.00%、98.33%。此外,空育131一直是聚合后抗性频率提升幅度最大的品种。
2.3 不同年际稻瘟病病菌致病性分析
黑龙江省稻瘟病病菌致病性在年际间存在波动,从病菌致病性分化角度分析,2009—2011年病菌的致病性相似系数分别介于0.35~0.97、0.19~1.00、0.20~1.00之间,其差值分别为0.62、0.81、0.80。由图1可知,以相似系数0.36为阈值,2009—2011年病菌分别被划分为2、10、3个致病类群;以相似系数0.51为阈值,病菌分别被划分为6、10、9个致病类群;以更高的相似系数为阈值,不同年际的病菌将被划分为更多的致病类群。从病菌致病力角度分析,2009—2011年,病菌对24个抗瘟基因的致病率分别介于8.33%~87.50%、833%~95.83%、12.50%~95.83%之间,平均值分别为5583%、69.58%、69.51%。此外,2010、2011年开始有相同致病型的菌株群出现,致病率平均值分别为9107%、8472%。从病菌含有无毒基因角度分析,与24个抗瘟基因相对应的无毒基因全部出现,但年际间出现频率不同,2009—2011年单个菌株含有无毒基因的平均数量依次为10.60、730、732个。综合比较,无毒基因Pi-9和APi-z5出现的频率较高且相对稳定,分别占当年总出现频率的8.96%、959%、10.25%与7.70%、10.50%、12.53%。endprint
3 讨论
黑龙江省为粳稻主产省,本研究选用的水稻品种在其域内累计推广面积巨大,后代种质分布广泛;水稻单基因系品种具有粳稻血缘,常规杂交结实率高、后代种质稳定快;稻瘟病病菌采自当地主产区,数量较多,具备年际间准确信息。因此,试验靶标代表性强、可操作性好、应用价值大,在为理论研究奠定基础的同时可通过生态手段解决现实难题。
从水稻品种抗性角度分析,存在2种波动趋势:空育131的抗性呈逐年下降的趋势;其他品种的抗性呈先急剧下降后略有上升的趋势,除松粳9外,其他多数品种的抗性难以恢复到2009年的水平。从品种搭配后的联合抗病性分析,同样呈先下降后上升的趋势,这是因为搭配种植的基础源自水稻品种的自身抗性。除绥粳4与龙盾104等个别组合外,绝大部分组合的联合抗病性系数偏低,联合致病性系数偏高,不适宜推广应用。马军韬等曾以2006年黑龙江省的稻瘟病病菌为选择压力,对13个水稻品种的抗性及联合抗病性进行了分析,其抗性频率介于47.19%~93.82%之间,平均值为7537%,联合抗病性系数、致病性系数平均值分别为0.55、0.15[13],本研究2009年的结果与之基本持平,明显高于2010年和2011年的研究结果。虽然两者选用的水稻品种和病菌存在一定的差别,仍可从侧面说明病菌的致病力已发生一定程度的改变。此外,水稻品种搭配后,自身抗性均强的品种共有抗病性较高,自身抗性均差的品种共有感病风险也较高,说明供试品种间抗性相似度较高,从侧面说明其抗性遗传基础相对狭窄。水稻品种的抗性主要源自本身含有的抗瘟基因,一般可稳定遗传,对相同类型病菌的抵抗能力基本无变化,其抗性之所以有波动,主要是病菌致病力及致病性在年际间发生改变引起的。
从抗瘟基因抗性角度分析,波动趋势多样,主体呈急剧下降又略有回升的趋势,仅少数基因抗性较好且相对稳定。张国民等以2006年黑龙江省的稻瘟病病菌为选择压力,对24个抗瘟基因的抗性进行了分析,发现其抗性频率介于3.93%~9775%之间,平均值为43.48%,Pi-9、Pi-z5、Pi-ta2、Pi-12、Pi-20和Pi-5共6个基因抗性频率超过70%[4],本研究中2009年的结果与之基本持平,明显高于2010年和2011年的研究结果。就抗性较高的优质基因而言,Pi-9、Pi-z5基因抗性较好且持续稳定,可以广泛推广应用;Pi-12、Pi-20基因抗性已不再突出,但仍有一定抗性,可以有限度地推广应用;Pi-ta2、Pi-12基因抗性已急剧下降,应限制应用。上述结论再次说明,病菌的致病力已发生一定程度的改变。从基因聚合角度分析,绥粳4和松粳9通过杂交手段完成聚合后,抗性改良效果明显,后代种质可以大力推广;空育131基因聚合后抗性升幅最大,后代种质可以谨慎推广。雷财林等曾分析了7个抗瘟基因对黑龙江省12个水稻品种的抗性改良效果,认为Pi-9基因对品种的抗性具有极强的改良效果[14],如能进一步导入Pi-z5、Pi-12基因,各品种对稻瘟病可表现免疫,与本研究的结论具有较强的相似度。目前,国内外共发现约101个各类抗瘟基因[15-17],远多于本研究的基因群体,可进一步引进Pi-gm、Pi-40等优质抗源以再次提升当地品种的抗性。
从稻瘟病病菌致病性角度分析,病菌的致病率平均值及致病性相似系数差值均呈先急剧上升后略有下降的趋势,说明病菌的致病力及致病性分化均呈先急剧上升后略有下降的趋势。关于病菌致病性变异,可能主要有2个方面原因:(1)内在遗传机制影响。病菌受自身异核现象、突变和准性重组等细胞学机制影响,菌丝融合现象普遍,田间自然突变率高于10%,完成致病性变异。(2)外在环境条件影响。随着黑龙江省水稻种植面积的增加,很多未经审定的低抗性品系开始占有一定的种植比例,受互作关系影响,加剧了病菌致病力及致病性分化的复杂性与严重程度。2006、2007年黑龙江省稻瘟病连续大发生,产量损失巨大。在随后的2008、2009年,天气条件不利于发病,农民防控措施得当,只个别地区零星发病。自2010年以来,稻瘟病没有再次大发生,防控措施开始松懈,病菌定殖、扩繁能力增强,致病力随之增强;随着种植品种持续增加及区域间引种日趋频繁,病菌致病性分化总体呈加剧趋势是可能的。马军韬等分析了2009—2010年哈尔滨市稻瘟病病菌变异情况后认为,当地病菌的致病力增强、致病性分化加剧[18],本研究的结论与之一致。此外,以稻瘟病病菌含有的无毒基因为现实依据,以Flor的“基因对基因”假说[19]為理论基础,可推导出品种含有抗瘟基因的情况,为抗性合理布局提供理论借鉴。结果显示,龙稻4和龙稻7可能含有Pi-sh基因,垦稻12可能含有Pi-kS基因,但上述结论仅为推测,还须结合标记技术进行验证。
参考文献:
[1]宋成艳. 黑龙江省水稻新品种(系)抗稻瘟病性鉴定及利用[J]. 植物保护,2011,37(4):142-145.
[2]支庚银,张国民,雷材林,等. 黑龙江省2007年水稻稻瘟病生产调研及建议[J]. 黑龙江农业科学,2010(4):68-70.
[3]马辉刚,曹九龙,胡水秀,等. 水稻品种对稻瘟病的抗性分析和利用评价[J]. 江西农业大学学报,2006,28(2):213-216.
[4]张国民,马军韬,肖佳雷,等. 已知抗瘟基因在黑龙江省寒地稻区的评价与利用[J]. 植物病理学报,2011,41(1):72-79.
[5]杨秀娟,阮宏椿,杜宜新,等. 福建省稻瘟病菌致病性及其无毒基因分析[J]. 植物保护学报,2007,34(4):337-342.
[6]张亚玲,王宝玉,台莲梅,等. 黑龙江省稻瘟病菌生理小种对水稻品种致病性分析[J]. 中国植保导刊,2014,34(2):22-26.
[7]赵正洪,周 政,吴伟怀,等. 湖南省稻瘟病菌生理小种的组成及其致病性[J]. 湖南农业大学学报(自然科学版),2014,40(2):173-177.endprint
[8]王世维,郑文静,赵家铭,等. 辽宁省稻瘟病菌无毒基因型鉴定及分析[J]. 中国农业科学,2014,47(3):462-472.
[9]周江鸿,王久林,蒋琬如,等. 我国稻瘟病菌毒力基因的组成及其地理分布[J]. 作物学报,2003,29(5):646-651.
[10]Mackill D J. Inheritance of blast resistance in near-isogenic lines of rice[J]. Phytopathology,1992,82(7):746-749.
[11]Hayashi N,Kobayashi N,Cruz C,et al. Protocols for the sampling of the diseases specimens and evaluation of blast disease in rice[J]. JIRCAS Working Report,2009,63:17-33.
[12]袁 洁,杨学辉,何海永. 部分水稻品种对稻瘟病菌群体的抗病性分析[J]. 种子,2006,25(8):14-16.
[13]马军韬,张国民,辛爱华,等. 黑龙江省水稻品种对稻瘟病的抗性分析及评价利用[J]. 中国农学通报,2008,24(2):332-334.
[14]雷财林,张国民,程治军,等. 黑龙江省稻瘟病菌生理小种毒力基因分析与抗病育种策略[J]. 作物学报,2011,37(1):18-27.
[15]Wang Y,Wang D,Deng X,et al. Molecular mapping of the blast resistance genes Pi2-1 and Pi51(t) in the durably resistant rice ‘Tianjingyeshengdao[J]. Phytopathology,2012,102(8):779-786.
[16]Jiang H C,Feng Y T,Bao L,et al. Improving blast resistance of Jin 23B and its hybrid rice by marker-assisted gene pyramiding[J]. Molecular Breeding,2012,30(4):1679-1688.
[17]Xiao W M,Yang Q Y,Sun D Y,et al. Identification of three major R genes responsible for broad-spectrum blast resistance in an indica rice accession[J]. Molecular Breeding,2015,35(1):1-11.
[18]馬军韬,张国民,辛爱华,等. 哈尔滨地区抗瘟基因抗性分析及水稻品种抗性评价与利用[J]. 植物保护学报,2015,42(2):160-168.
[19]Flor H H. Current status of the gene for gene concept[J]. Annual Review of Phytopathology,1971,9(1):275-296.endprint
猜你喜欢
应用能源技术(2020年11期)2021-01-26
黑龙江省人民政府公报(2017年6期)2017-07-25
黑龙江省人民政府公报(2017年22期)2017-03-26
黑龙江省人民政府公报(2017年21期)2017-03-20
南水北调与水利科技(2016年6期)2017-01-06
湖南畜牧兽医(2016年3期)2016-06-05
兽医导刊(2016年12期)2016-05-17
当代畜禽养殖业(2014年7期)2014-02-27
现代检验医学杂志(2014年1期)2014-02-06
