
养分异质条件下连接和断开结缕草克隆分株抗氧化酶及丙二醛的生理整合
2018-03-09 09:19徐苏男陈忠林张利红
草业科学 2018年2期
徐苏男,李 悦,陈忠林,张利红
(辽宁大学环境学院,辽宁 沈阳 110036)
在自然界中植物生存所需要的必要元素(如光照、水分、养分等)并不是均匀分布的,不断积累的资料显示,这些资源在空间分布上具有斑块性、梯度性,呈现空间异质性[1-2]。生理整合是克隆植物的重要特性,是指物质、资源(如光和同化物、矿质养分和水分等)、信息等通过连接物或间隔子在克隆分株之间进行传输与分享的过程[3-4]。克隆分株可以通过间隔子从相邻分株中获得一定的资源,从而有效缓解资源异质性所致克隆分株资源获得上的差异,同时也可以通过节间长度变化实现其在生境中的选择和迁移[5-7]。因此生理整合使克隆植物在生存、生长及繁殖等方面较非克隆植物更具优势[8],克隆植物对资源空间异质性的适应更强。
近年来,国内外对于克隆植物相连分株间生理整合的研究日益增多,主要集中在外部胁迫(如水、重金属)对克隆分株间水分调控、光合产物分配、表型可塑性的影响及利用同位素示踪法(如15N、32P)研究营养元素在克隆分株间的传输特征[9-12],而对于养分异质条件下克隆植物是否能够通过主动调节生理代谢去适应外部环境,其抗氧化酶活性及丙二醛含量是否存在生理整合,生理整合有何特征的研究比较少见。
结缕草(Zoysiajaponica)为禾本科画眉草亚科结缕草属,主要分布于我国辽东半岛、山东半岛,具有耐旱、耐贫瘠、耐践踏等诸多优点,是一种应用广泛的暖季型草坪草。本研究以克隆植物结缕草为材料,通过分析养分异质条件下结缕草克隆分株叶片抗氧化酶活性及丙二醛含量的变化,揭示克隆植物能否通过主动调节生理代谢实现生理整合及其整合特点,以期更深刻地认识在养分异质环境下克隆植物与环境间及其不同分株间的相互关系和相互作用。
1 材料与方法
1.1 试验材料
供试结缕草选自辽宁大学校园内,同一生长年份、生长相对均匀一致的克隆分株,以保证试验中的结缕草遗传特性和起始状态的一致性。将所选植株栽植在直径13 cm、高10 cm盛有花圃土的花盆中,每盆一个分株作为母株。待每一分株匍匐茎生长到6个复合节时将其顶端植入另一个花盆中,直到长出新的子株,此期间母株和子株在两个花盆中分别成长并保持其匍匐茎相连,约两个月后进行养分异质处理,期间按照结缕草的生长需要进行适度浇水和施肥。供试土壤取自辽宁大学生态实验田,基本理化性状:全氮含量为1.03 g·kg-1,全磷含量为0.4 g·kg-1,全钾含量为13.3 g·kg-1,碱解氮含量为60.2 mg·kg-1,有效磷含量为7.1 mg·kg-1,有效钾含量为68.34 mg·kg-1,pH为6.75。
1.2 试验方法
试验设高、中、低3个养分水平,养分以NH4NO3、NaH2PO4和KCl的混合营养液的形式加入。每个处理中处于不同花盆的母株和子株每周分别加入浓度不同但体积相同(30 mL)的营养液。高养分处理营养液的浓度为7.085 g·L-1NH4NO3+2.063 g·L-1NaH2PO4+2.368 g·L-1KCl,中养分处理营养液的浓度分别为高养分的20%,低养分处理施加相同体积的水。试验期间适时适量浇水,满足植物生长所需水分。在第1次添加营养液前一天将拟进行剪断的分株进行剪断处理,整个养分异质处理持续4周,并于第5周进行各项数据测量。
连接的分株分为5个处理:母株和子株低养分,母株低养分子株中养分,母株低养分子株高养分,母株中养分子株低养分,母株高养分子株低养分(表1)。剪断的分株进行同样处理。每种处理3个重复。
1.3 测量方法
过氧化物酶(POD)活性采用愈创木酚法测定[13];超氧化物歧化酶(SOD)活性采用NBT光化还原法测定[14];过氧化氢酶(CAT)活性采用氧化还原法测定[14];丙二醛(MDA)含量采用硫代巴比妥酸显色法测定[15]。
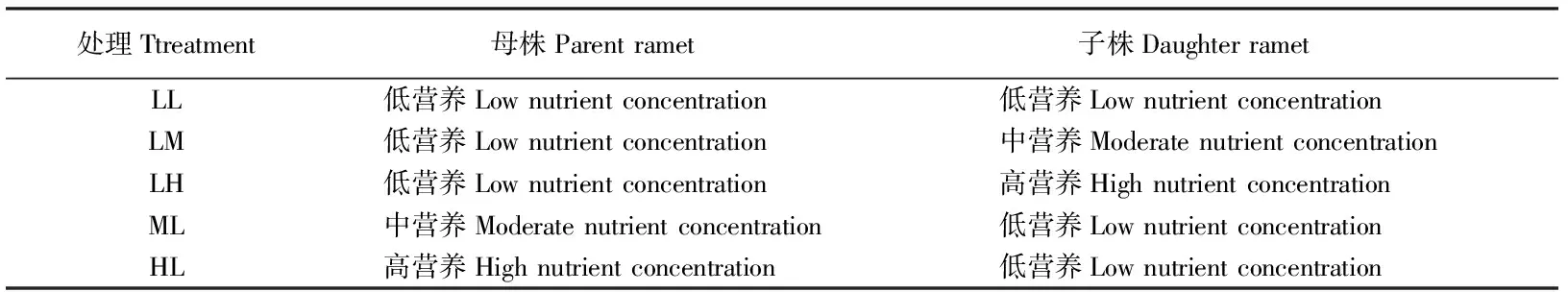
表1 试验设计Table 1 Experimental design
下文中处理编号同表1所列。
LL, LM, LH, ML, and HL are the same as following figures.
1.4 数据统计分析
试验数据统计分析采用SPSS 17.0软件,用ANOVA(analysis of variance,LSD检验法)进行方差分析,采用Excel 2010制图。
2 结果与分析
2.1 养分异质条件下结缕草克隆分株SOD活性的生理整合
养分异质条件下母株和子株相连时,母株SOD活性LL组最低,LM组和LH组比LL组略有升高(图1),相互之间差异不显著(P>0.05),ML和HL组母株SOD活性显著高于其他3组(P<0.05);子株SOD活性总体上随养分升高而上升,LL组最低,LH组最高且与其他各组呈显著差异(P<0.05),ML组和HL组子株SOD活性比LL组分别高38.67%和43.75%,且差异显著(P<0.05)。
养分异质条件下母株和子株断开时,母株SOD活性随养分升高而上升,ML和HL组与其他3组差异显著(P<0.05);子株SOD活性变化与母株类似,养分水平越高SOD活性越大,且不同养分水平之间差异性显著(P<0.05)(图1)。
2.2 养分异质条件下结缕草克隆分株POD活性的生理整合
养分异质条件下母株与子株相连时,母株POD活性以HL组最高,且与其他4个处理组差异显著(P<0.05),ML组母株POD活性稍高于LL组,与中高养分子株相连的LM和LH组母株POD活性甚至低于LL组;子株POD活性随养分水平升高而升高,LL组最低,LH组最高,且LL、LM、LH组之间差异显著(P<0.05),与SOD活性不同,ML和HL组子株POD活性与LL组相比升高不多,相互之间差异不显著(P>0.05)(图2)。
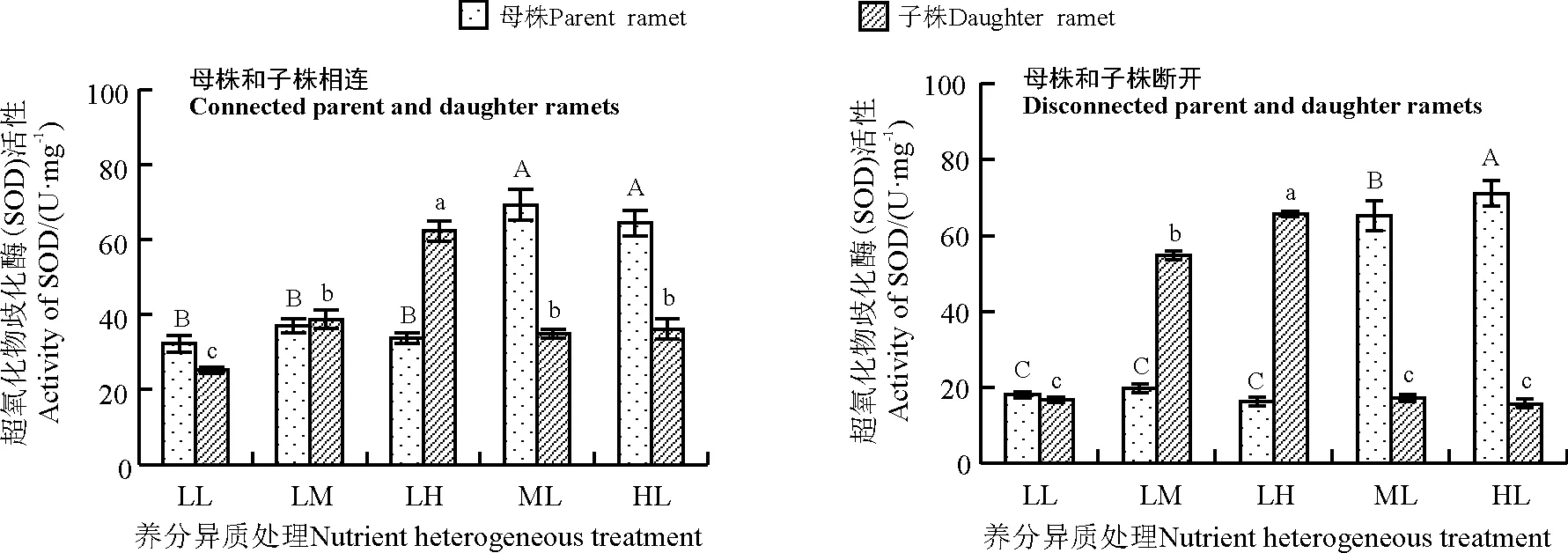
图1 养分异质条件下结缕草克隆分株叶片超氧化物歧化酶(SOD)活性Fig. 1 SOD activity in clonal ramet leaves of Zoysia japonica under nutrient heterogeneity
不同大写字母表示母株不同养分处理间差异显著(P<0.05),不同小写字母表示子株不同养分处理间差异显著(P<0.05),下同。
Different capital(lowercase)letters indicate significant difference among different heterogeneous nutrient for concentrations parent(daughter)ramets at the 0.05 level;similarly for the following figures.

图2 养分异质条件下结缕草克隆分株叶片过氧化物酶(POD)活性Fig. 2 POD activity in clonal ramet leaves of Zoysia japonica under nutrient heterogeneity
养分异质条件下结缕草母株与子株断开时叶片中POD活性显示,养分水平越高,母株POD活性越大,且各营养水平之间差异显著(P<0.05);子株POD活性变化与母株相似,LH组最高,LM组次之,其他3组之间基本一致,LH组与其他各组差异显著(P<0.05)。
2.3 养分异质条件下结缕草克隆分株CAT活性的生理整合
养分异质条件下结缕草母株子株相连时,LL组母株CAT活性最低,虽然ML组和HL组母株CAT活性仍高于LL组,但只有ML组与LL组之间差异显著(P<0.05);LM组和LH组母株CAT活性比LL组都有所提高,但只有LH组与LL组差异显著(P<0.05)(图3)。LL组子株CAT活性最低,LH组CAT活性最高,LM组次之,且相互之间差异显著(P<0.05),子株低养分组之间,ML组与HL组子株CAT活性分别比LL组高64.29%和75.00%,且差异显著(P<0.05)。
养分异质条件下母株子株断开时,母株CAT活性随着养分水平的升高而上升,HL组、ML组分别与其他各组差异显著(P<0.05);子株CAT活性变化与母株一样,养分越高CAT活性越大,LH组、ML组分别与其他各组存在显著差异(P<0.05)(图3)。
2.4 养分异质条件下结缕草克隆分株MDA的生理整合
养分异质条件下结缕草母株子株相连时,母株MDA含量LL组最高,LM组和LH组比LL组分别低0.90%和5.94%,其中LH组与LL组差异显著(P<0.05),随着母株养分的提高,ML组和HL组母株MDA含量进一步下降,且与其他3组差异显著(P<0.05);子株MDA含量也是LL组最高,随着子株养分水平升高,LM组和LH组子株MDA含量逐渐下降,均与LL组存在显著差异(P<0.05), ML和HL组子株MDA含量与LL组相比也有明显下降,且都与LL组差异显著(P<0.05)(图4)。
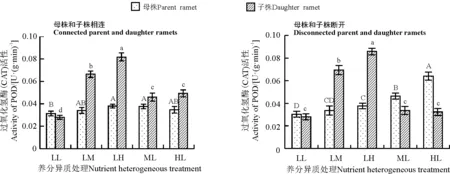
图3 养分异质条件下结缕草克隆分株叶片过氧化氢酶(CAT)活性Fig. 3 CAT activity in clonal ramet leaves of Zoysia japonica under nutrient heterogeneity

图4 养分异质条件下结缕草克隆分株叶片MDA含量Fig. 4 MDA levels in clonal ramet leaves of Zoysia japonica under nutrient heterogeneity
养分异质条件下母株子株断开时,母株MDA含量LL组最高,HL组最低,ML组和HL组与LL组差异显著(P<0.05);子株MDA含量也是LL组最高,LM组和LH组均与LL组差异显著(P<0.05)(图4)。
3 讨论与结论
SOD、POD、CAT是植物体内最主要的3种清除活性氧的膜保护酶,MDA是植物细胞膜脂过氧化的最终产物。在正常情况下植物体内活性氧的产生与防御系统的清除能够维持动态平衡,其酶活性及MDA含量应保持稳定状态。但是在植物生长环境发生变化时,如不同程度干旱、水淹,不同浓度重金属,不同剂量UV-B辐射,不同温度等条件,植物体内这种平衡将被打破,如果长时间处在不利条件下,最终将造成活性氧代谢失调,抗氧化酶活性降低,丙二醛含量升高,细胞膜结构遭到破坏[16-21]。在本研究中,养分条件发生变化时,结缕草分株抗氧化酶活性及丙二醛含量则不同,当母株与子株断开时,可以明显看出低营养水平下母株和子株的SOD、POD、CAT这3种酶活性始终低于中高营养水平,而MDA的含量始终高于中高营养水平,这表明,结缕草分株长时间处于相对低养分条件下时,将造成其酶活性受到一定抑制,细胞内积累氧自由基,细胞膜遭受破坏,这与上述研究结果类似。
本研究结果表明,在养分异质条件下母株与子株相连时,处于中高低养分条件的母株可以显著提高与其相连的子株的SOD、CAT酶活性,降低其MDA含量。说明在养分异质条件下,结缕草母株与与其相连的子株之间发生了生理整合,通过生理整合提高了处于低营养水平的子株的抗氧化酶活性,降低了细胞间活性氧的含量,保护了细胞结构。整合方式可能有二:一是处于中高养分条件的母株通过向处于低养分的子株输出资源,从而提高了子株养分水平,这种资源传输型整合方式在对过路黄(Lysimachiachristinae)的研究中也被发现[22];二是植株通过内源信号主动调节生理代谢激发处于相对不利条件下子株的保护酶活性,而降低了处于相对适宜条件下母株的酶活性。但是,在本研究中当子株处于中高养分条件下,与其相连的处于低养分条件的母株SOD、POD、CAT这3种酶活性没有显著提高,MDA含量降低也不明显。说明此种条件下子株与母株之间并没有出现生理整合,或整合程度非常微弱。克隆植物分株间生理整合并不一定是双向的,其相互之间存在源—汇关系,整合方向是从“源”到“汇”,牛建忠等[23]对野牛草(Buchloedactyloides)的研究表明,在养分异质条件下,姊株能显著地从妹株分株生物量获益,但妹株并不能从姊株获益,属于单向获益;盛丽娟等[24]和冀琳珂等[25]的研究表明,分株的年龄是影响源-汇关系的重要因素之一。本研究中生长年限较长的母株充当了生理整合的源,而生长年限较短的子株则是生理整合的汇,资源及内源信号单向地从母株传向子株,而不能从子株传向母株。
研究表明,无芒雀麦(Bromusinermis)分株在在异质盐分环境下[26],积雪草(Centellaasiatica)分株在异质性重金属镉胁迫下[10],野牛草分株在异质养分条件下[27],都存在生理整合。生理整合可以提高UV-B辐射下活血丹(Glechomalongituba)的抗氧化酶活力[20],提高了淹水胁迫下狗牙根(Cynodondactylon)根部活性氧清除能力[17]。这些结果也支持了养分异质条件下结缕草分株之间存在生理整合,且生理整合可能有助于提高处于相对低养分条件下结缕草分株的抗氧化酶活性,降低其丙二醛含量。
综上所述,养分异质条件下处于中高养分水平的结缕草母株可以显著提高与其相连的处于低养分水平子株的SOD、CAT活性,降低其MDA含量,但POD活性无显著变化;处于中高养分水平的子株对与其相连的处于低养分水平母株的SOD、POD、CAT酶活性的提高及MDA含量的降低都不显著。养分异质条件下结缕草母株对子株SOD、CAT酶活性和MDA含量存在明显生理整合,子株对母株生理整合不明显,属于单向整合;POD活性在母株子株相互之间生理整合不明显。
References:
[1] Hutchings M J,de Kroon H.Foraging in plants:The role of morphological plasticity in resource acquisition.Advances in Ecological Research,1994.25:159-238.
[2] 施建敏,叶学华,陈伏生,杨清培,黎祖尧,方楷,杨光耀.竹类植物对异质生境的适应:表型可塑性.生态学报,2014,34(20):5687-5695.
Shi J M,Ye X H,Chen F S,Yang Q P,Li Z Y,Fang K,Yang G Y.Adaptation of bamboo to heterogeneous habitat:Phenotypic plasticity.Acta Ecologica Sinica,2014,34(20):5687-5695.(in Chinese)
[3] Marshall C.Source-sinkrelation of interconnected ramets.//Clonal Growth in Plants:Regulation and Function.the Hague:SPB Academic Publishing,1990:23-42.
[4] 胡俊靖,陈卫军,郭子武,陈双林,杨清平,李迎春.基于抗氧化系统的美丽箬竹水分生理整合作用分析.生态学杂志,2015,34(4):962-966.
Hu J J,Chen W J,Guo Z W,Chen S L,Yang Q P,Li Y C.Effect analysis of water physiological integration ofIndocalamusdecorusbased on antioxidant system.Chinese Journal of Ecology,2015,34(4):962-966.(in Chinese)
[5] Liu F H,Liu J,Yu F H,Dong M.Water integration patterns in two rhizomatous dune perennials of different clonal fragment size.Flora,2007,202:106-110.
[6] 王沫竹,董必成,李红丽,于飞海.根茎型植物扁秆荆三棱对光照强度和养分水平的生长响应及资源分配策略.生态学报,2016,36(24):8091-8101.
Wang M Z,Dong B C,Li H L,Yu F H.Growth and biomass allocation responses to light intensity and nutrient availability in the rhizomatous herbBolboschoenusplaniculmis.Acta Ecologica Sinica,2016,36(24):8091-8101.(in Chinese)
[7] 赖苏雯,李德志,陈惠娟,耿松,陈青青,贠小涛,樊志华,欧阳义,黄瑶瑶,王明祖.结缕草克隆生长对生境资源梯度水平和次序的响应.草业科学,2015,32(11):1789-1800.
Lai S W,Li D Z,Chen H J,Geng S,Chen Q Q,Yun X T,Fan Z H,Ouyang Y,Huang Y Y,Wang M Z.The clonal growth ofZoysiajaponicain response to the levels and orders of habitat resource gradient.Pratacultural Science,2015,32(11):1789-1800.(in Chinese)
[8] 钱永强,孙振元,韩蕾,巨关升.异质水分环境下野牛草相连分株间光合同化物的生理整合及其调控.生态学报,2010,30(15):3966-3973.
Qian Y Q,Sun Z Y,Han L,Ju G S.Photosynthate integration and regulation within clones of buffalograss under heterogeneous water supply.Acta Ecologica Sinica,2010,30(15):3966-3973.(in Chinese)
[9] 毛舒燕,刘东焕,姜闯道,石雷,张金政,邢全,刘立安.水分胁迫条件下草莓克隆分株间水分调控及其对光合功能的影响.生态学报,2009,29(12):6446-6457.
Mao S Y,Liu D H,Jiang C D,Shi L,Zhang J Z,Xing Q,Liu L A.The effects of water stress on water translocation and photosynthetic characteristics between clonal ramets in strawberry.Acta Ecologica Sinica,2009,29(12):6446-6457.(in Chinese)
[10] 刘富俊,黎云祥,廖咏梅,陈劲松,权秋梅,龚新越.异质性重金属镉胁迫下克隆整合对匍匐茎草本植物积雪草生长的影响.植物生态学报,2011,35(8):864-871.
Liu F J,Li Y X,Liao Y M,Chen J S,Quan Q M,Gong X Y.Effects of clonal integration on growth of stoloniferous herbCentellaasiaticasuffering from heterogeneous heavy metal Cd2+stress.Acta Phytoecologica Sinica,2011,35(8):864-871.(in Chinese)
[11] 李立科,李德志,赵鲁青,潘宇,吕媛媛,赵美霞,高锦瑾,王浩,程立丽,何莹.克隆植物结缕草主匍匐茎中氮素的传输特征.东北林业大学学报,2011,39(11):40-45.
Li L K,Li D Z,Zhao L Q,Pan Y,Lyu Y Y,Zhao M X,Gao J J,Wang H,Cheng L L,He Y.Nitrogen transmission in primary stolons of clonal plantZoysiajaponica.Journal of Northeast Foretry University,2011,39(11):40-45.(in Chinese)
[12] Chapman D F,Hay M J M.Translocation of phosphorus from nodal roots in two contrasting genotypes of white clover (TrifoliumrepensL.).Physiologia Plantarum,1993,89:323-330.
[13] 陈建勋,王晓峰.植物生理学实验指导.广州:华南理工大学出版社,2002:155-157.
Chen J X,Wang X F.Plant Physiology Experiments.Guangzhou:South China University of Technology Press,2002:155-157.(in Chinese)
[14] 高俊凤.植物生理学实验指导.北京:高等教育出版社,2006:210-218.
Gao J F.Plant Physiology Experiments.Beijing:Higher Education Press,2006:210-218.(in Chinese)
[15] 张志良,瞿伟菁.植物生理学实验指导.2.北京:高等教育出版社,1998:258-260.
Zhang Z L,Qu W J.Plant Physiology Experiments.2.Beijing:Higher Education Press,1998:258-260.(in Chinese)
[16] 张彦妮,雷蕾,夏斌.干旱胁迫及复水对黄连花幼苗生长和生理特性的影响.草业科学,2016,33(9):1681-1689.
Zhang Y N,Lei L,Xia B.Effects of drought stress and rewatering on seedling growth and physiological characteristics ofLysimachiadavurica.Pratacultural Science,2016,33(9):1681-1689.(in Chinese)
[17] 李兆佳,喻杰,樊大勇,谢宗强,熊高明,张想英.克隆整合提高淹水胁迫下狗牙根根部的活性氧清除能力.生态学报,2011,31(17):4992-4999.
Li Z J,Yu J,Fan D Y,Xie Z Q,Xiong G M,Zhang X Y.Clonal integration enhances the ability to scavenge reactive oxygen species in root ofCynodondactylonsubjected to submergence.Acta Phytoecologica Sinica,2011,31(17):4992-4999.(in Chinese)
[18] 何洁,高钰婷,贺鑫,刘长发,周一兵.重金属Zn和Cd对翅碱蓬生长及抗氧化酶系统的影响.环境科学学报,2013,33(1):312-320.
He J,Gao Y T,He X,Liu C F,Zhou Y B.The effect of Zn and Cd on growth and antioxidant enzymes activity ofSuaedaheteropteraKitagawa.Acta Scientiae Circumstantiae,2013,33(1):312-320.(in Chinese)
[19] 孙园园,关萍,何杉,石建明.镉胁迫对多花黑麦草镉积累特征、生理抗性及超微结构的影响.草业科学,2016,33(8):1589-1597.
Sun Y Y,Guan P,He S,Shi J M.Effects of Cd stress on Cd accumulation,physiological response and ultrastructure ofLoliummultiflorum.Pratacultural Science,2016,33(8):1589-1597.(in Chinese)
[20] 李倩,毛少利,李为民,李阳.克隆整合提高异质性 UV -B辐射下活血丹抗氧化酶活力.基因组学与应用生物学,2016,35(1):213-217.
Li Q,Mao S L,Li W M,Li Y.Clonal integration enhanced the activity of antioxidant enzymes inGlechomalongitubaunder heterogeneous UV-B radiation.Genomics and Applied Biology,2016,35(1):213-217.(in Chinese)
[21] 刘俊祥,许新桥,钱永强,巨关升,韩蕾,孙振元.多年生黑麦草抗氧化酶和植物络合素对Cd2+胁迫的应答.生态学杂志,2013,32(7):1787-1793.
Liu J X,Xu X Q,Qian Y Q,Ju G S,Han L,Sun Z Y.Responses of antioxidative enzymes and phytochelatins inLoliumperenneto Cd2+stress.Chinese Journal of Ecology,2013,32(7):1787-1793.(in Chinese)
[22] 陈劲松,刘鹏,刘庆.青藏高原东缘过路黄在资源交互斑块性生境中的克隆内资源共享.生态学报,2009,29(7):3532-3539.
Chen J S,Liu P,Liu Q.Intraclonal resource sharing of stoloniferous herbLysimachiachristinain response to reciprocal patchiness of light and nutrients in the eastern edge of the Qing-Zang Plateau in China.Acta Ecologica Sinica,2009,29(7):3532-3539.(in Chinese)
[23] 牛建忠,周禾,张称意,毛培胜,王显国,张佳华.野牛草分株对养分异质性供应的反应.草地学报,2006,14(4):306-309,314.
Niu J Z,Zhou H,Zhang C Y,Mao P S,Wang X G,Zhang J H.Effects of nutrients availability on linked two-ramets of a stoloniferous herb,the buffalograss.Acta Agrestia Sinica,2006,14(4):306-309,314.(in Chinese)
[24] 盛丽娟,李德志,朱志玲,王绪平,石强,王超华,范旭丽,柯世联.克隆植物的碳素生理整合及其生态学效应.应用与环境生物学报,2007,13(6):888-894.
Sheng L J,Li D Z,Zhu Z L,Wang X P,Shi Q,Wang C H,Fan X L,Ke S L.Carbon physiological integration in clonal plants and its ecological effects.Chinese Journal of Applied & Envionmental Biology,2007,13(6):888-894.(in Chinese)
[25] 冀琳珂,谢锦忠,张玮,卢鹏,张磊.雷竹克隆系统出笋期有机碳分布变化规律.生态学报,2016,36(23):7624-7634.
Ji L K,Xie J Z,Zhang W,Lu P,Zhang L.Organic carbon allocation pattern and changes regulation in various organs ofPhyllostachysviolascensclone system in shooting period.Acta Ecologica Sinica,2016,36(23):7624-7634.(in Chinese)
[26] 杨慧玲,薛瑞丽,叶永忠,王会勤,董鸣,黄振英.克隆整合对无芒雀麦在异质性盐分环境中存活和生长的影响.生态学报,2009,29(6):2827-2834.
Yang H L,Xue R L,Ye Y Z,Wang H Q,Dong M,Huang Z Y.Effects of clonal integration on growth and survival ofBromusinermis(Poaceae) in heterogeneous saline environments of the Otindag Sandland.Acta Ecologica Sinica,2009,29(6):2827-2834.(in Chinese)
[27] 罗栋,钱永强,刘俊祥,韩蕾,李伟,孙振元.克隆植物野牛草对异质营养的表型可塑性响应.草业学报,2014,23(3):104-109.
Luo D,Qian Y Q,Liu J X,Han L,Li W,Sun Z Y.Phenotypic responses of a clonal plant (Buchloedactyloides) to nutrient heterogeneity.Acta Prataculturae Sinica,2014,23(3):104-109.(in Chinese)
猜你喜欢
世界科学(2020年1期)2020-02-11
中国生物医学工程学报(2019年5期)2019-07-16
西藏农业科技(2019年2期)2019-07-10
Coco薇(2017年5期)2017-06-05
果树实用技术与信息(2016年8期)2016-09-21
云南师范大学学报(自然科学版)(2015年5期)2015-12-26
现代电生理学杂志(2015年4期)2015-07-18
中央民族大学学报(自然科学版)(2015年2期)2015-06-09
物理实验(2015年10期)2015-02-28
公务员文萃(2013年5期)2013-03-11
