
钙对马尾松针叶生理生化特性及细胞超微结构的影响
2018-04-23 12:52李德燕周运超
西北林学院学报 2018年2期
李德燕,周运超
(1.贵州大学 贵州省森林资源与环境研究中心,贵州 贵阳 550025;2.安顺学院,贵州 安顺 561000;3.贵州大学 林学院,贵州 贵阳 550025)
钙是植物生长发育的必需营养元素之一,在维持植物正常生长、稳定细胞壁、维持细胞膜通透性及膜蛋白的稳定性等方面发挥着重要作用,还作为细胞内第二信使参与植物的生长发育、光合作用电子传递与光合磷酸化、渗透调节作用及激素调控等生理生化过程[1-3]。马尾松(Pinusmassoniana)是我国特有的工业用材树种之一,分布于我国南方17个省(自治区、直辖市),是针叶树种中分布最广、综合利用程度最高的乡土树种。目前,有关钙对马尾松影响相关的报道,主要集中在施肥[4-6]、不同岩性土壤[7]、生物循环[8-10]、种子发芽[11-12]、幼苗生理特征[13]及蛋白质组学[14-15]等方面的研究,并取得了一系列丰硕的成果,表明钙对马尾松抗逆性、生长发育、产量等方面均具有重要影响。
我国酸雨区主要分布在长江以南[16],马尾松林地土壤中的盐基离子在酸雨的作用下发生淋失作用,特别是Ca2+、Mg2+、K+和Na+流失严重[17-18]。此外,长期种植马尾松引起林地水土流失加重,同时周期性采伐也使得林地大量养分消耗,使钙在生态系统中的循环受到影响[19-21],导致马尾松生长受抑,生产力降低。研究表明,钙浓度与马尾松幼苗的株高和地径生长情况密切相关[13],因而马尾松林地土壤中有效性钙含量多少与其林分效益紧密相关。然而,目前有关钙对马尾松生理生化影响的报道较少,而有关钙对马尾松针叶的渗透调节物质和内源激素含量以及细胞超微结构的影响特征尚不清楚。为此,本文以马尾松1年生苗为研究对象,通过砂培模拟试验探究钙对马尾松苗生理生化特征及细胞超微结构的影响,阐明马尾松对钙的适应性机理,为马尾松人工林种植及管理提供参考。
1 材料与方法
1.1 材料培养与处理
试验于2015年3月至8月在贵州大学林学院苗圃温室内进行。试验材料为生长健壮且粗度和高度一致的1年生马尾松苗,栽培基质为3%稀盐酸浸泡后蒸馏水充分清洗后的石英砂,每盆10 kg。试验共设6个处理,每处理15盆,每盆种植3株。营养液为Hoagland营养液配加Amon微量元素,6个供Ca2+水平分别为0、0.4、1、2、3 mmol·L-1和4 mmol·L-1,其中4 mmol·L-1模拟酸性土壤(pH 5.8左右)溶液中平均Ca2+浓度[22-24],Ca2+以Ca(NO3)2形式添加。马尾松生长期间,每3 d浇1次(250 mL/盆),并在第3次浇营养液前一天用大量蒸馏水洗盐1次,之后按此法浇营养液直到试验结束。于苗木移植后第80天,每个处理随机选取10株马尾松苗不同方向的健康针叶(功能叶)测定相对电导率、叶绿素、可溶性糖、淀粉和可溶性蛋白和内源激素含量等指标,以及针叶叶肉细胞观察;同时随机选取15株马尾松苗用于生物量测定。
1.2 测定指标及方法
将取出的马尾松植株洗净后,放入干净搪瓷盘中置于恒温箱内105℃杀青30 min,70℃烘至恒重后测其生物量。参照叶尚红[25]的方法测定马尾松针叶相对电导率,采用95%乙醇提取后测定叶绿体含量,采用蒽酮法测定可溶性糖与淀粉含量,采用考马斯亮蓝G-250法测定可溶性蛋白含量;参照杨卫兵等[26]的方法采用高效液相色谱测定针叶中激素含量(吲哚乙酸,IAA;脱落酸,ABA;玉米素核苷,ZR;赤霉素,GA);选择春梢中段南向健康针叶,取针叶中部0.5 cm为材料,参照王钧等[27]的方法进行处理,用LIMB V型超薄切片机将包埋的样品横切成80 nm的薄片后,用醋酸铀-柠檬酸铅双层染色后在H-7650型透射电镜上观测及拍照。
1.3 数据处理
采用SPSS(20.0)、Excel(2013)分析软件进行数据统计和差异显著性(P<0.05)分析,多重比较采用新复极差法进行检验。
2 结果与分析
2.1 钙对马尾松生长的影响
不同钙浓度下,马尾松苗生长情况存在一定差异(图1和图2)。2 mmol·L-1Ca2+处理下的马尾松生长较旺盛,株高生长量最大,针叶较宽且粗细均匀,主根较长,根尖个数最多,生物量最大;1 mmol·L-1和3 mmol·L-1Ca2+处理的植株生长势次之,与2 mmol·L-1Ca2+处理相比,针叶宽度和厚度降低,主根长、根体积、根尖数和生物量也有所降低;而0 mmol·L-1和4 mmol·L-1Ca2+处理的生长势相对较差,其中0 mmol·L-1Ca2+的植株生物量相对较低。这说明马尾松幼苗在土壤溶液有效钙含量为2 mmol·L-1左右时生长较适宜。
2.2 钙对叶绿体含量的影响
叶绿素含量作为植物抗逆性的重要指标之一,是植物养分状况、光合作用能力和生长态势的重要指示因子。从表1可知,随着Ca2+增加,马尾松苗针叶内叶绿素a、叶绿素b、叶绿素a+b和叶绿素a/b含量均呈先升高后降低的趋势,最高值均出现在2 mmol·L-1Ca2+处理中,而最低值依次出现在4 mmol·L-1、4 mmol·L-1、4 mmol·L-1和0 mmol·L-1Ca2+处理中,依次低于相应最高值18.65%、17.66%、18.39%和10.21%。可见,2 mmol·L-1Ca2+处理下马尾松光合作用能力较强,利于马尾松生长和发育。
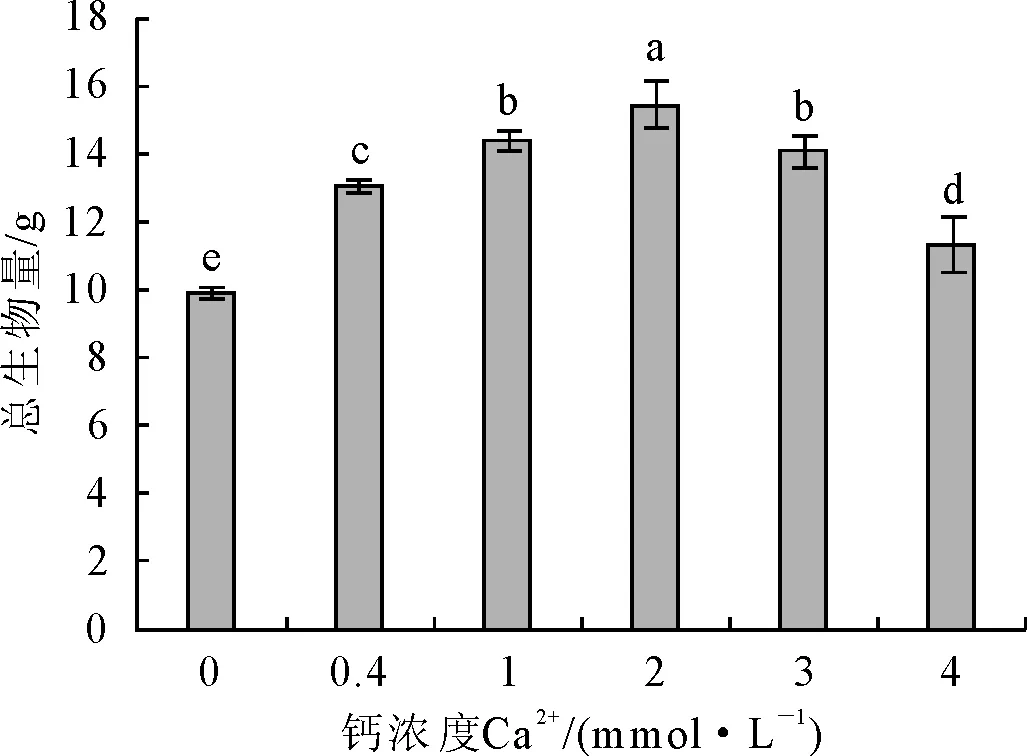
注:不同小写字母表示处理间差异显著(P<0.05)。下同。
图1不同钙浓度对马尾松生物量的影响
Fig.1 Effect of different Ca2+concentrationson total biomass ofP.massoniana

图2 不同钙浓度对马尾松生长状况的影响
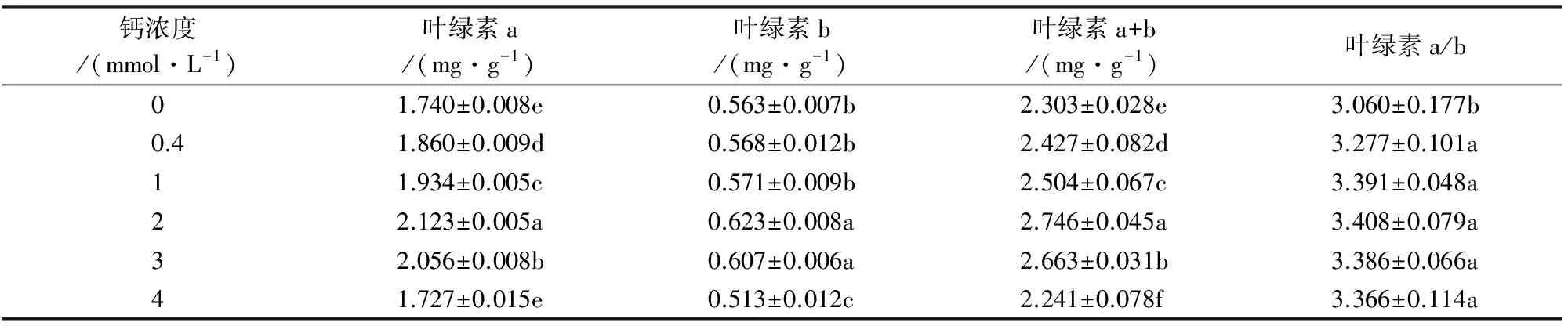
钙浓度/(mmol·L-1)叶绿素a/(mg·g-1)叶绿素b/(mg·g-1)叶绿素a+b/(mg·g-1)叶绿素a/b01.740±0.008e0.563±0.007b2.303±0.028e3.060±0.177b0.41.860±0.009d0.568±0.012b2.427±0.082d3.277±0.101a11.934±0.005c0.571±0.009b2.504±0.067c3.391±0.048a22.123±0.005a0.623±0.008a2.746±0.045a3.408±0.079a32.056±0.008b0.607±0.006a2.663±0.031b3.386±0.066a41.727±0.015e0.513±0.012c2.241±0.078f3.366±0.114a
注:不同小写字母表示处理间差异显著(P<0.05)。
2.3 钙对相对电导率、可溶性糖、蛋白质和淀粉含量的影响
叶片相对电导率是植物受害的敏感指标之一,与细胞膜透性正相关。可溶性糖和可溶性蛋白质是植物体内重要的渗透调节物质,植物通过大量积累渗透调节物质以保持体内膨压,从而维持植物正常的生理过程。淀粉作为植物主要的长期储存物质之一,并在一定条件下可与可溶性糖相互转化。马尾松针叶相对电导率、可溶性糖和蛋白质含量随供Ca2+浓度增大先降低后升高,最低值都在2 mmol·L-1Ca2+处理中出现,而最高值依次出现在4 mmol·L-1、4 mmol·L-1和0 mmol·L-1Ca2+处理中,比2 mmol·L-1Ca2+处理依高出了12.41%、13.80%和17.15%;淀粉含量随着钙浓度增加先升高后降低,2 mmol·L-1Ca2+处理(最大值)的淀粉含量比0 mmol·L-1Ca2+处理(最小值)高出16.28%(表2)。由此可知,与2 mmol·L-1Ca2+处理相比, 0 mmol·L-1和4 mmol·L-1Ca2+处理下的针叶细胞膜受损严重,马尾松通过积累大量的渗透调节物质以保持体内的膨压,从而维持其细胞内生理代谢活动的稳定。
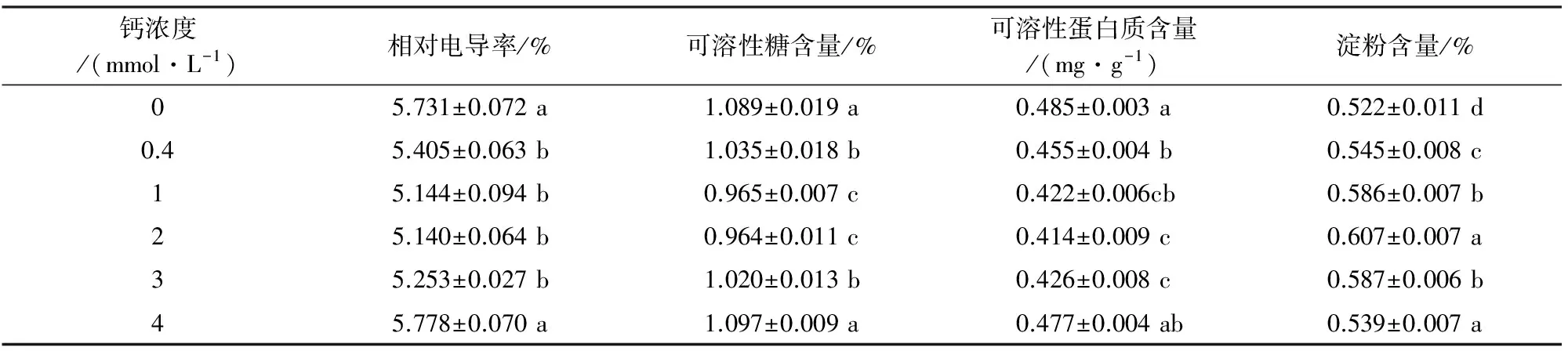
表2 不同钙浓度对马尾松针叶相对电导率、可溶性糖、蛋白质和淀粉含量的影响
2.4 钙对内源激素的影响
2.4.1 内源激素含量 一般认为,植物内源激素IAA、GA和ZR是生长促进型激素,而ABA是生长抑制型激素。从图3可知,马尾松针叶内GA、ZR和IAA含量随供Ca2+浓度增加呈先增加后降低的趋势,最高值分别出现在2 mmol·L-1、1 mmol·L-1和2 mmol·L-1Ca2+处理中,而最低值依次出现在4 mmol·L-1、0 mmol·L-1和4 mmol·L-1Ca2+处理中,分别比最高值降低了30.04%、35.85%和37.28%(图3A~图3C)。针叶内ABA含量随供Ca2+浓度增加先降低后增加,其最高值和最低值分别出现在4 mmol·L-1和2 mmol·L-1Ca2+处理中,而最高值是最低值的1.43倍。由此说明,不同供Ga2+水平下的马尾松,通过改变内源激素的含量水平,调节其体内生理代谢的各个环节,以利于自身生长和发育。
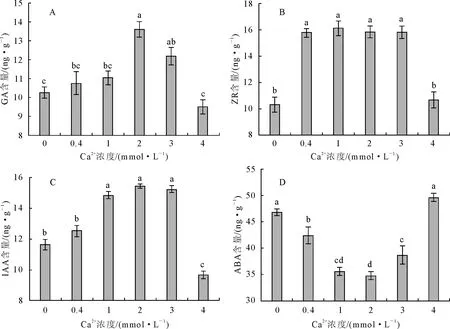
图3 不同钙浓度对马尾针叶内源激素含量的影响
2.4.2 内源激素间比例 由图4可知,马尾松针叶中ABA/GA、ABA/ZR、ABA/IAA和ABA/(GA+ZR+IAA)值随着钙浓度增加均呈现先降低后增加的趋势,但比值的大小存在差异。其中,最低值均出现在2 mmol·L-1Ca2+的处理中,最高值均出现在4 mmol·L-1Ca2+的处理中,1~3 mmol·L-1Ca2+处理下的马尾松针叶中ABA/GA、ABA/ZR、ABA/IAA和ABA/(GA+ZR+IAA)比值无明显变化。说明不同供Ca2+水平下的马尾松,其生理过程受体内多种激素的协同作用,通过调节激素含量间比值,共同决定其生长发育情况。
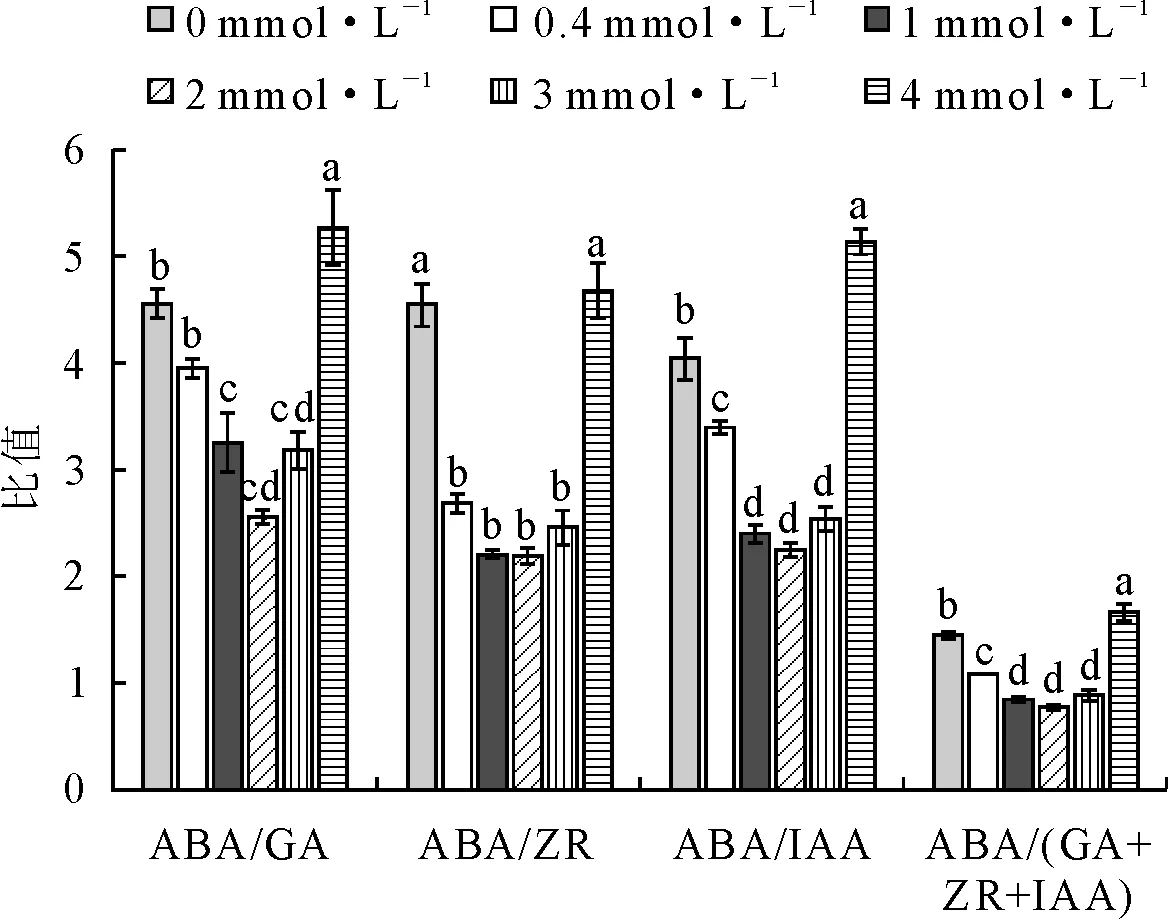
图4 钙浓度对马尾针叶内源激素比例的影响
2.5 钙对针叶细胞超微结构的影响
植物的形态结构与自然环境密切相关,其结构的变化与生理机能的统一是植物适应环境的基础。不同钙浓度下马尾松苗的叶绿体内均有不同程度的淀粉粒出现,同时细胞中的原生质层与细胞壁的关系也存在一定差异(图5)。供Ca2+浓度为2 mmol·L-1的马尾松针叶细胞壁内外壁平滑、完整,细胞核、叶绿体与线粒体形态较完整,未出现质壁分离现象;叶绿体中淀粉粒含量较多,间质浓密,嗜锇颗粒含量较少;线粒体数量不多,嵴丰富、清晰,内含物较多(5D1-3、5E1-3)。在0~1 mmol·L-1Ca2+的处理中,随着钙浓度增加,马尾松针叶叶肉细胞中质壁分离程度逐渐减弱,叶绿体和线粒体形态逐渐变得完整、规则,淀粉粒含量逐渐增多,嗜锇颗粒逐渐减少;其中,0 mmol·L-1Ca2+处理中部分叶绿体空泡化,并有部分线粒体被膜出现破裂,嵴结构变得模糊(5A1-3,5B1-3,5C1-3)。供Ca2+浓度为3~4 mmol·L-1的处理中,随Ca2+浓度增加,叶肉细胞中产生质壁分离现象加重,淀粉粒逐渐减少,嗜锇颗粒含量增多,线粒体数目增多,部分线粒体被膜出现破裂现象(5E1-3)。除2 mmol·L-1Ca2+处理的马尾松针叶细胞未出现质壁分离外,其余处理中细胞质壁分离的程度依次为3<1<0.4<4<0 mmol·L-1Ca2+,与细胞中的嗜锇颗粒含量和线粒体数的变化趋势相反,表明0 mmol·L-1Ca2+处理的马尾松针叶细胞结构受损程度相对较重,其次为4 mmol·L-1Ca2+处理。
3 结论与讨论
叶绿素含量是反映光合器官生理状况的重要值指标,其含量在一定程度反映了植物生长状况和光合作用能力,也可表征逆境中植物组织、器官的衰老状况[28-30]。植物细胞膜对维持细胞的微环境和正常生理代谢具有重要作用,而相对电导率是衡量细胞膜透性的重要指标,其大小反应了植物细胞受损的程度。植物可通过调节体内的渗透调节物质含量以避免或减少逆境带来的伤害。本试验中,适宜的钙(2 mmol·L-1Ca2+)能显著提高马尾松的光合色素(叶绿素a、叶绿素b和叶绿素a+b)含量、淀粉含量及叶绿素a/b值,降低针叶细胞的电导率、可溶性糖和蛋白质含量,有利于增强马尾松的净光合速率和有机物的积累,从而促进植株生长。0~1 mmol·L-1和 3~4 mmol·L-1Ca2+处理的马尾松针叶中光合色素降低,造成针叶捕光能力和光合活性降低,光合有机物合成能力下降,同时细胞膜透性增加,细胞内自由基的产生和清除平衡遭到破坏,引起膜脂过氧化反应,导致细胞膜受损、膜透性增加和细胞内电解质外渗,生理生化代谢遭到破坏[13]。然而,钙胁迫条件下马尾松针叶中部分淀粉水解,增加了可溶性糖含量,为生长提供所需的物质和能量,有利于植株维持一定渗透势,可有效缓解钙胁迫对马尾松造成的不利影响。但是,针叶细胞中蛋白质的合成和降解平衡被打破,引起蛋白质含量降低,导致呼吸代谢增强[31],使植株体内养分耗损增加,有机物质积累能力下降,最终表现为植株生物量降低。这可能与钙胁迫下马尾松对N的吸收代谢过程有关[32],其原因有待深入研究。
内源激素在植物生长发育过程中起着重要的调控作用,作为信号分子参与了一系列与逆境胁迫相适应的形态和生理生化变化[33]。IAA、GA和ZR为生长促进型激素,具有促进植物苗生长、延缓衰老的功效,而ABA作为生长抑制型激素,在逆境中可诱导相应的基因表达,从而提高植物对逆境胁迫的抗性[34]。本试验结果表明,适宜钙浓度可增加马尾松苗针叶中IAA、ZR和GA的含量,降低ABA含量、ABA/IAA、ABA/GA、ABA/ZR和ABA/(IAA+ZR+GA)的比值,而与高钙或低钙下针叶的内源激素含量及比值变化相反,表明马尾松内激素并不是单一激素起作用,而是共同协调发挥作用。同时,也表明马尾松通过降低生长促进型激素含量,减缓植株的生长速率,以适应钙胁迫环境。马尾松针叶ABA的积累会诱导气孔关闭,可引起针叶的光合速率、气孔导度和蒸腾速率相应降低[35],这是其应对钙胁迫的一种生理反应。ABA作为初始信号与ABA受体结合,并在磷酸酶和激酶的作用下激活细胞内的第二信使并诱导钙胁迫相关基因表达,确保马尾松细胞膜系统完整以维持细胞正常渗透势或膨压[36],从而提高马尾松对钙胁迫的适应性。
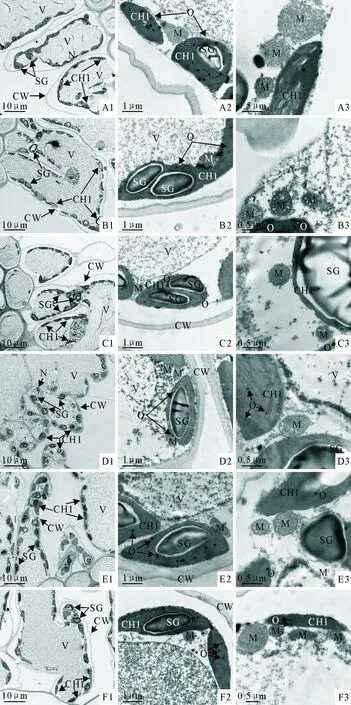
注:A1-A3、B1-B3、C1-C3、D1-D3、E1-E3 和F1-F3依次为0 mmol·L-1、0.4 mmol·L-1、1 mmol·L-1、2 mmol·L-1、3 mmol·L-1和4 mmol·L-1Ca2+处理;A1,B1,C1,D1,E1×1 500 s; A2,B2,C2,D2,E2×15 000; A3,B3,C3,D3,E3×30 000; Chl:叶绿体; CW:细胞壁; N:细胞核; O:嗜锇颗粒; SG:淀粉粒; V:vacuole; M:mitochondria。
图5钙对马尾针叶超微结构的影响
Fig.5 Effect of different Ca2+concentrations on the ultra-structure ofP.massoniananeedle tissue
完整的细胞结构是植物体维持正常生理功能的基础。逆境条件下,植物细胞器结构破坏,导致细胞生理功能降低甚至丧失,是植物遭受逆境伤害的重要原因之一。逆境中的植物因细胞内水分外渗,原生质随着液泡一起收缩而发生质壁分离。本试验中,除2 mmol·L-1Ca2+处理的马尾松叶肉细胞未观察到质壁分离现象外,其余处理均发生不同程度质壁分离现象,而细胞质壁分离程度与马尾松幼苗植株生物量变化趋势相反,表明钙胁迫下马尾松的细胞功能受损,正常生理活动受到一定影响,导致其生物量降低。叶绿素a/b值反映了叶绿体中内囊体的垛叠程度,而内囊体作为光能吸收、传递和转换的场所,低钙或高钙条件下马尾松针中叶绿素a/b值降低,说明了光能转化为生物化学能的能力降低[37]。嗜锇颗粒由叶绿体或类囊体的膜脂被ROS氧化而形成[38],故马尾松针叶细胞中嗜锇颗粒增加表明叶绿体内部膜脂过氧化程度加剧,从而对马尾松的生长产生抑制作用。淀粉作为光合作用的产物,淀粉粒数量减少说明马尾松对钙胁迫采取了积极的防御措施,通过淀粉粒降解而合成更多有机溶质,从而调节渗透压[39]。线粒体是氧化磷酸化、释放能量的场所,其数目与细胞的生理功能及需求密切相关,马尾松针叶细胞中叶绿体周围的线粒体数量明显增多,说明线粒体数量增多可能是对钙胁迫下单位线粒体活性降低的一种补偿方式[40-41],一定程度上缓减了钙对马尾松的胁迫作用。因此,低钙或高钙引起马尾松针叶叶绿体和类囊体发育异常,使得叶绿体膜结构和类囊体基粒片层排列遭到一定破坏,淀粉粒降解,嗜锇颗粒增多,部分线粒体膜破损且内嵴模糊,质壁分离现象发生,导致马尾松针叶细胞的生理生化代谢发生紊乱。由此可见,细胞形态结构的改变是钙胁迫下马尾松生物量降低的重要因素之一。
综上所述,不同供钙水平下,马尾松幼苗可通过调节体内的激素含量及比值,稳定细胞内正常生理生化代谢进程,从而保护细胞膜、细胞器的结构和功能完整,改善针叶的光合状况,以利于其生长发育,其中2 mmol·L-1Ca2+对马尾松苗生长较有利。因此,在马尾松人工林培育过程中,根据林地土壤中钙含量及其生长状况进行科学合理补充钙肥,对实现马尾松人工林可持续发展具有重要意义。
参考文献:
[1] FERGUSON I B,DROBAK B K.Calcium and the regulation of plant growth and senescence[J].Journal of Materials Research,1988,28(17):2339-2350.
[2] WARD M R,ASLAM M,HUFFAKER R C.Enhancement of nitrate uptake and growth of barley seedlings by calcium under saline conditions[J].Plant Physiology,1986,80(1):520-524.
[3] BUSH D S.Calcium regulation in plant cells and its role in signaling[J].Annual Review of Plant Physiology & Plant Molecular Biology,1995,46(1):95-122.
[4] 周运超,谌红辉.施肥对马尾松针叶营养成分的影响[J].山地农业生物学报,2000,19(2):99-103.
ZHOU Y C,CHEN H H.Effect of fertilization on foliar nutrient content of Masson Pine[J].Journal of Mountain Agriculture and Biology,2000,19(2):99-103.(in Chinese)
[5] 王金凤.酸雨对马尾松针叶结构、生理和器官元素含量的影响[D].南宁:广西大学,2004.
[6] 黄永梅,段雷,靳腾,等.投加石灰石和菱镁矿对酸化土壤上马尾松(Pinusmassoniana)林的影响[J].生态学报,2006,26(3):786-792.
HUANG Y M,DUAN L,JIN T,etal.Effect of limestone and magnesite applications on masson pine (Pinusmassoniana) forest growing on acidified soil[J].Acta Ecologica Sinica,2006,26(3):786-792.(in Chinese)
[7] 周政贤.中国马尾松[M].北京:中国林业出版社,2001.
ZHOU Z X.Masson Pine in China[M].Beijing:China Forestry Publishing House,2001.(in Chinese)
[8] 刘文飞,樊后保,谢友森,等.闽西北马尾松人工林营养元素的积累与分配格局[J].生态环境学报,2008,17(2):708-712.
LIU W F,FAN H B,XIE Y S,etal.Nutrient accumulation and distribution in a Masson pine stand in Northwestern Fujian[J].Ecology and Environmental,2008,17(2):708-712.(in Chinese)
[9] 吴飞华,刘廷武,裴真明,等.酸雨引起森林生态系统钙流失研究进展[J].生态学报,2010,30(4):1081-1088.
WU F H,LIU T W ,PEI Z M ,etal.Calcuim depletion in forest ecosystem induced by acid rain areview[J].Acta Ecologica Sinica,2010,30(4):1081-1088.(in Chinese)
[10] 杨会侠,汪思龙,范冰,等.马尾松人工林发育过程中的养分动态[J].应用生态学报,2010,21(8):1907-1914.
YANG H X,WANG S L,FAN B,etal.Dynamics of nutrients in an age sequence ofPinusmassonianaplantation.[J].Chinese Journal of Applied Ecology,2010,21(8):1907-1914.(in Chinese)
[11] 韦小丽,廖明.钙浸种对马尾松种子发芽的影响[J].种子,2005,24(4):34-36.
WEI X L,LIAO M.Effects of Calcium-soaking on Germination ofPinusmassonianaSeed[J].Seed,2005,24(4):34-36.(in Chinese)
[12] 简才源,韦小丽,陈艳伟,等.多种引发剂混合处理对马尾松种子发芽的影响[J].种子,2016,35(10):23-27.
JIAN C Y,WEI X L,CHEN Y W,etal.Effects of multiple initiators treatments on the seed germination ofPinusmassoniana[J].Seed,2016,35(10):23-27.(in Chinese)
[13] 李德燕,周运超.钙浓度对马尾松幼苗生长和生理特征的影响[J].林业科学研究,2017,30(1):174-180.
LI D Y,ZHOU Y C.Effects of calcium concentration on growth and physiological characteristics ofPinusmassonianaseedling[J].Forest Research,2017,30(1):174-180.(in Chinese)
[14] 赵振洲.马尾松种子发育过程元素及蛋白质组学特征[D].南京:南京林业大学,2009.
[15] HU W J,CHEN J,LIU T W,etal.Proteome and calcium-related gene expression inPinusmassoniana,needles in response to acid rain under different calcium levels[J].Plant & Soil,2014,380(1-2):285-303.
[16] REN R.Acid rain in china past,present and its future[J].Journal of Beijing Polytechnic University,1997,23(3):129-132.
[17] 刘秀菊,周国逸.土壤累积酸化对鼎湖山马尾松林物质元素迁移规律的影响[J].浙江大学学报:农业与生命科学版,2005,31(4):381-391.
LIU X J,ZHOU G Y.Effects of cumulative acidification of soil on element transfer inPinusmassonianaLamb.forest at Dinghushan[J].Journal of Zhejiang University:Agric.& Life Sci.,2005,31( 4):381-391.(in Chinese)
[18] 杨平平,徐仁扣,黎星辉.淋溶条件下马尾松针对土壤的酸化作用[J].生态环境学报,2012,21(11):1817-1821.
[19] STUPAK I S,NORDFJELL T N,GUNDERSEN P G.Comparing biomass and nutrient removals of stems and fresh and predrie[J].Canadian Journal of Forest Research,2007,38(10):2660-2673.
[20] LATTIMORE B,SMITH C T,TITUS B D,etal.Environmental factors in woodfuel production:Opportunities,risks,and criteria and indicators for sustainable practices[J].Biomass and Bioenergy,2009,33(10):1321-1342.
[21] 葛晓改,周本智,肖文发.马尾松人工林凋落物产量、养分含量及养分归还量特性[J].长江流域资源与环境,2014,23(7):996-1003.
[22] 饶立华.植物矿质营养及其诊断[M].北京:农业出版社,1993.
[23] 罗绪强,王世杰,张桂玲,等.钙离子浓度对两种蕨类植物光合作用的影响[J].生态环境学报,2013(2):258-262.
③全面开展监测评价。建成11个坡地径流场、163个观测小区和14个沟道控制站,基本覆盖全市的水土流失监测网络。依托科技部中德合作项目,引进德国先进设备,构建土壤蒸渗仪监测点→坡面土壤侵蚀和平原区监测点→小流域监测→大流域监测构成的监测网络,对降水、径流、泥沙、土壤水分循环、水质水量、生物多样性、水文形态特征等开展监测,及时评估水土保持效益,为生态清洁小流域建设提供科学依据。
[24] 耿增超,李新平.园林土壤肥料学[M].西安:西安地图出版社,2002.
[25] 叶尚红.植物生理生化实验教程[M].2版.昆明:云南科技出版社,2007.
[26] 杨卫兵,王振林,尹燕枰,等.外源ABA 和GA 对小麦籽粒内源激素含量及其灌浆进程的影响[J].中国农业科学2011,44(13):2673-2682.
[27] 王钧,邬卉,薛生国,等.锰胁迫对杠板归细胞超微结构的影响[J].生态学报,2014,34(4):798-806.
WANG J,WU H,XUE S G,etal.Manganese stress on the ultrastructures of a manganese tolerant plant,PolygonumperfoliatumL.[J].Acta Ecologica Sinica,2014,34(4):798-806.(in Chinese)
[28] NIEVA F J J,CASTILLO J M,LUQUE C J.Ecophysiology of tidal and non-tidal population of the invading cordgrass Spartina densiflora:seasonal and diurnal patterns in a mediterranean climate[J].Estuarine,Coastal and Shelf Science,2005,57:919-928.
[29] 张刚,魏典典,邬佳宝,等.干旱胁迫下不同种源文冠果幼苗的生理反应及其抗旱性分析[J].西北林学院学报,2014,29(1):1-7.
ZHANG G,WEI D D,WU J B,etal.Effects of drought stress on physiological characteristics ofXanthorocerassorbifoliafrom different provenances and analysis on drought-resistance in seedling stage[J].Journal of Northwest Forestry University,2014,29(1):1-7.(in Chinese)
[30] 梁小红,安勐颍,宋峥,等.外源甜菜碱对低温胁迫下结缕草生理特性的影响[J].草业学报,2015,24(9):181-188.
LIANG X H,AN M Y,SONG Z,etal.Effects of exogenous glycine betaine on the physiological characteristics ofZoysiajaponicaunder low-temperature stress[J].Acta Prataculturae Sinica,2015,24(9):181-188.(in Chinese)
[31] 田莉莉,方金豹,王力荣,等.华光油桃解除休眠过程中几项生理指标的变化[J].果树学报,2006,23( 1):121-124.
TIAN L L,FANG J B,WANG L R,etal.Changes of several physiological indexes during dormancy releasing in nectarine cultivar Huaguang[J].Journal of Fruit Science,2006,23(1):121-124.(in Chinese)
[32] 周录英,李向东,王丽丽,等.钙肥不同用量对花生生理特性及产量和品质的影响[J].作物学报,2008,34(5):879-885.
ZHOU L Y,LI X D,WANG L L,etal.Effects of different Ca applications on physiological characteristics,yield and quality in peanut[J].Acta Agronomice Sinica,2008,34(5):879-885.(in Chinese)
[33] RUBIO V,BUSTOS R,IRIGOYEN M L,etal.Plant hormones and nutrient signaling[J].Plant Molecular Biology,2009,69(4):361-373.
[34] MAYABA N,BECKETT R P,CSINTALAN Z,etal.ABA increases the desiccation tolerance of photosynthesis in the afromontane understory mossAtrichumandrogynum[J].Annals of Botany,2001,88:1093-1100.
[35] 张云霞,石勇,王瑞刚,等.初始盐胁迫下ABA与CaM对胡杨叶片气体交换的调控[J].林业科学,2008,44(1):57-64.
ZHANG Y X,SHI Y,WANG R G,etal.Effects of ABA and CaM on leaf gas exchange ofPopuluseuphraticain the process of initial salinity[J].Scientia Silvae Sinicae,2008,44(1):57-64.(in Chinese)
[36] MEHROTRAAR,BHALOTHIA P,BANSAL P,etal.Abscisic acid and abiotic stress tolerance-different tiers of regulation[J].Journal of Plant Physiology,2014,171(7):486-496.
[37] 刘志华,时丽冉,白丽荣,等.盐胁迫对獐毛叶绿素和有机溶质含量的影响[J].植物生理与分子生物学学报,2007,33(2):165-172.
LIU Z H,SHI L R,BAI L R,etal.Effects of salt stress on the contents of chlorophyll and organic solutes inAeluropuslittoralisvar.sinensisDebeaux[J].Journal of Plant Physiology and Molecular Biology,2007,33(2):165-172.(in Chinese)
[38] ZHANG M P,ZHANG C J,YU G H,etal.Changes in chloroplast ultrastructure,fatty acid components of thylakoid membrane and chlorophyll a fluorescence transient in flag leaves of a super-high-yield hybrid rice and its parents during the reproductive stage[J].Journal of Plant Physiology,2010,167:277-285.
[39] 梁文斌,薛生国,沈吉红,等.锰胁迫对垂序商陆叶片形态结构及叶绿体超微结构的影响[J].生态学报,2011,31(13):3677-3683.
LIANG W B,XUE S G,SHEN J H,etal.Manganese stress on morphological structures of leaf and ultrastructures of chloroplast of a manganese hyperaccumulatorPhytolaccaamericana[J].Acta Ecologica Sinica,2011,31(13):3677-3683.(in Chinese)
[40] 黎明,杨芳绒,商卫卫.低温对宜昌润楠生理指标及叶片超微结构的影响[J].西北林学院学报,2010,25(4):19-22.
LI M,YANG F R,SHANG W W.Effects of low temperature on cold resistance and leaf ultramicroscopic structure inMachilusichangensis[J].Journal of Northwest Forestry University,2010,25(4):19-22.(in Chinese)
[41] 濮晓珍,尹春英,周晓波,等.铁皮石斛组培苗移栽驯化过程中叶片光合特性、超微结构及根系活力的变化[J].生态学报,2012 ,32(13):4114-4122.
PU X Z,YIN C Y,ZHOU X B,etal.Changes in photosynthetic properties,ultrastructure and root vigor ofDendrobiumcandidumtissue culture seedlings during transplantation[J].Acta Ecologica Sinica,2012,32(13):4114-4122.(in Chinese)
猜你喜欢
农业灾害研究(2022年6期)2022-12-02
绿色科技(2019年5期)2019-11-29
音乐研究(2019年5期)2019-11-22
湖南饲料(2019年4期)2019-10-17
中国生物医学工程学报(2019年5期)2019-07-16
作文周刊·小学四年级版(2019年8期)2019-04-23
西南林业大学学报(2018年5期)2018-10-23
国际木业(2018年3期)2018-09-10
中国粮油学报(2018年12期)2018-03-19
农业环境科学学报(2017年11期)2017-12-07
