
筇竹叶片解剖结构的研究
2018-04-23 12:53董文渊郑静楠张孟楠
西北林学院学报 2018年2期
刘 培,董文渊,郑静楠,浦 婵,张孟楠
(1.西南地区生物多样性保育国家林业局重点实验室(西南林业大学),云南 昆明 650224; 2.西南林业大学 云南生物多样性研究院,云南 昆明 650224;3.西南林业大学 环境科学与工程学院,云南 昆明 650224)
筇竹(Qiongzhueatumidinoda),属禾本科(Gramineae)筇竹属(Qiongzhuea)植物, 为地下茎复轴混生的小型竹种,被列为国家三级保护的珍稀濒危竹种之一[1],仅分布于中国的西南部分地区,筇竹节间隆起独特,是重要的工艺用竹,筇竹笋味道鲜美,营养丰富,远销海内外,具有重要的生态价值、观赏价值和经济价值。近年来筇竹无性系种群严重退化,同时开花现象较为常见,群数量急剧减少[2-3]。竹类植物有生长快速、分布广泛、使用多样性等特点,是木材不可或缺的替代物[4]。植物叶结构特征与系统分类[5-6]、光合速率[7]和抗逆性[8]等密切相关。解剖形态观察和比较分析作为认识、保护、开发筇竹的有效途径却鲜见报道[9]。筇竹叶解剖学研究作为筇竹研究的基础,是筇竹后续研究不可缺少的一部分,将对筇竹资源的可持续开发利用和保护提供重要的依据。
1 材料与方法
1.1 试验材料与仪器
材料:筇竹叶片2016年2-3月采集于云南省昭通市大关县木杆镇细沙村大罗汉坝。该地区地处云南省东北部,属于中温带气候,温差较小,年平均气温9~12℃,年平均光照966.7 h,年平均降水量991.7 mm。试验地竹林海拔1 550~1 640 m,坡度39°~44°。土壤以黄棕壤为主,成土母岩为花岗岩,土层深厚且湿润,呈酸性反应;林分郁闭度大,林下形成深厚的腐殖层,林下生长有其他植物。
主要仪器:生物组织包埋机、石蜡切片机、烘片机、Nikon E100光学显微-拍摄系统。
1.2 试验方法
分别取自秆龄1~4年生的筇竹,每一年龄各选取3株,每株从中部冠层取不同枝条上长势良好、无病虫害的成熟叶片,每枝至少3片。将取好的叶片立即放入70%的FAA固定液固定,切片前用15%的HF去硅38 h,流水冲洗1 d,常规石蜡制片,切片厚度8~10 μm,番红-固绿对染,Nikon E100光学显微镜下观察拍照,D3K-MS测量细胞长度、厚度,每类细胞至少测量50个,同时测量以下指标:主脉木质部导管数、叶肉细胞层数、梭形细胞长、梭形细胞宽、泡状细胞数。
1.3 指标测定方法
直接观察制片图片,获取主脉木质部导管数、叶肉细胞层数、泡状细胞数(选取3个视野进行测定,取均值) 。
在40倍镜下利用图像分析系统随机测定梭形细胞、表皮细胞、叶肉细胞、薄壁细胞、厚壁细胞、泡状细胞的长度和宽度(梭形细胞至少测3个,其余细胞每类至少测50个,取均值)。测量的数据利用SPSS 17.0软件进行分析。
2 结果与分析
2.1 筇竹叶片解剖结构
筇竹叶片的显微结构与其他竹类植物的叶片在结构上差异不大,主要分为表皮、叶肉、叶脉3部分(图1)。因此,在种的分类上没有多少参考意义,符合早期一些学者的研究结论[10]。
2.1.1 表皮 筇竹叶片表皮分为上表皮和下表皮且各只有1层细胞,长5.22~20.38 μm,宽3.90~14.68 μm。上表皮细胞较大,具有光滑平厚的角质层(图1B-7),并有少量气孔和硅细胞,在2个维管束间,具有大型泡状细胞(图1A-2,图1B-2)。泡状细胞一般为2~5个,长14.98~34.88 μm,宽9.99~32.97 μm,排列为扇形,中间细胞较大,两旁较小,常深入到叶肉1/3以上的部位。但在叶片的边缘无泡状细胞的分布。下表皮细胞形状差异很大,不存在泡状细胞,角质层向外突出成刺状形成大小不等的乳突(图1A-8,图1B-8),并排列着丰富的气孔和硅质细胞(图1B-3)。
2.1.2 叶肉 筇竹叶肉细胞常为3~4层,靠近上表皮的第1层的细胞较大,排列较整齐,为细胞壁自上而下单向指状向内褶叠的臂细胞(图1B-4),长7.67~25.20 μm,宽2.68~13.86 μm。其他层的叶肉细胞壁呈放射状或不规则状。维管束两侧的叶肉细胞为无色透明的大型梭形细胞,长24.49~78.17 μm,宽5.37~15.01 μm。筇竹叶梭形细胞仅存在于离主脉最近的2个一级平行脉之间,且在叶缘的部位没有观察到梭形细胞的分布。赵惠如[11]等认为梭形细胞的有无、大小与叶肉细胞的分化程度有关。但有学者根据梭形细胞的位置特点,推测其可能与叶肉细胞的水分贮存和运输有关;也有学者根据其无色透明的特点,认为梭形细胞有利于光线更好的透进叶肉细胞[12]。
2.1.3 叶脉 禾本科植物的叶脉分为主脉及两侧的平行脉,且由维管束鞘及维管束组成[13]。竹类的叶维管束外围无“花环”构造,因此它是一个C3低光效植物[14]。按照丁雨龙[15]等的命名方法,又可将主脉两侧的平行脉分为一级平行脉,二级平行脉等。筇竹叶脉的维管束鞘通常由2层细胞组成:外层为较大的薄壁细胞,长5.68~13.74 μm,宽2.70~10.09 μm;内层为厚壁细胞,长3.11~11.64 μm,宽1.83~8.19 μm。叶片以主脉的维管束发育最为完善,越细脉维管束的结构越简化,在二级平行脉及叶缘处的维管束往往无后生导管的发育,仅剩下孔径较小的管胞和筛胞。
主脉维管束往往与许多小维管束交织在一起形成一个复合的维管系统(图1-A1,图2-A1,图2-A2,图2-A3,图2-A4),这些小维管束具有完备的厚壁内鞘和木质部及韧皮部的分化,主脉外的维管束鞘细胞排列不规则,外层薄壁细胞大小不一,且往往不成完整的鞘状,内层是由 1~2 层排列不太整齐的厚壁细胞构成。
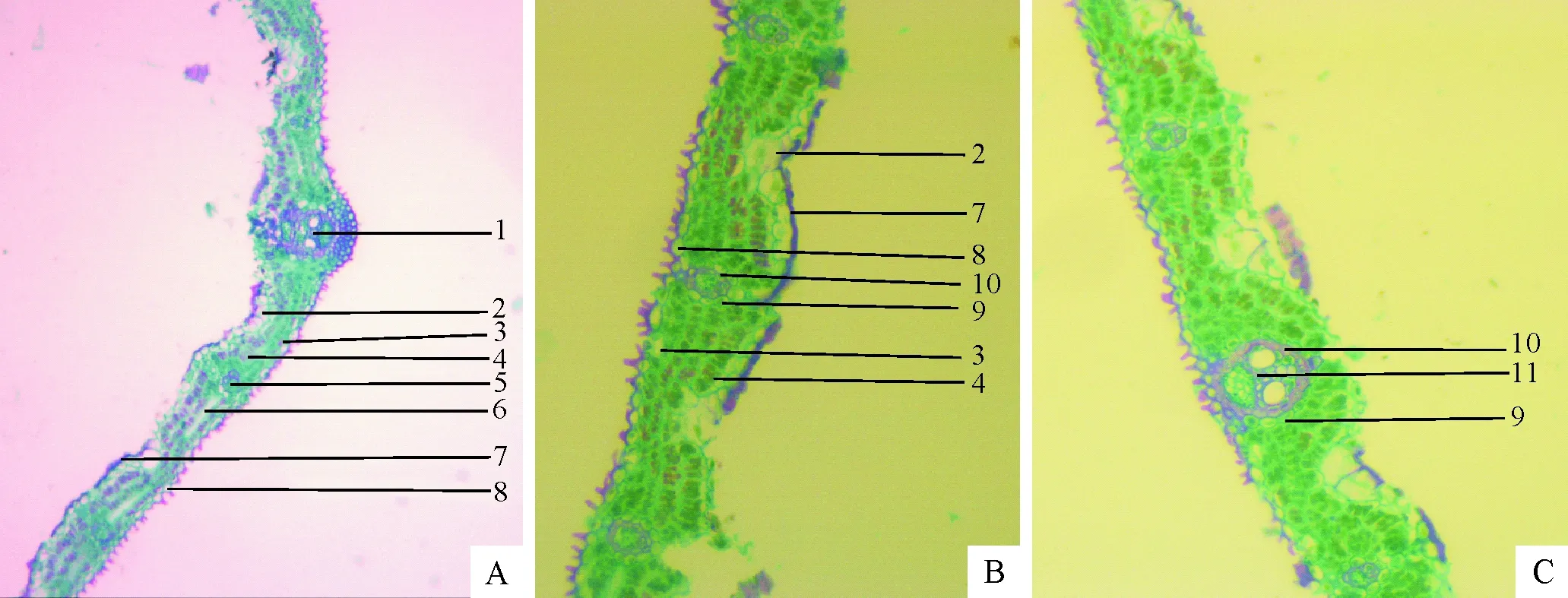
注:(A):筇竹叶片横切面,10X;(B)(C):筇竹叶片横截面,40X;1-主脉维管束;2-泡状细胞;3-气孔;4-叶肉;5-二级平行脉;6-梭形细胞;7-上表皮;8-下表皮;9-薄壁细胞;10-厚壁细胞;11-一级平行脉。
图1筇竹叶片解剖结构
Fig.1 Leaves anatomical structure ofQ.tumidinoda
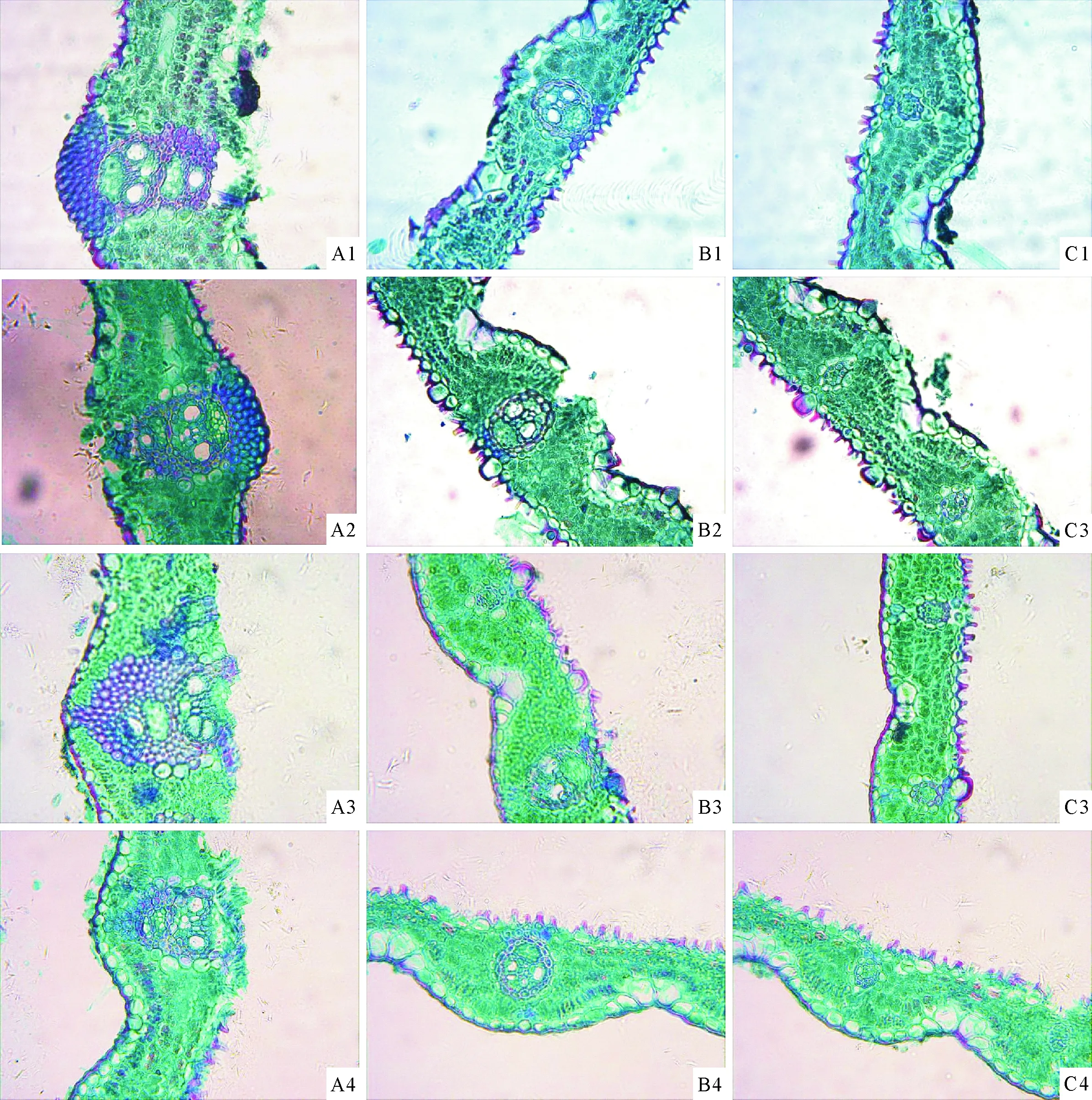
注:A1、B1、C1分别为1年生秆龄筇竹叶片主脉维管束、一级平行脉、二级平行脉;A2、B2、C2分别为2年生秆龄筇竹叶片主脉维管束、一级平行脉、二级平行脉;A3、B3、C3分别为3年生秆龄筇竹叶片主脉维管束、一级平行脉、二级平行脉;A4、B4、C4分别为4年生秆龄筇竹叶片主脉维管束、一级平行脉、二级平行脉。
图2不同秆龄筇竹叶片解剖结构
Fig.2 Leaves anatomical structure ofQ.tumidinodawith different stalk ages
一级平行脉有韧皮部、原生木质部、次生木质部导管等结构(图1-C1,图2-B1,图2-B2,图2-B3,图2-B4),韧皮部靠近远轴面。维管束外被2层鞘细胞包围,外层为薄壁细胞,内层为厚壁细胞,维管束鞘细胞排列规则整齐。
二级平行脉木质部没有大型的导管,韧皮部也较小(图1-A5,图2-C1,图2-C2,图2-C3,图2-C4)。外层薄壁鞘细胞比内层厚壁鞘细胞发达,束外两层维管束鞘细胞排列规则整齐。
叶脉维管束木质部导管及韧皮部的排列,靠近上表皮的近轴面为原生导管,两侧的后生导管和原生导管呈“V”形排列,后生木质部往往在同一侧分化出两个大小相等的导管。韧皮部有大的筛管和小的伴胞组成。无论主脉及次级平行脉维管束的两侧均有厚壁组织与两侧的表皮相连,但以靠近下表皮的厚壁组织较为发达,数量多且木质化程度高。这些结构与王曙光[16]等描述的云南箭竹叶片结构相似。
2.2 筇竹叶解剖结构分析
2.2.1 筇竹种群内叶片解剖变异分析 通过对筇竹叶片解剖指标分析(表1)可知,筇竹种群内叶片除叶肉细胞层数外,其余指标的变异系数均超过15.68%,说明筇竹叶形态在种群内遗传多样性比较丰富。其中,变异系数最大的是叶肉细胞宽度,达26.08%,变异系数最小的是叶肉细胞层数,仅有9.30%,说明叶肉细胞宽度受环境影响较大,而叶肉细胞层数受环境影响较小,遗传相对稳定。
2.2.2 筇竹叶片不同细胞长宽回归分析 为了研究筇竹叶片各类细胞长宽之间的相关关系,对筇竹表皮细胞、泡状细胞、叶肉细胞、薄壁细胞、厚壁细胞的长与宽进行线性回归分析(表2)。
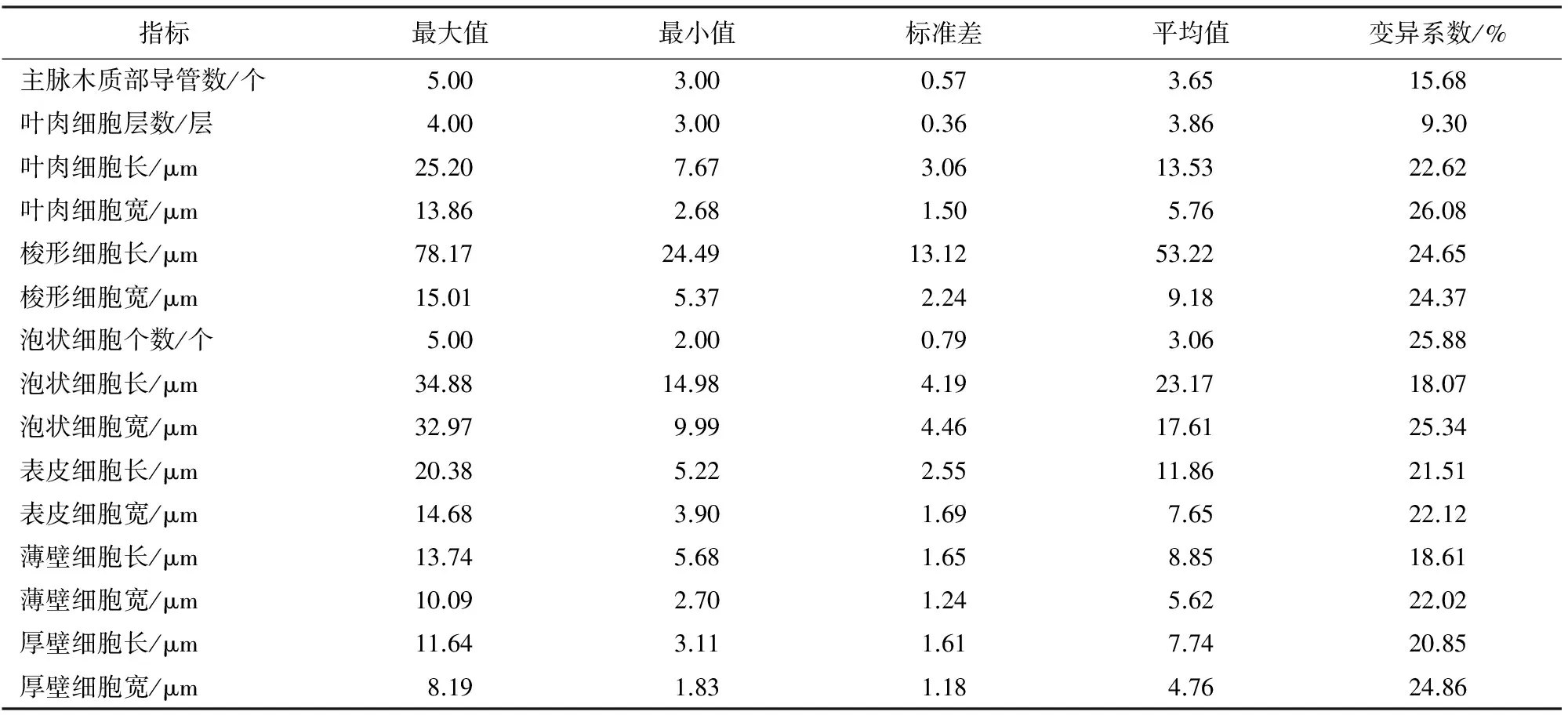
表1 筇竹种群内叶指标变异系数

表2 不同细胞长宽线性回归方程
经F检验,F分布的显著概率全为0.000,说明表皮细胞宽、泡状细胞宽、叶肉细胞宽、薄壁细胞宽、厚壁细胞宽作为自变量,分别与表皮细胞长、泡状细胞长、叶肉细胞长、薄壁细胞长、厚壁细胞长作为因变量时存在高度相关的线性关系。由表2也可以看出,除叶肉细胞长与叶肉细胞宽相关性偏低,其余细胞长宽之间具有较高相关性。
表皮细胞长与表皮细胞宽之间的回归方程为:
Y=X+4.208,R=0.664
泡状细胞长与泡状细胞宽之间的回归方程为:
Y=0.731X+10.229,R=0.779
叶肉细胞长与叶肉细胞宽之间的回归方程为:
Y=0.621X+9.951,R=0.305
薄壁细胞长与薄壁细胞宽之间的回归方程为:
Y=0.813X+4.281,R= 0.611
厚壁细胞长与厚壁细胞宽之间的回归方程为:
Y=0.757X+4.139,R= 0.555
在研究中,可用各类细胞的宽度对各类细胞的长度进行预测。
2.2.3 不同秆龄筇竹叶片解剖结构分析 筇竹叶片的更新是周期性的,通常为1 次/a,可以通过竹叶更新时遗留的小枝痕数来判断竹株的年龄。为了研究不同秆龄筇竹叶片之间是否存在差异,对不同秆龄的筇竹叶片细胞进行多重比较,结果见表3。
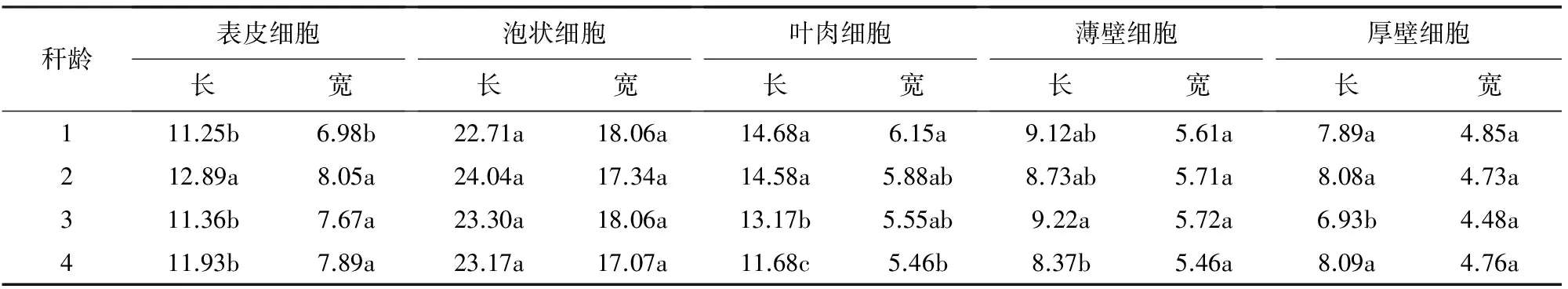
表3 不同秆龄筇竹叶片细胞差异比较
注:同一列里标有相同字母的平均值在a=0.05水平上差异不显著。
由表3可知,不同秆龄的泡状细胞长度、泡状细胞宽度、薄壁细胞宽度及厚壁细胞宽度差异不显著。不同秆龄的表皮细胞长度、表皮细胞宽度、叶肉细胞长度、叶肉细胞宽度、薄壁细胞长度、厚壁细胞长度中,仅表皮细胞长度秆龄2年生竹与1、3、4年生竹差异显著;表皮细胞宽度秆龄1年生竹与2、3、4年生竹差异显著;叶肉细胞宽度秆龄1年生竹与4年生竹差异显著;薄壁细胞长度秆龄3年生竹与4年生竹差异显著;厚壁细胞长度秆龄3年生竹与1、2、4年生竹差异显著;叶肉细胞长度秆龄1年生竹与2年生竹差异不显著,其余年生之间均有显著差异。根据数据比较,多数指标不同年龄之间的差异并不显著。即使有差异也是个别之间的差异,因此,筇竹秆龄并没有对叶片形态特征产生明显差别。
由表3还可知,不同秆龄筇竹叶片细胞变化规律,表皮细胞长宽均为秆龄2年生>4年生>3年生>1年生;泡状细胞长度为秆龄2年生>3年生>4年生>1年生;泡状细胞宽度为秆龄1年生≥3年生>2年生>4年生;叶肉细胞长宽均为秆龄1年生>2年生>3年生>4年生;薄壁细胞长度为秆龄3年生>1年生>2年生>4年生;薄壁细胞宽度为秆龄3年生>2年生>1年生>4年生;厚壁细胞长度为秆龄4年生>2年生>1年生>3年生;厚壁细胞宽度为秆龄1年生>4年生>2年生>3年生。叶肉细胞中含大量叶绿体,叶片进行光合作用的主要部分。叶绿素含量决定植物对太阳辐射的利用率[17],从不同秆龄的叶肉细胞长宽变化规律看出,1、2年生筇竹光合作用能力明显高于3、4年生竹,因此,在筇竹无性系繁殖造林中,最好选用1、2年生新竹,有利于提高筇竹存活率。
3 结论与讨论
植物叶的形态结构,在不同的环境条件下会表现出差异,这是植物对环境适应的结果,从植物组织结构的研究中可以了解植物对特定环境的适应性[18]。
本研究通过石蜡切片,对筇竹的叶片的显微结构进行了分析,发现筇竹叶片表皮细胞长5.22~20.38 μm,宽3.90~14.68 μm;泡状细胞长14.98~34.88 μm,宽9.99~32.97 μm;叶肉细胞长7.67~25.20 μm,宽2.68~13.86 μm;薄壁细胞长5.68~13.74 μm,宽2.70~10.09 μm,厚壁细胞长3.11~11.64 μm,宽1.83~8.19 μm。筇竹叶片整体显微结构与其他竹类叶片结构差异不大,在种的分类上没有多少参考意义,只有找出各种竹类叶间结构稳定的差异性,才是对竹类系统分类有价值的参考依据。
对筇竹叶片显微结构统计分析发现,种群内筇竹叶片在显微结构上存在变异,各指标变异系数范围为9.30%~26.08%,除个别指标外,变异系数均在15%以上。
构建了筇竹叶片各类细胞长宽之间的回归方程,表皮细胞长与表皮细胞宽之间的回归方程为Y=X+4.208,R=0.664;泡状细胞长与泡状细胞宽之间的回归方程Y=0.731X+10.229,R=0.779;叶肉细胞长与叶肉细胞宽之间的回归方程Y=0.621X+9.951,R=0.305;薄壁细胞长与薄壁细胞宽之间的回归方程Y=0.813X+4.281,R=0.611;厚壁细胞长与厚壁细胞宽之间的回归方程Y=0.757X+4.139,R=0.555,可用各类细胞的宽度对各类细胞的长度进行预测。
对不同秆龄的筇竹叶片细胞进行多重比较,发现仅少数指标之间存在差异,多数指标不同年龄之间的差异并不显著,筇竹秆龄并没有对叶片形态特征产生明显差别。1、2年生筇竹光合作用能力明显高于3、4年生竹,因此,在筇竹无性系繁殖造林中,最好选用1、2年生新竹,有利于提高筇竹存活率。
筇竹叶片的更新是周期性的,通常为1 次/a,种群内叶片显微结构存在变异,说明筇竹叶对生境变化反应灵敏,形态可塑性大,对环境适应能力强,遗传多样性比较丰富,原因可能是形态特征是基因与环境共同作用的产物。这些变异都将有利于筇竹的保护和开发利用,本文丰富了筇竹研究的基础信息,但仅对筇竹叶片进行了研究,在实践中还需要对筇竹其他营养器官的变异进行系统的研究。
参考文献:
[1] 董文渊,黄宝龙,谢泽轩,等.筇竹生长发育规律的研究[J].南京林业大学学报:自然科学版,2002(3):43-47.
DONG W Y,HUANG B L,XIE Z X,etal.The study on the growth and development rhythm ofQiongzhueatumidinoda[J].Journal of Nanjing Forestry University:Natural Sciences Edition,2002(3):43-47.(in Chinese)
[2] 董文渊,黄宝龙,谢泽轩,等.筇竹开花结实特性的研究[J].南京林业大学学报:自然科学版,2001,25(6):30-32.
DONG W Y,HUANG B L,XIE Z X,etal.Studies on the characteristics of blossoming and seed bearing ofQiongzhueatumidinoda[J].Journal of Nanjing Forestry University:Natural Sciences Edition,2001,25(6):30-32.(in Chinese)
[3] 董文渊,黄宝龙,谢泽轩,等.筇竹种子特性及实生苗生长发育规律的研究[J].竹子研究汇刊,2002,21(1):58-60.
[4] 何文志,王曙光,邓琳,等.元江箭竹维管束及韧皮部纤维细胞壁发育过程[J].西北林学院学报,2017(3):225-232.
HE W Z,WANG S G,DENG L,etal.Developmental process of the vascular bundles and phloem fiber wall inFargesiayuanjiangensis[J].Journal of Northwest Forestry University,2017(3):225-232.(in Chinese)
[5] 蔡联炳,郭延平.禾本科植物叶片表皮结构细胞主要类型的演化与系统分类和发育途径的探讨(续)[J].西北植物学报,1996(1):65-72.
[6] 曲波,朱明星,陈旭辉,等.22种禾本科植物叶片泡状细胞形态特征的初步研究[J].西北植物学报,2010,30(8):1595-1601.
QU B,ZHU M X,CHEN X H,etal.Morphological character of bulliform cellsin 22 species of poaceae[J].Acta Botanica Boreali-Occidentalia Sinica,2010,30(8):1595-1601.(in Chinese)
[7] 刘永康,郑和平,徐艳霞,等.冬小麦品种更替过程中旗叶结构与光合作用的关系研究[J].河北农业科学,2013,42(3):10-14.
LIU Y K,ZHENG H P,XU Y X,etal.Relationship between flag leaf structure and photosynthesis of winter wheat cultivars[J].Journal of Henan Agricultural Sciences,2013,42(3):10-14.(in Chinese)
[8] 邹雪娟,李润唐,吴钿,等.3个菠萝品种叶片组织结构及其与抗寒性的关系研究[J].广东农业科学,2012(5):38-41.
ZOU X J, LI R T,WU T,etal.Studies on the relationship between leaf tissue structure and cold resistance of 3 pineapple cultivars[J].Guangdong Agricultural Sciences,2012(5):38-41.(in Chinese)
[9] 代勋,王磊,汤家鑫,等.筇竹生物学研究进展[J].世界竹藤通讯,2011(5):34-36.
DAI X,WANG L,TANG J X,etal.Research advance of biology ofQiongzhueatumidinoda[J].World Bamboo and Rattan,2011(5):34-36.(in Chinese)
[10] 杨淑敏,江泽慧,任海青.竹类解剖特性研究现状及展望[J].世界竹藤通讯,2006,4(3):1-6.
YANG S M,JIANG Z H,REN H Q.Present status and advances in anatomical characteristics of bamboo[J].World Bamboo and Rattan,2006,4(3):1-6.(in Chinese)
[11] 赵惠如,龚祝南.竹类叶片的内部解剖与系统演化[J].南京师大学报:自然科学版,1995,18(4):102-108.
[12] 龙春玲,杨光耀,刘腾飞,等.厚壁毛竹与毛竹叶片的光学解剖结构比较分析[J].安徽农业大学学报,2015,42(1):39-44.
LONG C L,YANG G Y,LIU T F,etal.Comparative anatomy of leaves betweenPhyllostachysedulis‘Pachyloen’ andPhyllostachysedulis[J].Journal of Anhui Agricultural University,2015,42(1):39-44.(in Chinese)
[13] 曹慧娟.植物学[M].2版.北京:中国林业出版社,2003:116-128.
[14] 马玉栋,苟光前,孟文艺,等.贵州特有、极危竹种——爬竹、小蓬竹的解剖学研究[J].山地农业生物学报,2012(1):20-24.
[15] 丁雨龙,赵奇僧.竹叶结构的比较解剖及其对系统分类的评价[J].南京林业大学学报,1994,18(3):1-6.
[16] 王曙光.云南箭竹的生物学特性研究[D].南京:南京林业大学,2009.
[17] 俞欣妍,刘健,余坤勇,等.叶片—冠层尺度的毛竹林分光谱特征[J].西北林学院学报,2017(3):7-13.
YU X Y,LIU J,YU K Y,etal.Spectral scale differences between leaf and canopy of spectral characteristics ofPhyllostachyspubescensforest[J].Journal of Northwest Forestry University,2017(3):7-13.(in Chinese)
[18] 刘济明,李丽霞,赵晓鹏,等.小蓬竹不同天然居群解剖形态差异[J].江苏农业科学,2016,44(5):231-234.
猜你喜欢
蔬菜(2022年12期)2022-01-01
星星·散文诗(2021年34期)2021-12-21
河北农业大学学报(2021年5期)2021-11-10
新疆农业科学(2020年9期)2020-10-13
河南农业科学(2019年9期)2019-09-24
农村百事通(2019年1期)2019-01-30
中国全科医学(2017年14期)2017-05-24
中国医药指南(2017年17期)2017-01-16
中国医学影像学杂志(2015年11期)2015-12-28
中国肿瘤临床(2015年17期)2015-12-21
