
一株食窦魏斯氏菌的分离鉴定及其质粒的序列分析
2018-05-01 18:11张广峰张楠笛赵婷婷向文良
食品工业科技 2018年8期
张广峰,张楠笛,赵婷婷,许 琴,谭 霄,向文良
(西华大学食品与生物工程学院,四川省食品生物技术重点实验室, 古法发酵(酿造)生物技术研究所,四川成都 610039)
魏斯氏菌属于乳酸菌,广泛分布于各类发酵食品、土壤等环境中。不仅具有β-葡糖苷酶活性[1]、产细菌素能力[2-3]和较好的抗真菌性[4-5],而且产生的乳酸和胞外多糖对食品的风味有一定影响[6]。故近年来,魏斯氏菌在微生物学、医药学、发酵工业、食品加工等方面的应用受到研究者的广泛关注[7-8]。在以往得到的研究成果中,乳酸菌的诸多生理功能都与其携带的质粒相关,如产胞外多糖、产细菌素、抗生素抗性等[9]。因此对质粒生物学和遗传学的研究受到了极大关注。
许多魏斯氏菌拥有一个或多个天然质粒,但是相对于乳杆菌属、乳球菌属、肠球菌属等质粒,对魏斯氏菌质粒研究较少[9-10]。Park[11]、金红星[12]、Kim[13]等分别从食窦魏斯氏菌(Weissellacibaria)分别分离出3、2、6个质粒,王海娟等[14]从融合魏斯氏菌(Weissellaconfusa)中分离出了2个质粒,并对部分质粒完成了序列分析及载体构建。数据库中魏斯氏菌质粒的相关数据较少,以致于不能推断质粒阅读框中的具体功能[13]。所以丰富魏斯氏菌质粒数据库对魏斯氏菌属质粒复制、遗传、进化、功能等方面具有重要意义。
中国泡菜中蕴含丰富的乳杆菌质粒资源[15],因此本研究以中国四川泡菜为分离源,筛选其中含质粒乳酸菌并对质粒进行全序列测定及功能分析,以期为构建良好的载体提供理论基础。
1 材料与方法
1.1 材料与仪器
泡菜液 四川盈宇食品有限公司;大肠杆菌(EscherchiacoliDH5α) 天根生化科技(北京)有限公司;pGEM-T Simple Vector 普洛麦格(北京)生物技术有限公司;溶菌酶、限制性核酸内切酶 美国Thermo scientific;Taq DNA聚合酶、T4 DNA连接酶 Promega有限公司;质粒小量提取试剂盒 OMEGA公司;胶回收试剂盒 天根生化科技有限公司;土壤DNA Extraction试剂盒 福际生物技术有限公司;LB培养基、LB液体培养基、MRS液体培养基和改良MRS培养基(含0.75 g/100 mL的CaCO3) 均按文献[16]配制。
Biometra PCR仪 德国耶拿分析仪器有限公司;Allegra X-15R冷冻离心机 贝克曼库尔特公司;720BR电泳凝胶成像分析仪 Bip-rad(美国);SGSP-02恒温隔水式培养箱 黄石恒丰器械有限公司;DYY-8C型电泳仪 北京市六一仪器厂。
1.2 实验方法
1.2.1 乳酸菌的分离纯化 取25 mL泡菜液于225 mL灭菌生理盐水中混合均匀,经10倍梯度稀释后涂布于改良MRS 培养基中,37 ℃倒置培养48 h。挑取周围形成透明圈的菌落划线纯化,同时进行革兰氏染色和过氧化氢酶实验。将革兰氏染色结果为阳性、过氧化氢酶阴性的菌株初步判定为乳酸菌[17]。
1.2.2 质粒DNA的提取 将纯化的乳酸菌接种至MRS液体培养基,37 ℃过夜培养。采用质粒小量提取试剂盒提取质粒DNA,经过以下修改:菌体以转速12000 r/min离心2 min、ddH2O洗涤后重悬于含有30 mg/mL溶菌酶的SolutionI中,37 ℃温育1 h,其余操作按照说明书进行。1%琼脂糖凝胶电泳检测提取质粒DNA。
1.2.3 含质粒乳酸菌的鉴定 乳酸菌的生化特征分析按照《乳酸菌细菌分类鉴定及实验方法》[18]进行。参照文献[19]方法提取含质粒乳酸菌基因组DNA。16S rDNA的PCR扩增引物为Eu27F(5′-AGAGTTTGATCCTGGCTCAG-3′)和1492R(5′-GGTTACCTTGTTACGACTT-3′)。扩增条件:95 ℃预变性5 min,95 ℃变性1 min,50 ℃退火1 min,72 ℃延伸2 min,30个循环,72 ℃保持10 min。扩增产物连接到pGEM-T后转化到E.coliDH5α中,筛选阳性克隆子对16S rDNA测序。所得序列提交至NCBI进行同源性比对,利用MEGA 5.0软件构建系统发育树。
1.2.4 质粒序列测定 根据质粒条带,回收其中的大质粒命名为pA010。分别利用不同的限制性内切酶EcoR I、BamH I和PstI对分离纯化的质粒pA010进行酶切分析,回收酶切反应液。参照都立辉等[17]方法补平酶切后的黏性末端并在3′端引入碱基A,回收其中DNA片段同pGEM-T载体连接并转化到E.coliDH5α中,筛选阳性克隆测序。以测序结果中最长序列片段为基础设计引物 W1CF(5′-TTTTCCACTTCGTTATTCTTG-3′)和 W1CR(5′-TTGACAACGACAAGTATTAGAGC-3′)扩增质粒pA010剩余片段。PCR扩增条件:95 ℃ 5 min,95 ℃ 1 min、50 ℃ 1 min、72 ℃ 2 min、30 个循环,72 ℃ 10 min。将PCR扩增片段与载体连接、转化到E.coliDH5α,挑选阳性克隆测序,拼接各个测定序列获得质粒pA010的全序列。
1.2.5 质粒的生物信息学分析 利用NCBI网站上的ORF Finder(Open reading frame finder)寻找该质粒可能存在的开放阅读框(ORF),然后对质粒的核苷酸及氨基酸序列进行分析。DNAman6.0绘制质粒图谱,CDD网站用于假定蛋白保守结构域的寻找,CLUSTAL X实现序列的多重比对。通过分析质粒上的序列信息,推导质粒复制方式,并将该质粒全序列提交GenBank数据库。
2 结果与分析
2.1 含质粒菌株的分离鉴定
从四川泡菜中分离到11株含溶钙圈的菌株,编号为W1~W11,均呈乳白色、边缘整齐的圆形菌落,油镜下观察显示菌体为杆状、紫色。对这11株菌进行质粒普查发现只有菌株W5携带2个天然质粒(图1)。将其中大质粒命名为pA010,分离纯化该质粒,通过酶切和PCR扩增对其进行序列测定。

图1 四川泡菜中乳酸菌的质粒提取电泳图Fig.1 Plasmid extracts electrophoresis of Lactic acid bacteria in Sichuan Pickle注:M为DNA Marker;1~11为乳酸菌W1~11的质粒提取液。
对含质粒菌株W5进行常规生理生化实验,结果如表1。根据形态学和生化特征,检索乳酸菌分类系统,初步确定该菌株属于魏斯氏属(Weissella)。提取含质粒菌株W5的基因组DNA并进行16S rDNA扩增,其扩增产物序列大小为1463 bp。将菌株序列与GenBank数据库中标准菌株的序列进行BLAST比对发现,该菌株与食窦魏斯氏菌(W.cibaria)的16S rDNA相似性达99%。该菌株与标准菌株的16S rDNA的系统发育表明,该菌株与食窦魏斯氏菌具有较大的亲缘关系(图2)。一般来讲,在种分类等级上,如果2个分类单位间的16S rDNA序列同源性大于97.5%,则认为属于同一个种[20]。结合菌株的形态学和生理生化特征,这株含内源质粒的乳酸菌被鉴定为食窦魏斯氏菌。

图2 含质粒菌株W5基于16 S rDNA序列的系统发育树Fig.2 Phylogenetic tree of strain W5 containing plasmid based on 16S rDNA sequence

表1 含质粒菌株W5部分生理生化特征Table 1 Part of physiological and biochemical results of strain W5 containing plasmid
2.2 质粒全序列测定
将回收纯化的质粒pA010分别用限制性内切酶EcoR I、BamH I和PstI进行酶切分析,发现该质粒仅有4个EcoR I酶切位点(图3)。回收酶切反应液,末端补平后连入pGEM-T,经测序分析EcoR I酶切的4个片段长度分别为5762、2820、2677、371 bp。以最长序列片段为基础设计引物,对pA010进行PCR扩增,以确定各个质粒片段的连接顺序,最终得到pA010的完整序列(GenBank登录号为MF951102)。

图3 质粒pA010酶切分析Fig.3 Restriction enzyme analysis of pA010注:M为DNA Marker;E、B、P分别代表EcoR I、BamH I和Pst I酶切pA010。
2.3 质粒pA010的一般结构分析
食窦魏斯式菌W5较大质粒pA010序列大小为11630 bp,为一个闭合环状双链DNA分子,其中GC含量34.6%。通过NCBI网站上的ORF Finder软件查找ORF,搜索到12个序列长度大于300 bp的ORFs。由于一个具有起始密码子和终止密码子的ORF不一定都是蛋白质的编码序列,因此搜索到的ORFs中还有非编码基因和重复序列,所以再将序列进行BLASTp蛋白质同源片段分析[21]。12个ORFs中有4个ORFs编码已知功能蛋白,其他8个ORFs编码假定蛋白(表2)。通过DANman6.0查找质粒的EcoR I、BamH I和PstI位点发现,质粒中无BamH I和PstI的酶切位点、4个EcoR I酶切位点,此结果与图4一致。为标识已知功能蛋白、假定蛋白和EcoR I酶切位点在质粒中的位置,通过DANman6.0构建质粒图谱,如图4所示。

表2 质粒pA010推断的ORFs编码的蛋白质Table 2 The putative ORFs and the encoded proteins of plasmid pA010

图4 食窦魏斯式菌W5中pA010的质粒图谱Fig.4 Plasmid map of pA010 from W. cibaria W5
2.4 质粒pA010的功能分析
ORF2编码了一个INT_C_like_3,与植物乳杆菌中酪氨酸重组酶显示了99%的相似性,与乳酸片球菌质粒pSMB74中的酪氨酸重组酶显示了95%的相似性。这个蛋白属于DNA breaking/re-joining enzymes超家族,可能与质粒多聚体的解离有关[22]。Motlagh A等[23]表明,这个蛋白经常被发现在乳酸菌的θ型复制质粒。
ORF3编码复制蛋白Rep,与植物乳杆菌G63内源质粒pG6302复制蛋白的相似性达100%,与植物乳杆菌内源质粒pLME300复制蛋白的相似性达93%。这些同源Rep有一个保守的涉及质粒复制起始的Rep_3结构域(pfam01051),这个结构域属于RepA_N质粒家族,一般为θ型复制质粒所有[15]。而且pLME300[24]和pG6302[25]被认为是一种θ型复制的质粒,所以pA010被认为是一种θ型复制的质粒。复制蛋白的上游515 bp有类似起始质粒复制的重复序列,预测可能是复制起始区(图5)。复制子区域包含3个同向重复序列(Direct repeats,DR)和3个反向重复序列(Invert repeats,IR),还有一段富含A、T序列。Xi等[26]认为正向重复序列是复制蛋白的目标且在质粒的不相容性上有重要作用,而且A/T富含区被认为是复制的起始。
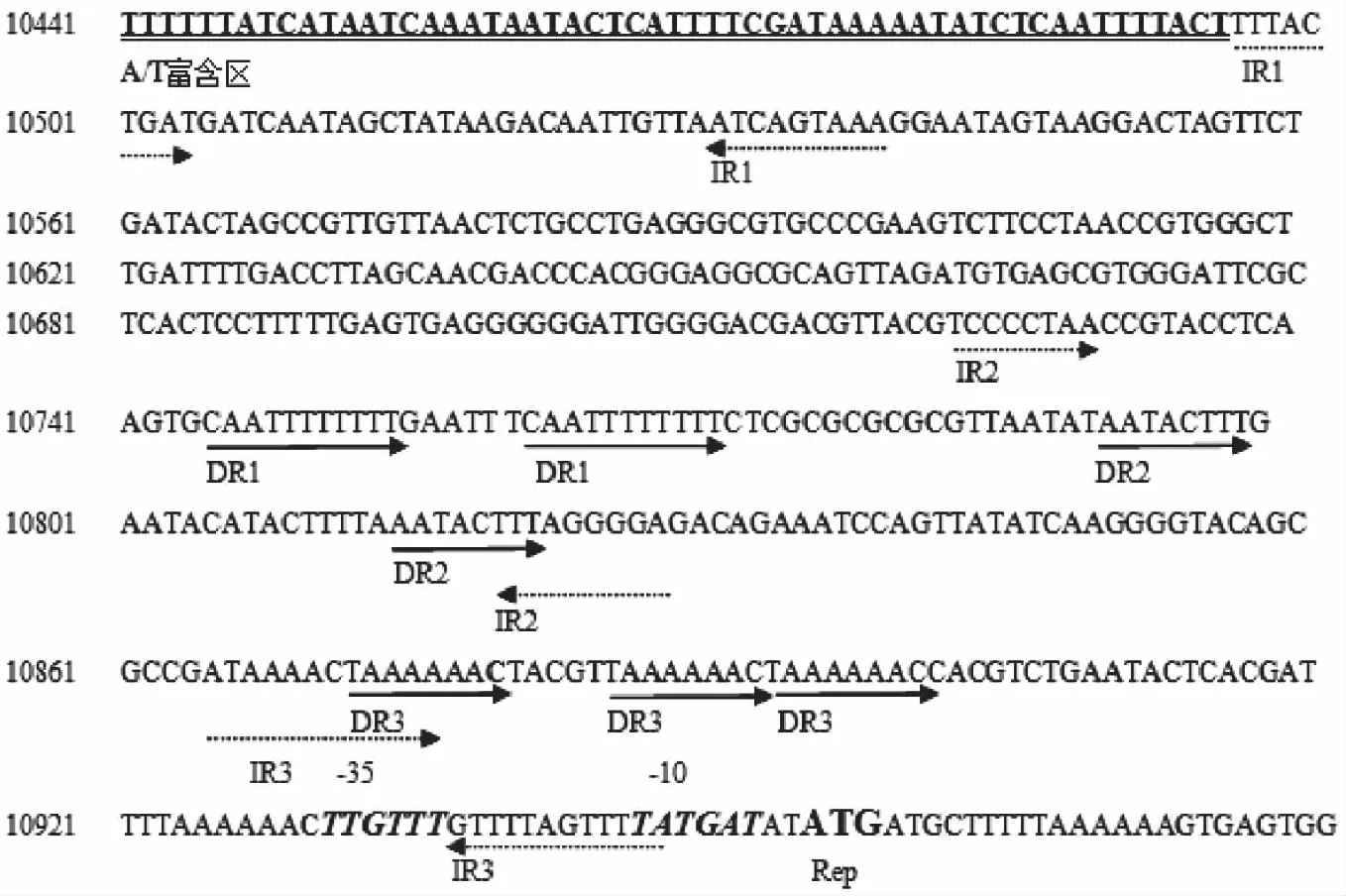
图5 质粒pA010假定的复制起始区Fig.5 The predicted origin region of replication of pA010注:双下划线为A/T富含区;实线箭头为正向重复(DR);虚线箭头为反向重复(IR);加粗斜体字母为-10和-35区;加粗ATG为复制蛋白(Rep)的起始点。
ORF11和ORF12编码的蛋白分别与乳酸片球菌H中质粒pSMB74的PemK和PemI蛋白显示了100%和94%的相似性。Fang等[25]报道,PemK和PemI是一个II型毒素和抗毒素系统(type II Toxin and Antitoxin system,II-TA),其中PemK编码稳定的毒素,PemI编码不稳定的抗毒素。ORF11和ORF12之间重叠7个碱基,提示两个基因可能共同构成一个操纵子(图6)。细菌质粒编码的TA系统作为沉溺系统(plasmid addiction systems,PAS),通过细胞分裂后死亡的机制维持质粒的稳定性[27]。而且TA系统利于细菌持留感染的形成(细菌进入一种低生长或休眠状态),在这种状态下细胞对周围环境的适应力可以得到很大提升,如对抗生素的耐药增加、在有害环境下保持一部分生存个体等[28]。在植物乳杆菌NC7中的质粒p256[29]、植物乳杆菌CD033中质粒pCD033[26]和唾液乳杆菌UCC118中的质粒pSF118[27]中都编码了一个TA系统。近年来,乳酸菌中TA研究受到了相当的重视,不仅是因为质粒携带的TA基因维持细胞质粒的遗传稳定性,更是由于乳酸菌中TA的研究对乳酸菌中菌株的多样化的元基因组分析以及乳酸菌开发有重要意义[30]。

图6 pemI、pemK在pA010上的遗传结构及其蛋白序列同源性比对 Fig.6 Genetic structure and sequence alignment of the pemI/pemK locus in pA010注:A为pemI/pemK遗传结构示意图,箭头所示pemI和pemK的转录方向;B为pemI编码产物与PemI蛋白序列比对; C为pemK编码产物与PemK蛋白序列比对,pSMB74、p256、pSF118、pCD033分别表示P. acidilactici H、L. plantarum NC7、L. salivarius UC118、L.plantarum CD033中的质粒。
3 结论
本实验以四川泡菜为分离源,从中筛选到一株含质粒的食窦魏斯氏菌。其中大质粒pA010全序列结果显示该质粒全长11630 bp,GC含量为34.6%,同时编码了四个可识别功能蛋白:酪氨酸重组酶、复制蛋白、抗毒素PemI和毒素PemK。通过分析这些蛋白的功能发现,该质粒可能是θ型复制质粒,而且毒素-抗毒素系统对质粒稳定以及魏斯氏菌耐受周围环境提供了可能。
[1]Lee K W,Han N S,Kim J H. Purification and characterization of beta-glucosidase fromWeissellacibaria37[J]. Journal of Microbiology & Biotechnology,2012,22(12):1705-1713.
[2]Li S W,Chen Y S,Lee Y S,et al. Comparative genomic analysis of bacteriocin-producingWeissellacibaria110[J]. Applied Microbiology and Biotechnology,2017,101(3):1-11.
[3]王筱梦,江芸,孙芝兰,等. 1株产细菌素乳酸菌的鉴定及所产细菌素的诱导合成现象[J]. 食品科学,2016,37(21):170-175.
[4]Baek E,Kim H,Choi H,et al. Antifungal activity ofLeuconostoccitreumandWeissellaconfusain rice cakes[J]. J Microbiol,2012,50(5):842.
[5]常伟,饶瑜,胡怀容,等. 一株抗真菌食窦魏斯氏菌AT6的特性分析及培养条件优化[J]. 食品工业科技,2014,35(6):193-196.
[6]Galle S,Schwab C,Arendt E,et al. Exopolysaccharide-formingWeissellastrains as starter cultures for sorghum and wheat sourdoughs[J]. Journal of Agricultural & Food Chemistry,2010,58(9):5834.
[7]王海娟,戴雨珂,潘渠. 魏斯氏菌的研究现状[J]. 成都医学院学报,2014,9(6):747-750.
[8]Lee K W,Park J Y,Jeong H R,et al. Probiotic properties ofWeissellastrains isolated from human faeces[J]. Anaerobe,2012,18:96-102.
[9]孙大庆,李洪飞,宋大巍,等.乳杆菌属天然质粒研究进展[J]. 食品科学,2015,36(11):251-255.
[10]孙大庆,李洪飞,杨健. 植物乳杆菌编码复制起始蛋白天然质粒的系统进化分析[J]. 微生物学通报,2017,44(5):1047-1055.
[11]Park M S,Kim S H,Kim J D,et al. Molecular characterization of plasmid DNA fromWeissellacibariaisolated from Kimchi[J]. Korean Journal of Genetics,2007,29(1):29-35.
[12]金红星,杨希寅,成文玉. 辣白菜中分离含内源性质粒的魏斯氏菌及其鉴定[J]. 中国酿造,2012,31(1):77-79.
[13]Kim S Y,Oh C G,Lee Y J,et al. Sequence analysis of a cryptic plasmid pKW2124 fromWeissellacibariaKLC140 and construction of a surface display vector[J]. Journal of Microbiology & Biotechnology,2013,23(4):545.
[14]王海娟,戴雨珂,苏森森,等. 一个融合魏斯氏菌隐蔽质粒的序列分析[J]. 生物技术通报,2016,32(4):217-221.
[15]杨四佳,王颖,李宇婷,等. 泡菜中乳杆菌的快速检出和乳杆菌质粒资源的初步调查[J]. 食品与生物技术学报,2013,32(6):639-644.
[16]杜连祥,路福平. 微生物学实验技术[M]. 北京:中国轻工业出版社,2014:349-354.
[17]都立辉,霍贵成,鞠兴荣,等. 一株植物乳杆菌的分离鉴定及其隐蔽质粒的序列分析[J]. 食品科学,2010,31(17):236-239.
[18]凌代文. 乳酸细菌分类鉴定及实验方法[M]. 北京:中国轻工业出版社,1999:117-127.
[19]Xiang W L,Liang H Z,Liu S,et al. Isolation and performance evaluation of halo tolerant phosphate solubilizing bacteria from the rhizospheric soils of historic Dagong Brine Well in China[J]. World Journal of Microbiology and Biotechnology,2011,27(11):2629-2637.
[20]StackebrandT E,Gebel B M. Taxonomic note:a place for DNA-DNA reassociation and 16S rRNA sequence analysis in the present species definition in bacteriology[J]. International Journal of Systematic Bacteriology,1994,44(4):846-849.
[21]Silvia H,Reingard G,Stefan H. Characterization of theLactobacillusplantarumplasmid pCD033 and generation of the plasmid free strainL.plantarum3NSH[J]. Plasmid,2015,81(11):9-20.
[22]Rob V H,Raphael L,Gipsi L M,et al. Towards a more accurate annotation of tyrosine-based site-specific recombinases in bacterial genomes[J]. Mobile DNA,2012,3(1):6.
[23]Motlagh A,Bukhtiyarova M,Ray B. Complete nucleotide sequence of pSMB 74,a plasmid encoding the production of pediocin AcH inPediococcusacidilactici[J]. Letters in Applied Microbiology,1994,18(6):305.
[24]Gfeller K Y R,Melle L,Teuber M,et al. Sequence and genetic organization of the 19.3-kb erythromycin-and dalfopristin-resistance plasmid pLME300 fromLactobacillusfermentumROT1[J]. Plasmid,2003,50(3):190-201.
[25]Fang F,Flynn S,Li Y,et al. Characterization of endogenous plasmids fromLactobacillussalivariusUCC118[J]. Applied & Environmental Microbiology,2008,74(10):3216-3228.
[26]Xi X,Fan J,Hou Y,et al. Characterization of three cryptic plasmids fromLactobacillusplantarumG63 that was isolated from Chinese pickle[J]. Plasmid,2013,70(3):321-328.
[27]孙瑞,宁德刚. 细菌Ⅱ型毒素-抗毒素系统活性的调控[J]. 微生物学通报,2016,43(12):2714-2719.
[28]楚海荣,付玉荣,伊正君. 细菌毒素-抗毒素系统研究进展[J]. 细胞与分子免疫学杂志,2015,31(2):271-274.
[29]Sørvig E,Skaugen M,Naterstad K,et al. Plasmid p256 fromLactobacillusplantarumrepresents a new type of replicon in lactic acid bacteria,and contains a toxin-antitoxin-like plasmid maintenance system[J]. Microbiology,2005,151(2):421-431.
[30]徐谓,李洪军,贺稚非. 乳酸菌中毒素-抗毒素系统研究进展[J]. 食品科学,2015,36(19):260-264.
猜你喜欢
现代畜牧科技(2021年9期)2021-10-13
江西农业学报(2021年4期)2021-04-20
数学大王·中高年级(2019年12期)2019-12-24
小学阅读指南·低年级版(2016年10期)2016-09-10
恋爱婚姻家庭(2016年18期)2016-07-23
西南医科大学学报(2015年1期)2015-08-22
中国当代医药(2015年9期)2015-03-01
中国洗涤用品工业(2015年7期)2015-02-28
西南军医(2015年6期)2015-01-23
中国酿造(2014年9期)2014-03-11
