
无籽沙糖桔E3泛素连接酶U-box基因的克隆及表达分析
2018-05-04 06:13唐文武莫灿坤吴秀兰
江西农业大学学报 2018年2期
唐文武,莫灿坤,吴秀兰
(肇庆学院 生命科学学院,广东 肇庆 526061)
植物自交不亲和性是部分显花植物为促进异花授粉,防止近亲繁殖而形成的一种遗传机制,它涉及高等植物受精过程中雌蕊和花粉之间的相互作用,是研究细胞间相互识别、信号传递及基因时空表达的一种理想模型。根据其遗传机制可分为配子体、孢子体自交不亲和两种类型,其中配子体自交不亲和现象在蔷薇科、玄参科、茄科等植物中较常见,其自交不亲和反应主要由花粉F-box和花柱中的S-RNase等基因控制[1-2]。泛素/26S蛋白酶体途径是真核生物最重要的蛋白降解途径,参与调控了80%~85%的蛋白质降解过程[3]。泛素/26S途径主要由泛素激活酶E1、泛素结合酶E2、泛素连接酶E3和26S蛋白酶体组成[4],它们会共同作用引起S-RNase等特异蛋白的泛素化并降解,从而影响花粉萌发和花粉管的生长延伸,导致自交不亲和的发生[5]。因此,泛素/26S蛋白酶体途径在植物的自交不亲和中发挥着重要作用。
泛素连接酶E3在底物蛋白特异性识别中起到关键作用,根据其共价键结合方式,可分为HECT、RING-finger和U-box结构域等3种类型[6]。U-box域是植物特有的一类高度保守的功能结构域,由70多个氨基酸残基构成,在细胞发育、信号转导和光周期反应等反面具有重要生理功能。在拟南芥和芸薹属植物中包含U-box域的ARC1和ARM蛋白在自交不亲和反应中发挥重要调节作用[7-8]。无籽沙糖桔是沙糖桔芽变材料中选育出的自交不亲和品种,在生产和科学研究中具有重要价值[9]。苗红霞等[10-11]通过SSH技术从无籽沙糖桔材料中获得一些与泛素/26S蛋白酶体途径相关的非S因子,并筛选到1个含U-box域的EST序列在子房中高表达。本研究以此为基础,利用无籽沙糖桔和普通有籽沙糖桔为试材,分离克隆了沙糖桔U-box基因的cDNA全长序列,并对其DNA与蛋白质序列、表达特性、同源进化关系等开展研究,为研究该基因在柑橘自交不亲和反应中的作用及选育奠定基础。
1 材料与方法
1.1 供试材料
供试的各类沙糖桔材料种植于肇庆市四会果园8 a生果树,于春季盛花期分别对无籽沙糖桔和普通有籽沙糖桔的叶、芽、花丝、花粉、雌蕊、柱头、子房、花柱进行取样,并获取无籽沙糖桔自交授粉、异花授粉(无籽沙糖桔×有籽沙糖桔)后不同发育时期的雌蕊。以上材料摘取后利用液氮速冻后带回实验室,贮藏在-80 ℃冰箱中备用。
1.2 实验方法
1.2.1 总RNA的提取及cDNA合成 总RNA提取按照北京天恩泽的Column Plant RNAout试剂盒说明步骤操作,经12 g/L琼脂糖凝胶电泳检测质量,再利用紫外分光光度仪测定RNA含量。cDNA第一链合成利用M-MLV试剂盒(Life Technology公司)及说明方法进行,DNase I 和RNase 抑制剂购于TAKARA公司。
1.2.2CrU-box基因克隆及生物信息学分析 以SSH文库中筛选出的无籽沙糖桔EST序列为基础[11],参照甜橙基因组(http://citrus.hzau.edu.cn)中登录号为Cs6g21870的U-box基因序列,设计1对引物U-box-F/R(表1)扩增沙糖桔U-box基因的cDNA,其25 μL PCR反应体系为模板cDNA 1μL,10×PCR buffer 2.5 μL,引物各0.5 μL,dNTP 1.0 μL,ExTaq0.25 μL,补ddH2O至25 μL。PCR反应程序如下:94 ℃ 5 min;30个循环包括94 ℃ 45 s,55 ℃ 45 s,70 ℃ 90 s;最后72 ℃再延伸10 min。然后利用TAKARA公司生产的琼脂糖凝胶DNA回收试剂盒回收PCR产物,连接到克隆载体pMD19-T vector,转化至大肠杆菌DH5α,通过PCR及酶切鉴定的阳性克隆送华大基因公司测序。利用SMART软件(http://smart.embl-heidelberg.de/)对CrU-box基因编码蛋白进行保守结构域分析,利用 BLAST工具对CrU-box蛋白进行比对搜索,下载其它植物同源U-box蛋白序列,利用MEGA 5.0 软件构建系统进化树。
1.2.3CrU-box基因的实时荧光定量PCR分析 提取无籽沙糖桔和普通有籽沙糖桔叶、芽、花丝、花粉、雌蕊、柱头、子房、花柱等不同组织器官的总RNA,以及无籽沙糖桔自交授粉、异交授粉(无籽沙糖桔×有籽沙糖桔)后9个不同时段(0,12,24,48,72,96,120,144,168 h)的雌蕊总RNA,并反转录合成cDNA。设计特异性引物qU-box-F/R(表1)以及柑橘actin为内参引物(表1),利用TARAKA公司的SYBR Premix ExTaqII荧光染料试剂盒,TOYOBO 公司生产的7300 Real-time PCR扩增仪进行实时荧光定量PCR。具体实验过程严格按照SYBR Premix ExTaqII说明书进行,反应结束后采用2-ΔΔCT法[12]进行结果分析,计算出CrU-box基因相对表达量,并绘制柱形分析图,统计分析采用one-way ANOVA法,n=3。
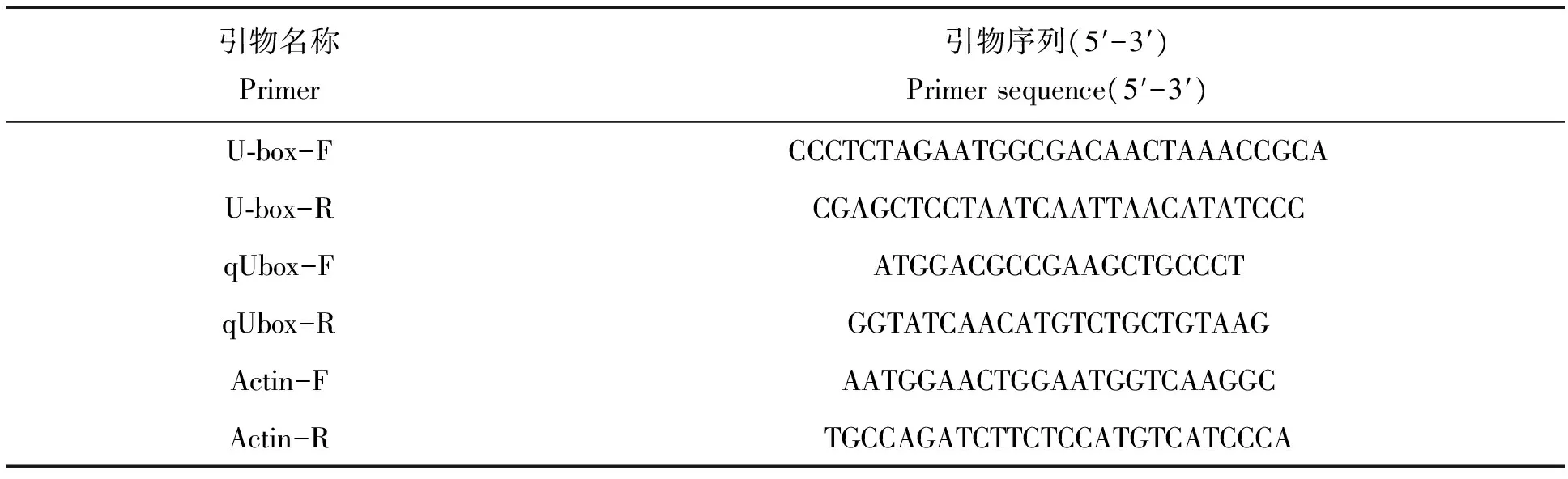
表1 相关引物序列
2 结果与分析
2.1 CrU-box基因克隆
提取无籽沙糖桔和普通有籽沙糖桔的总RNA,经12 g/L琼脂糖凝胶电泳检测显示28S、18S RNA的条带清晰明亮(图1A),且A280/A260值介于1.80~2.0,A260/A230值大于2.0,表明提取的总RNA较完整且无DNA污染。利用U-box-F/R引物扩增无籽沙糖桔和普通有籽沙糖桔的雌蕊cDNA,结果见图1B。由图可知,该特异性PCR引物在无籽沙糖桔和普通有籽沙糖桔中均扩增获得一条清晰的条带,其长度约3 000 bp。
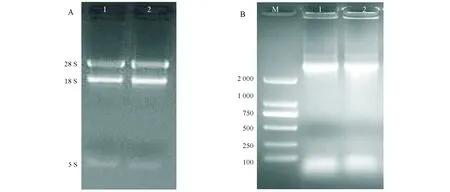
A:总RNA提取;B:PCR扩增结果;1:无籽沙糖桔;2:普通有籽沙糖桔A:Total RNA;B:Production of PCR;1:Wuzishatangju;2:Putongyouzishatangju图1 沙糖桔RNA提取及PCR扩增结果Fig.1 RNA extraction and PCR products of U-box gene in Shatangju
2.2 无籽沙糖桔和普通有籽沙糖桔的CrU-box基因序列分析
将上述PCR扩增产物经回收后克隆并测序,DNA测序结果及蛋白序列见图2、图3。由图2可知CrU-box基因的ORF长度为3 150 bp,预测编码1 049 aa的蛋白质。无籽沙糖桔和普通有籽沙糖桔在ORF区域存在4个bp的差异,分别为第837位的G变为A、1 215位的C变为T、1 837位的A变为T、1 868位的C变为G,并导致第279位的缬氨酸V变为异亮氨酸I、第612位的酪氨酸Y变为苯丙氨酸F(图3)。

WZSTJ为无籽沙糖桔的CrU-box序列;PTYZSTJ为普通有籽沙糖桔的CrU-box序列WZSTJ:CrU-box sequence of Wuzishatangju;PTYZSTJ:CrU-box sequence of Putongwuzishatangju图2 无籽沙糖桔和普通有籽沙糖桔CrU-box基因部分序列比对Fig.2 comparison partial sequence of U-box gene between wuzishatangju and youzishatangju

图3 无籽沙糖桔和普通有籽沙糖桔CrU-box蛋白部分氨基酸序列比对Fig.3 Comparison partial amino acid sequence of U-box protein between wuzishatangju and youzishatangju
2.3 CrU-box结构域及系统进化分析
利用SMART软件对无籽沙糖桔、普通有籽沙糖桔的CrU-box基因编码的蛋白进行保守结构域分析,结果发现CrU-box蛋白的951~1 021位点存在1个U-box保守结构域,该U-box域位于C端,主要修饰RING锌指结构,表明CrU-box属于U-box家族成员之一。利用BLAST工具对CrU-box蛋白开展比对搜索,选取含U-box结构域且相似性较高的11种植物U-box蛋白序列,与普通有籽沙糖桔和无籽沙糖桔的CrU-box蛋白进行序列比对和系统进化树分析,结果见图4。蛋白序列比对结果显示,12种含U-box结构域的同源蛋白间序列相似性均较高(80.7%~99.6%)。其中沙糖桔和甜橙(C.sinensis)相似性最高,达到99.6%;其次与克里曼丁桔(C.clementina)相似性为99.2%;与黄瓜(C.sativus)序列差异最大,但相似性也达80.7%。该结果表明植物U-box蛋白在进化过程中保守,进化上柑橘属植物聚为一类,而榴莲(Duriozibethinus)、雷蒙德氏棉(Gossypiumraimondii)、烟草(Nicatianaattenuata)因同源性较高聚为一类,木薯(Manihotesculenta)、葡萄(Vitisvinifera)、苹果(Malusdomestica)、桃(Prunuspersica)、核桃(Juglansregia)、黄瓜(Cucumissativus)聚为一类。
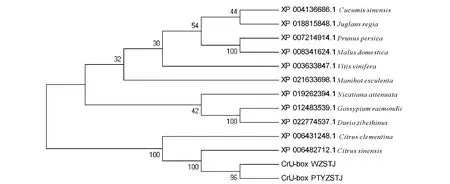
WZSTJ为无籽沙糖桔的CrU-box蛋白;PTYZSTJ为普通有籽沙糖桔的CrU-box蛋白WZSTJ:Protein of Cru-box from Wuzishatangju;PTYZSTJ:Protein of Cru-box from Putongyouzishatangju图4 CrU-box与其它植物U-box蛋白的系统进化分析Fig.4 Phylogenetic tree analysis of U-box protein in plant
2.4 CrU-box基因的时空表达量分析
CrU-box基因在无籽沙糖桔和普通有籽沙糖桔不同器官间的实时荧光定量PCR结果见图5。由图可知,CrU-box基因在无籽和普通有籽沙糖桔的7个不同组织器官均有表达,其中芽、叶、花瓣、花丝、柱头和花柱表达量均较低,而子房表达量远高于其他器官表达量。在无籽沙糖桔中,CrU-box基因在子房中表达量是最低的花瓣表达量的12.5倍(P<0.01);在普通有籽沙糖桔中,CrU-box基因在子房中表达量是最低花瓣表达量的10倍(P<0.01)。由此可知,CrU-box基因在无籽沙糖桔和普通有籽沙糖桔中均表现出在子房中高表达量特征。
由于CrU-box基因在子房中的高表达特征,为进一步了解其在授粉后的表达量水平,本试验研究了自交授粉(无籽沙糖桔×无籽沙糖桔)、异交授粉(无籽沙糖桔×普通有籽沙糖桔)后CrU-box基因在雌蕊器官中不同时间段的表达水平,结果见图6。由图可知,CrU-box基因在自交授粉和异交授粉后的0~96 h表达量逐渐增加,96 h表达量达到最大值,96~168 h表达量逐渐下降。但在授粉后48,72,96 h时间段上,自交授粉的CrU-box基因相对表达量远高于异交授粉后的表达量;尤其是无籽沙糖桔自交授粉后的96 h表达量远高于异交授粉后96 h相对表达量,且自交授粉96h表达量是异交授粉的4.5倍(P<0.01)。因此,笔者初步推测无籽沙糖桔自交授粉后CrU-box基因的高表达水平,可能激活泛素分子,导致S-RNase等特异蛋白的泛素化及降解,从而引起花粉管的延伸受阻,导致自交不亲和的发生。
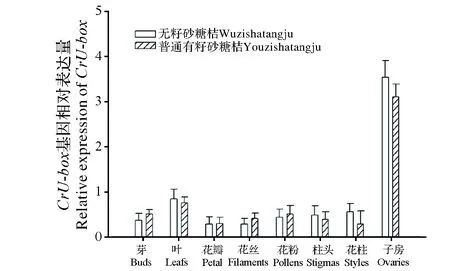
图5 CrU-box基因在无籽沙糖桔和普通有籽沙糖桔不同组织间的表达量Fig.5 Expression analyses of CrU-box gene in different tissues from Wuzishatangju and Putongshatangju by real-time PCR
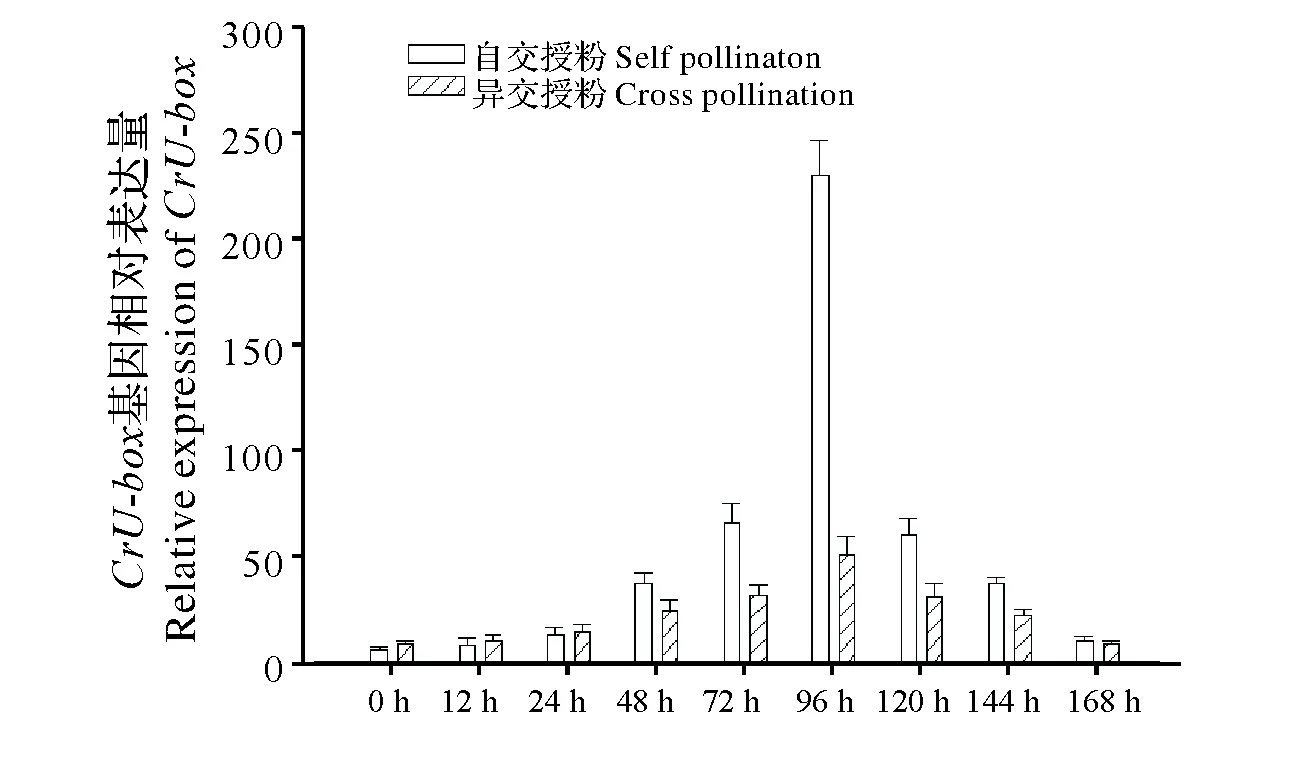
自交授粉为无籽沙糖桔×无籽沙糖桔;异交授粉为无籽沙糖桔×有籽沙糖桔Self pollination(Wuzishatangju×Wuzishatangju);Cross pollination(Wuzishatangju×Putongyouzishatangju)图6 CrU-box基因在自交授粉和异交授粉后不同时间段的表达量 Fig.6 Expression analys of the CrU-box gene in different stages after self-pollination and cross-pollination by real-time PCR
3 讨论与结论
柑橘是我国重要的经济作物,无籽性是柑橘新品种选育的重要目标,而自交不亲和是柑橘无籽果实形成的主要因素。无籽沙糖桔是从广东10 月橘的芽变材料中选育出的新品种,其属于配子体自交不亲和类型[9]。研究表明,配子体自交不亲和反应主要由花粉F-box和花柱中的S-RNase等基因控制[1-2],其它如SKP1-like 蛋白[13]、SSK1互作蛋白[14]、Cullin 1[15]等非S因子也参与自交不亲和反应。泛素/26S蛋白酶体途径作为最重要的蛋白降解途径,在植物的自交不亲和反应中发挥着重要作用[3]。泛素激活酶E1、泛素结合酶E2、泛素连接酶E3等多种酶共同作用促进自交不亲和反应中一些蛋白如S-RNase的泛素化并将其降解,从而抑制花粉管的生长与延伸。作为E3泛素连接酶重要成分的U-box蛋白,在细胞发育、信号转导和光周期反应等反面具有重要生理功能[16]。
为探明无籽沙糖桔自交不亲和的分子机制,本研究以无籽沙糖桔和普通有籽沙糖桔为试材,克隆了无籽沙糖桔E3泛素连接酶CrU-box基因,并在无籽沙糖桔和普通有籽沙糖桔的ORF区域发现4个核苷酸的差异,并导致2个位点的氨基酸发生改变。基因表达分析显示,CrU-box基因在子房中表达量最高,其授粉后0~96 h表达量逐渐增加,96 h表达量达到最大值,96~168 h表达量逐渐下降。但在授粉后48,72,96 h时间段上,自交授粉的CrU-box基因表达量远高于异交授粉后的表达量。该结果与前人发现的无籽沙糖桔自交不亲和发生的抑制部位在子房,自交授粉72 h花粉管开始向远离胚珠的子房底部生长,从而不能完成授粉的结果相一致[17]。在拟南芥中包含U-box/ARM结构域的ARC1蛋白,是SRK激酶的下游信号因子。其自花授粉时,SRK会被SCR/SP11磷酸化,并引起ARC1蛋白磷酸化,并通过U-box结构域与E2泛素结合酶相互作用,使得底物泛素化,进而导致自交不亲和反应的发生[7]。本研究发现的无籽沙糖桔自交授粉后的48~96 h时期CrU-box基因表达量远高于异交授粉,因此笔者推测高特异表达的CrU-box蛋白可能与其它泛素分子结合,导致S-RNase等特异蛋白的泛素化及降解,从而引起花粉管延长受阻或向远离胚珠的子房底部生长,导致无法完成受精过程并形成自交不亲和现象。但CrU-box蛋白作为非S因子,如何参与自交不亲和性反应过程仍需进一步研究。
参考文献:
[1] Sassa H,Kakui H,Miyamoto M,et al.S locus F-box brothers:multiple and pollen-specific F-box genes with S haplotype specific polymorphisms in apple and Japanese pear[J].Genetics,2007,175(4):1869-1881.
[2] Zhang S J,Huang S X,Zhang X L,et al.Identification of S-genotypes of Japanese plum cultivars by PCR and analysis on polymorphism of S-RNase genes[J].Fruit Sci,2008,25(3):338-342.
[3] 陈默,于丽杰,金晓霞,等.植物泛素/26S蛋白酶体途径的研究进展[J].中国生物工程杂志,2014,34(4):118-126.
Chen M,Yu L J,Jin X X,et al.The progress on the ubiquitin/26S proteasome pathway in plants[J].China Biotechnology,2014,34(4):118-126.
[4] Moon J,Parry G,Estelle M.The ubiquitin-proteasome pathway and plant development[J].Plant Cell,2004,16(12):3181-3195.
[5] Sun P,Li S,Lu D,et al.Pollen S-locus F-box proteins of Petunia involved in S-RNase-based self-incompatibility are themselves subject to ubiquitin-mediated degradation[J].Plant Journal,2015,83(2):213-223.
[6] Pickart C M.Mechanisms underlying ubiquitination[J].Annual Reviews of Biochemistry,2001,70(70):503.
[7] Indriolo E,Goring D R.A conserved role for the ARC1 E3 ligase in Brassicaceae self-incompatibility[J].Front Plant Sci,2014,5(5):181-185.
[8] Samuel M A,Mudgil Y,Salt J N,et al.Interactions between the S-domain receptor kinases and AtPUB-ARM E3 ubiquitin ligases suggest a conserved signaling pathway in Arabidopsis[J].Plant Physiology,2008,147(4):2084-2095.
[9] 叶自行,曾泰,许建楷,等.无子沙糖橘(十月橘)的选育[J].果树学报,2006,23(1):149-150.
Ye Z X,Zeng T,Xu J K,et al.Wuzishatangju,a new mandarin cultivar[J].Journal of Fruit Science,2006,23(1):149-150.
[10] Miao H X,Qin Y H,da Silva J A,et al.Identification of differentially expressed genes in pistils from self-incompatibleCitrusreticulataby suppression subtractive hybridization[J].Molecular Biology Reports,2012,40(1):159-169.
[11] 苗红霞.无籽沙糖桔自交不亲和相关基因的分离与鉴定[D].广州:华南农业大学,2012.
Miao H X.Isolation and identification of self-incompatibility related genes fromCitrusreticulataBlanco cv.‘Wuzishatangju’[D].Guangzhou:South China Agricultural University,2012.
[12] Livak K J,Schmittgen T D.Analysis of relative gene expression data using Real-time quantitative PCR and the 2-ΔΔCTmethod[J].Methods,2001,25(4):402-408.
[13] Matsumoto D,Yamane H,Abe K,et al.Identification of a Skp1-like protein interacting with SFB,the pollen S determinant of the gametophytic self-incompatibility inPrunus[J].Plant Physiology,2012,159(3):1252-1262.
[14] Yuan H,Meng D,Gu Z Y,et al.A novel gene,Md SSK1,as a component of the SCF complex rather than Md SBP1 can mediate the ubiquitination of S-RNase in apple[J].J Exp Bot,2014,65(12):3121-3131.
[15] Li W,Chetelat R T.The role of a pollen-expressed Cullin1 protein in gametophytic self-incompatibility in Solanum[J].Genetics,2014,196(2):439-442.
[16] 邵麟惠,郑兴卫,李聪.蒺藜苜蓿E3泛素连接酶U-box基因克隆及表达分析[J].草业学报,2016,25(7):62-72.
Shao L H,Zheng X W,Li C,et al.Cloning and expression analysis of aU-boxgene of E3 ubiquitin ligase fromMedicagotruncatula[J].Acta Prataculturae Sinica,2016,25(7):62-67.
[17] Ye W,Qin Y,Ye Z,et al.Seedless mechanism of a new mandarin cultivar ‘Wuzhishatangju’(CitrusreticulataBlanco)[J].Plant Science,2009,177(1):19-27.
猜你喜欢
中国果业信息(2021年1期)2021-12-01
西北农林科技大学学报(自然科学版)(2021年5期)2021-05-31
银潮(2020年12期)2020-12-22
绿色科技(2020年5期)2020-05-29
草堂(2018年11期)2018-11-13
食品界(2017年7期)2017-08-24
饮食与健康·下旬刊(2016年12期)2017-01-05
阅读与作文(小学低年级版)(2016年10期)2016-10-12
中国果业信息(2015年4期)2015-01-23
中国果业信息(2013年1期)2013-01-23
