
鞣花酸的生物学效应
2019-02-18 10:03吴小磊钟晨史金铭
中国林副特产 2019年1期
吴小磊,钟晨,史金铭
(东北林业大学生命科学学院,哈尔滨150040)
1 引言
1.1 来源
鞣花酸(ellagic acid,EA)发现于1831年,是一种天然酚类,广泛存在于各种软果、坚果等植物组织中,例如核桃[1],草莓[2],葡萄[3],桃子[4]和石榴[5]等。
1.2 化学性质及代谢形式
分子式:C14H6O8
IUPAC命名:2,3,7,8-Tetrahydroxy-chromeno[5,4,3-cde]chromene-5,10-dione
其它命名:4,4′,5,5′,6,6′-Hexahydroxydiphenic acid 2,6,2′,6′-dilactone
结构式:

鞣花酸是没食子酸的二聚衍生物,是一种多酚二内酯,呈反式没食子酸单宁结构。它是一种黄色针状晶体,熔点(吡啶)大于360℃,微溶于水、醇,溶于碱、吡啶,不溶于醚。
鞣花酸通常与葡萄糖,阿拉伯糖,西洛糖等缩合形成糖苷,但更易参与鞣花单宁(Ellagitannin,ET)的形成,植物往往通过水解鞣花单宁产生鞣花酸[6]。在人体中,肠道微量吸收水解鞣花单宁所释放的鞣花酸,大部分鞣花酸进一步被微生物群落代谢为尿石素[7-9]。这些代谢物具有生物可利用性,也具备生物学效应,所以有人认为尿石素才是机体摄入鞣花酸后产生的真正“有益成份”,是鞣花酸能作为抗癌,抗细胞增殖的活性成分的主要原因[10]。
2 生物学功能
鞣花酸具有多种生物活性功能,如抗氧化[11],抑制增殖[12],诱导凋亡[13],阻断病毒感染[14],抑制炎症[15],还可以作为凝血剂[16]等。
2.1 抗氧化作用
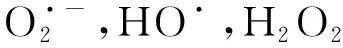
活性氧簇(Reactive oxygen species,ROS)是一类含氧化合物,在体内会损伤核酸,氧化多不饱和脂肪酸和氨基酸,甚至引起炎症反应等。研究表明,EA降低了ROS水平[20-21],进而降低了ROS对机体所引发的伤害。而在野百合碱诱导的小鼠肺动脉高压(PAH)模型中,EA处理导致IL-1β、NLRP3等炎症因子表达降低,同时caspase-1的水平也受到了EA的抑制[22]。综上来看,EA处理后氧化应激(oxidative stress,OS)的减少是改善整个炎症反应的前提条件:首先,NLRP3炎性体参与炎症的多功能细胞因子IL-1β的激活,而ROS的产生是NLRP3激活的中心[23-24];其次,激活的NALP3复合体中的caspase-1(p20)有助于IL-1β的成熟,因此作为上游效应引发者的ROS成为激活caspase-1,IL-1β的关键[24-25]。总而言之,EA通过下调ROS,产生了积极的抗炎症级联效应。
除了调节ROS水平,EA也对非酶糖基化反应中羰基类氧化毒素的产生有抑制效果。蛋白质羰基化是一种由羰基引起的不可逆的氧化反应,被认为是蛋白质氧化损伤和原纤维形成的重要指标[26]。在山梨酸钾和葡萄糖对人血清白蛋白影响的研究中,Taghavi等[27]发现,加速原纤维形成的羰基化合物经EA处理而减少,进而影响了人血清白蛋白淀粉样原纤维的形成,这与其它研究[11,28]结果相一致。具有抗氧化活性的EA终止了氧化反应,进而表现了一种积极的抗纤维化作用。
不仅如此,EA也对抗氧化信号通路产生影响。研究显示,EA能通过上调Nrf2(Nuclear factor (erythroid-derived 2)-like 2)的表达,产生抗氧化效果,保护HaCaT角质形成细胞免受长波紫外线(ultraviolet A ,UVA)引发的氧化应激的伤害[29];同时,EA对Keap1(Kelch-like ECH-associated protein 1)表达也存在抑制作用[29-31]。从以上实验结果来看,Nrf2/Keap1信号通路可能是EA调节机体抗氧化的潜在分子机制。机体中,Keap1可以与Nrf2结合,有助于泛素化蛋白酶降解Nrf2[32];当低水平的ROS或者亲电试剂作用于Keap1时,Nrf2脱离Keap1并转运至核中,与Maf蛋白形成一个二聚体,结合到抗氧化剂反应元件(ARE)或亲电子反应元件(EpRE)上,调节编码抗氧化剂和细胞保护酶的基因的表达[33-39]。在整个过程中,Nrf2和Keap1因EA影响的改变对氧化应激的改善非常重要。
首先根据不同模型处理方法对光谱产生的不同影响[9-11],确定原始光谱的化学计量学模型处理方法。实验先选取6张雷公藤去皮净制完全后的打粉样本光谱,经平均得到1张雷公藤净制完全打粉样本的平均参考光谱,以其为基础,在5800 cm-1~5500 cm-1与4400 cm-1~4200 cm-1的特征谱段处进行验证。实验选取雷公藤未去皮净制的打粉样本2批、去皮净制完全的打粉样本2批、未去皮净制完全的打粉样本2批,合计6批样本,各扫描6张,合计36张NIR光谱,对相应相关系数模型进行验证,求得各6张NIR光谱的平均相关系数),扫描验证结果如下:
此外,EA还能通过上调谷胱甘肽合成酶、谷氨酸-半胱氨酸连接酶的催化亚基(GCLC)和调节亚基(GCLR)来提高谷胱甘肽水平[40],以及诱导NADPH:醌氧化还原酶,血红素氧合酶1[41-44],超氧化物歧化酶,过氧化氢酶,谷胱甘肽过氧化物酶,谷胱甘肽转移酶[21,29,45]等涉及氧化还原反应酶的表达,实现其抗氧化活性。
2.2 放射增敏作用
放射治疗是目前对肿瘤细胞治疗的主要手段之一,而放射敏感性的增加对于辐射剂量的优化和治疗比率的提高有着重要意义。Das等[46]发现EA具有放射增敏效应。EA和γ射线共同处理使与炎症反应有关的IL-6,COX-2,TNF-α的表达降低,并且导致血管生成标志物基质金属蛋白酶(Matrix metallopeptidase 9,MMP-9)水平的下降。同时,活性氧簇(ROS)的产生和积累成为细胞凋亡的诱因,克服天然抗氧化剂的防御作用,在癌细胞中诱导氧化应激并对生物分子造成损伤,其中ROS诱导的DNA损伤促使p53易位至细胞核,随后与细胞周期密切相关的p21表达,细胞周期发生停滞。受EA影响的ROS在放射增敏效应和抗氧化效应中的相反现象可能归因于研究对象和EA处理浓度的不同。此外,EA和γ射线联合处理后,细胞出现的磷脂酰丝氨酸(PS)外化和线粒体膜电位(Mitochondrial membrane potential,MMP)的丧失表明细胞凋亡的启动,而Bax的过表达和caspase-3的活化表明细胞凋亡的发生。EA提高了ROS水平及凋亡相关因子的水平,这与Bhosle等[47-48]的研究发现一致,它们为EA作为放射治疗的辅助手段提供了实验依据,对临床用药有一定的指导意义。
2.3 抗增殖作用和促凋亡作用
化学合成药物可通过抑制增殖、促进凋亡来有效地治疗癌症,但这往往有很强的副作用。近几年的研究发现包括鞣花酸在内的一些天然活性物质[49-51],具有抗增殖、促凋亡的特性。
Hong等[12]发现,EA抑制MCF-7乳腺癌细胞的增殖且使细胞主要在G0/G1期发生停滞,效果呈时间和剂量依赖性。EA处理24h后, cDNA微阵列的分析结果预示了对TGF-β/Smads信号传导途径有调节作用的16个基因(TGF-β1,TβR-Ⅰ,TβR-Ⅱ,Smad3,p-Smad3,Rb1,p-Rb1等)有明显变化,也正因此,TGFβ/Smads信号通路被认为是鞣花酸调控乳腺癌细胞周期阻滞的潜在分子机制。其中,由转化生长因子-β(TGF-β)携带的细胞因子信号将通过I型和II型丝氨酸/苏氨酸激酶受体(TβR-I和TβR-II)的异源性复合物来传递,配体结合导致受体复合物激活,进而导致TβR-II对TβR-Ⅰ的磷酸化。随后磷酸化TGF-β信号的胞内转导物Smads(R-Smads),磷酸化的R-Smads与Smad4相关联,进入细胞核,之后与大量的转录因子相互作用产生相应的生物学功能[52-55]。EA对TGFβ/Smads信号通路中相关因子的调控将直接影响与TGFβ/Smads相关的细胞周期。
除了TGFβ/Smads信号通路,PI3K/Akt(磷脂酰肌醇3-激酶/蛋白激酶B)信号通路也与细胞存活和凋亡之间的平衡紧密相关[56-57],被认为是EA处理结肠癌细胞引发生物学效应的潜在靶点。EA处理使细胞在G2/M期停滞,显著降低活化的PI3K,p-Ak(能灭活前caspase-9,阻断caspase-9参与的凋亡),Bcl-2,DNA聚合酶的辅助因子PCNA,细胞周期蛋白D1的水平,而Bax,cytc及caspase-3表达的增加,显示出EA抗增殖作用和细胞毒性[58]。该信号通路关键蛋白PI3K和Akt的失活促进了与细胞凋亡相关基因(caspase,p53等)的表达,这与许多多酚可调节Akt途径的研究相一致[51,59-60]。目前的研究结果表明,EA在诱导细胞凋亡方面有累加效应[61]。
2.4 抗炎作用
近几年研究发现,大多数植物来源的次级代谢产物能够直接影响炎症介质、第二信使、转录因子等产生来实现抗炎作用[64-65]。Alexandre等[15]在卵清蛋白诱导的小鼠哮喘模型中研究鞣花酸在过敏性肺部炎症中的疗效。结果表明,鞣花酸减少了嗜酸性粒细胞的数量,抑制了细胞募集和细胞因子(IL-4、IL-5、IL-13)的表达并呈时间和剂量依赖性,而且降低了白细胞浸润现象。研究者推测,由于鞣花酸抑制了激活蛋白1(AP-1)[66]和NF-κB[67],所以嗜酸性细胞的招募和细胞因子的产生才得到抑制。
许多粘附分子在炎症过程和免疫应答中也发挥着重要作用[68],EA对炎症的调节作用也在这些方面有所体现[69]在核桃(JuglansregiaL.)中鞣花酸对内皮细胞抗炎效应的研究中发现,在不影响细胞增殖速率的条件下,由炎性细胞因子TNF-α激活的血管细胞粘附分子-1(VCAM-1)和细胞内细胞粘附分子-1(ICAM-1)在实验中受到鞣花酸的抑制,这种效应可能影响某些疾病的发生。细胞粘附分子表达和活化的增加刺激了白细胞募集到血管内皮中,这是动脉硬化发病机制中的重要步骤[70],因此粘附分子是临床中潜在的治疗靶点。而鞣花酸对粘附分子的抑制现象不仅体现了其抗炎作用,而且为其具有潜在的抗动脉硬化能力提供了实验依据。
不仅如此,EA的抗炎作用在组织实验中也得到了很好的验证。Mohammad等[71]发现,全身施用EA可以降低一氧化氮(Nitric Oxide,NO)(只降低血清中,对爪组织中NO无影响)水平,减少多形核白细胞迁移到炎症部位,最终大鼠炎症爪水肿得到缓解且效果呈剂量依赖性。受到EA抑制的NF-κB引起下路的诱导型一氧化氮合酶(iNOS),促炎细胞因子TNF-α,IL-1β的表达水平下调,同时iNOS的产物NO下降[72]。由于L-精氨酸-NO途径参与EA对醋酸引起内脏疼痛的镇痛作用[73]以及EA可通过下调iNOS来抑制NO的释放[74],所以推测这种抗炎作用可能与L-精氨酸-NO途径受到影响有关。而MDA和血浆中的AST和ALT酶的降低[71]预示EA的抗炎作用可能部分归因于其抗氧化活性。
除此之外,EA还可以减少促炎、促纤维化的细胞因子(IL-1α,IL-6,IL-8,TNF-α和TGF-β)的表达,进而减少酒精诱导的炎症反应发生和纤维的产生[75-76]。
2.5 抗病毒作用
在一些研究中,鞣花酸的抗病毒活性也得以发现[77-78]。Promsong等[14]研究了鞣花酸对HIV-1复制的影响。他们的结果表明,鞣花酸抑制了HIV-1整合酶的表达,进而抑制了HIV-1的感染。而且不论HIV-1感染前后,鞣花酸都表现出抑制作用,可能意味着鞣花酸可以抑制HIV-1早期和晚期复制。鉴于抑制整合酶而呈现出显著的抗HIV-1活性,鞣花酸有望作为新型的抗HIV药物。
在其它病毒中,EA呈现不同机制的抗病毒作用。HBx是一种多功能的细胞信号转导和转录途径调节剂,参与乙型肝炎病毒(HBV)复制过程[79]。Pathak等[80]确定EA抑制病毒复制时HBx诱导的转录激活。所以,EA可以降低HBV感染细胞中HBeAg(一种乙型肝炎病毒蛋白)的分泌[81-82],减少HBeAg在血清中的积累,进而阻断了HBeAg引起的免疫耐受[83]。而在丙型肝炎病毒(HCV)中,非结构蛋白NS3具有解旋酶和NTPase活性,再与辅因子NS4A结合组成主要的病毒蛋白酶NS3/4A,而NS5B是RNA依赖的RNA聚合酶,参与病毒RNA的复制。Reddy等[84]和Ajala等[85]证明EA在体外能抑制NS3/4A蛋白酶活性,最终通过减少NS5B活性和HCV中RNA水平,表现出EA对HCV的抗病毒作用。
3 结束语
鞣花酸是一种天然存在的多酚化合物,与石榴等的保健功能密切相关。鞣花酸的结构功能、理化性质等不断被挖掘,其存在潜在的药理学特性一直是研究的热点。虽然鞣花酸的抗氧化,抗增殖,抗病毒等机制被揭示,但目前无法将其投入到疾病治疗中,还需要进一步临床实验评估鞣花酸及其代谢物的药理学作用。此外,鞣花酸生物利用度低也是阻碍其应用的主要问题之一,尿石素及其衍生物的开发利用或许有助于改善这一状况,以鞣花酸为本体修饰产生的衍生物已被用作生物膜形成的理想抑制剂[86]。鞣花酸的生物学效应的研究对癌症治疗和化学预防具有重大意义。相信随着对鞣花酸研究的深入,鞣花酸将在疾病治疗、美容保健等方面表现出积极作用。
猜你喜欢
昆明医科大学学报(2021年12期)2021-12-30
世界科学技术-中医药现代化(2021年5期)2021-11-05
湖南农业大学学报(自然科学版)(2021年4期)2021-08-14
当代水产(2021年6期)2021-08-13
现代临床医学(2021年4期)2021-07-31
昆明医科大学学报(2021年1期)2021-02-07
中华养生保健(2020年7期)2020-11-16
中成药(2019年12期)2020-01-04
建材发展导向(2019年10期)2019-08-24
时尚北京(2016年12期)2017-01-16
