
UV-B辐射对稻瘟病菌侵染阶段四个致病相关基因表达的影响
2019-04-08 07:46黄兰林梅馨月祖艳群何永美
农业环境科学学报 2019年3期
黄兰林,梅馨月,李 详,李 想,祖艳群,何永美,李 元
(云南农业大学资源与环境学院,昆明 650201)
稻瘟病是我国影响水稻生产最严重的病害之一,是由稻瘟病菌(Magnaporthe grisea)引起的水稻真菌性病害,是制约水稻生产的重要因子,导致水稻减产或品质严重下降[1]。适当剂量的UV-B(波长280~315 nm的紫外光)能有效减少病害,与此同时还能增加水稻苯丙氨酸解氨酶活性和类黄酮含量,在2.5 kJ·m-2和5 kJ·m-2UV-B辐射条件下会显著增加,这将有效抵御UV-B胁迫,减轻UV-B对水稻带来的伤害[2]。虽然在高强度UV-B辐射如4.0 kJ·m-2和8.0 kJ·m-2时,根据水稻品种、温度和降雨频率不同等原因,会对水稻生长和产量有不同的影响,但其影响程度远低于稻瘟病害[3]。
紫外线对真菌有很多影响,如死亡、抑制孢子萌发,生长延迟和突变[4]。UV-B对稻瘟病菌的生长、产孢[5]和稻瘟病菌侵染生长[6]有着延缓的作用,并根据辐射剂量不同而影响程度不同。UV-B对植物病原菌疾病、分生孢子及其萌发物,毒力都存在影响,如小麦条锈病[7]、番茄花叶病[8]、十字花科黑腐病[9]、玉米大斑病,这取决于UV-B处理时间、品种、接种水平和植物年龄,一定剂量的UV-B使分生孢子产孢时间滞后,致病力下降[10]。同时也发现随着UV-B辐射的增强,球孢白僵菌[11]、丝孢菌绿僵菌[12]、蝗绿僵菌萌发时间滞后,毒力降低[13]。
紫外线作为诱变剂主要针对生物大分子DNA[14],使其发生诱变,造成基因突变,这必然会影响基因转录和蛋白质的合成,这或许也会影响病菌的致病力[15]。例如木黴菌和米曲霉黑色素合成基因在低剂量UV-B辐射下表达水平的增加,减少了UV-B辐射带来伤害的同时提高了致病力[16],而十字花科黑腐病菌,在受射线如紫外光、X光照射后,Lon基因突变,细胞会因抑制细胞分裂蛋白累积,无法形成隔壁[17]而无法致病,玉米大斑病菌黑色素合成基因Stscd经UV-B照射45 min后表达量明显上升,1 h时表达量达到最高峰,之后开始下降[18]。这些由UV-B引起的基因表达改变,通过改变各种表型来影响侵染致病力[19]。研究发现,NUV(近紫外光)对病原菌的基因表达也会有重要的影响,水稻褐斑病NUV照射可以抑制产生孢子并使PHR1表达增强[20]。除致病基因外,响应NUV的光解酶基因表达水平被NUV辐射会特异性增强,这些反应由蓝/UVA吸收光感受器介导[21],在米曲霉[22]和粗糙脉孢菌[23]中,也发现了相似的真菌蓝光调节蛋白,由此来应对UV辐射[24]。
目前,国内外对于水稻稻瘟病菌致病基因已有大量报道,光胁迫如紫外和近紫外辐射对各类植物和昆虫病原菌致病基因的影响研究也屡见不鲜。但对于UV-B处理稻瘟病菌,对致病基因表达影响的报道尚少。在我国云南元阳梯田地区,高海拔下增强的紫外辐射条件下,稻瘟病病情指数远低于其他地区,其中在5 kJ·m-2强度下病情指数降到最低[6]。在稻瘟病菌侵染整个过程中,从芽管伸长到入侵宿主都涉及细胞壁的合成和变化,已知几丁质为真菌细胞壁的主要组分,几丁质酶是其中重要的胞壁修饰酶[25],Chitinase参与合成几丁质酶,同时也与菌丝生长及孢子的萌发密切相关[26],参与稻瘟病菌生长发育及形态建成以至侵染致病过程[27]。在稻瘟病菌穿透寄主表皮之前,MPG1基因能感知水稻表皮的疏水信号,这是稻瘟病菌能形成附着胞的必要条件之一[28];感知疏水信号后,MAGB基因诱导产生附着胞[29],形成附着胞后所需的侵入机械压力受cAMP途径调节,而此信号途径依赖由CPKA基因编码的蛋白激酶A的活性[30]。为了探寻其中增强的UV-B辐射能有效减少稻瘟病害的分子机理,本研究通过UV-B处理稻瘟病菌,利用实时荧光定量PCR对4个稻瘟病菌致病相关基因(Chitinase、MGP1、MAGB、CPKA)进行表达分析,以期通过分析其表达水平来解释UV-B能如何影响稻瘟病菌的致病力,为在分子层面探索UV-B影响稻瘟病害提供理论依据。
1 材料与方法
1.1 试验材料与使用仪器
1.1.1 供试材料
本试验供试菌株为田间稻瘟病发病水稻上分离保存所得的YN737。
水稻品种:白脚老粳。
1.1.2 试剂
购自 Invitrogen 的 Trizol Reagent、购自 Roche的Universal cDNA Master反转录试剂盒和Essential DNA Green Master,购自Sigma公司的曲利苯兰。
1.1.3 仪器
本试验采用的仪器:Eppendorf Centrifuge 5424 R、Thermo Nanodrop 2000、BIO-RAD Molecular Imag⁃er、Roche LightCycler 96 Instrument和 OLYMPUS BH-2显微镜等。
1.2 试验方法
1.2.1 菌株及水稻的培养
将水稻稻瘟病菌株YN737在米糠培养基上纯化培养7 d,用8 mm打孔器切取菌饼放置在铺有玻璃纸的PDA平板培养基上,28℃与22℃的光暗循环交替培养5 d至菌丝长满覆盖玻璃纸进行UV-B辐射处理。
挑选饱满健康的白脚老粳水稻种子进行催芽,待种子露白,将水稻播种于10 cm×20 cm×7 cm的方盆中,置于温室中培养。
1.2.2 UV-B辐射处理
将培养皿皿盖打开放置在0.5 h预热稳定后的紫外灯(波长280~320 nm,北京电光源研究所提供)正下方,通过调整紫外灯与培养皿的垂直距离来调整辐射强度,其中灯管用0.13 mm醋酸纤维素膜包被,以去除小于290 nm的短波辐射,用UV-B辐射测定仪(北京师范大学光电仪器厂)测定植株顶端辐射强度。辐射强度为0、2.5 kJ·m-2和5 kJ·m-2,辐射处理时间为10、60、120 min,以及辐射120 min后黑布包被避光放置6、24 h,未做辐射处理即辐射处理时间0 min为CK。
待水稻长至三叶一心期,剪取叶片分别制成长3 cm的叶段,采用滴液法将孢子悬浮液接种于水稻叶段上(黑暗保湿24 h),每片叶滴3滴孢子悬浮液,每滴5µL,接种后2、8、32 h、3 d采样染色,接种同时对水稻进行UV-B辐射处理,强度为0、2.5 kJ·m-2和5 kJ·m-2。
1.2.3 材料收集与染色
UV-B辐射处理后,一部分用灭菌的玻片将玻璃纸上的菌丝刮下放入盛有液氮的研钵迅速冷冻研磨;另一部分用无菌水小心洗下孢子,灭菌纱布滤掉菌丝,将孢子浓度调至1×106个·mL-1,将孢子悬浮液均匀喷洒在疏水表面,26℃黑暗保湿培养2、8 h后液氮速冻刮取表面培养物进行研磨。
接种观察试验为每一处理设置3次重复,每一重复为10个叶段,对叶段采用曲利苯兰-酒精乳酚油染色观察。在曲利苯兰-酒精乳酚油(无水乙醇20 mL,苯酚10 mL,83%乳酸10 mL,水10 mL,曲利苯兰10 mg)中沸水浴10 min;将染色后的叶段置于水合饱和氯醛溶液中脱色过夜;保存于50%甘油中。在OLYMPUS BH-2显微镜下观察叶段,接种后2、8 h和3 d照相记录稻瘟病菌萌发形成芽管、附着胞、侵染菌丝侵入水稻细胞的过程。
1.2.4 RNA的提取
取50~100 mg样品,加入1 mL Trizo(l样本的量不宜超过Trizol体积的10%)充分摇匀后室温静置15 min,4 ℃ 13 000 r·min-1离心10 min;小心吸取上清液至新的无酶管,加入200µL氯仿摇匀,室温静置5 min,4 ℃ 13 000 r·min-1离心15 min;吸取上清液,加入异丙醇200 µL,室温静置2~5 min,4 ℃ 13 000 r·min-1离心15 min;去上清,加入1 mL 70%乙醇,4℃13 000 r·min-1离心 5 min;去上清,小心吸取残余液体,室温静置2~10 min;加50~200 µL无核酸酶水,于-80℃保存。
1.2.5 RNA质量和浓度的检测
用1.2%的琼脂糖凝胶电泳检测RNA的完整性,利用Thermo Nanodrop 2000对RNA进行浓度检测和纯度检测。
1.2.6 cDNA模板的合成
用Universal cDNA Master反转录试剂盒对样品进行反转录,Reaction Buffer(5×conc)4 µL,Enzyme Mix(10×conc)2µL,根据RNA浓度调整模板RNA和水的用量,反转体系终体积为20µL,42、85、65℃分别水浴15、5、15 min,完成反转录。
1.2.7 引物设计
利用Primer Premier和NCBI/Primer-BLAST,分别设计了看家基因Actin和其他目的基因的引物(表1),由擎科昆明公司合成提供。
1.2.8 实时荧光定量PCR
以Actin为内参,按照20µL体系(水3µL、10×conc PCR Primer 2 µL、Master Mix10 µL、DNA Tem⁃late 5µL)进行qRT-PCR扩增,反应程序分别为95℃预变性600 s,三步95℃~60℃~72℃扩增,45个循环,95℃~65℃~97℃退火溶解。
1.3 数据处理与统计分析
数据用Excel 2010和LC 96进行整理作图,差异显著性检验采用SPSS 22.0 Duncan检验法(P<0.05)。
2 结果与分析
2.1 RNA的质量与浓度的检测
经过1.2%凝胶琼脂糖电泳,结果如图1所示,RNA18 S、28 S条带清晰,5 S模糊,说明RNA完整性良好。260、280 nm下测定RNA的吸光度,其中A260/A280值均在1.82~2.04之间,说明RNA有机污染少,降解少,质量较好,满足后续实验要求。4个目的基因与内参基因Actin的溶解曲线平滑,单一峰,无引物二聚体,温度介于85~88℃之间,说明引物具有特异性,扩增结果可靠。

表1 引物序列Table 1 Primer sequence
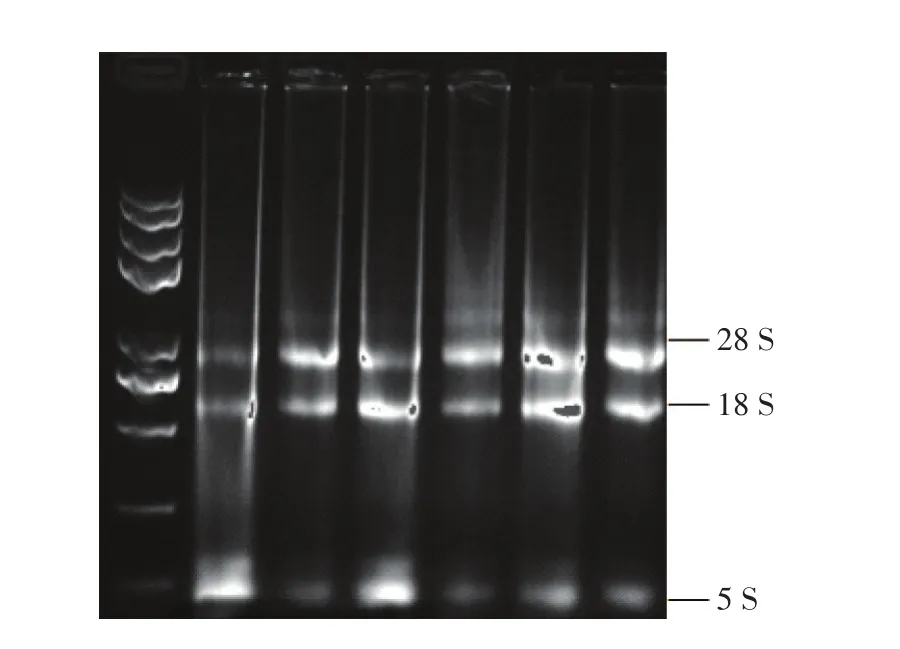
图1 凝胶琼脂糖电泳图Figure 1 Gel agarose electrophoresis
2.2 比较C(tΔΔCt)法分析各目的基因的相对表达变化
从图2 Chitinase的表达变化来看,在2.5 kJ·m-2辐射强度下,Chitinase只在辐射时间为60 min处理下与对照相比表达有显著升高,上调了93%,其余时间均没有显著变化,而在5 kJ·m-2辐射强度下,随着辐射时间的延长,Chitinase表达逐渐降低,在120 min处理下,相对表达量降到最低,从辐射10 min开始到辐射120 min结束,表达较对照分别下调了64%、22%和94%,而在放置6、24 h后,其表达量逐渐回升为对照的48%和54%。

图2 不同UV-B处理Chitinase的表达变化Figure 2 Changes of Chitinase expression under different UV-B treatments

图3 不同UV-B处理MGP1的表达变化Figure 3 Changes of MGP1 expression under different UV-B treatments
对于MGP1,两个辐射强度下(图3),各个处理时间对MGP1表达的影响不大,2.5 kJ·m-2辐射处理时,表达量在各处理时间并没有显著变化,仅在5 kJ·m-2辐射处理时,随着辐射时间的增加,表达量缓慢下调,到120 min处理时,表达量较对照下调了19%,并且在辐射结束后放置6 h和24 h,表达恢复至正常水平。
而MAGB(图4)则是在5 kJ·m-2辐射强度下,各处理时间没有显著变化,在2.5 kJ·m-2强度下,表达会在辐射10~120 min受到显著抑制,与对照相比分别下调了57%、17%和40%,去除辐射放置6 h后,MAGB的表达会回升为对照水平,放置24 h,表达回升且较对照上调了14%。
在两个辐射强度下,CPKA呈相似的表达趋势(图5),在5 kJ·m-2处理时的表达整体低于2.5 kJ·m-2。短时间的辐射使CPKA的表达上升,与对照相比,2.5 kJ·m-2辐射时,10 min与60 min的辐射处理与撤去辐射后放置6、24 h表达上调了1~2倍。5 kJ·m-2辐射时,10 min辐射处理时,CPKA的表达与对照相比上调了49%,辐射60 min和120 min时,表达下调了82%和61%,在撤去辐射后放置6、24 h,表达恢复至对照水平。以上结果表明,UV-B辐射剂量的增加对MGP1的表达没有显著影响,而Chitinase、MAGB、CPKA 3个基因的表达均有所下调,在去除辐射后,表达回升。

图4 不同UV-B处理MAGB的表达变化Figure 4 Changes of MAGB expression under different UV-B treatments

图5 不同UV-B处理CPKA的表达变化Figure 5 Changes of CPKA expression under different UV-B treatments
2.3 UV-B辐射处理下稻瘟病菌侵染阶段生长量的变化
芽管、附着胞和菌丝侵入形成菌落的过程中,5 kJ·m-2处理比2.5 kJ·m-2处理能更大程度地抑制病原菌的侵染(图6)。2.5 kJ·m-2处理下稻瘟病菌各侵染阶段的生长量在芽管形成阶段没有显著差异,在附着胞和菌落形成阶段与对照相比下降了31%和28.6%;而在5 kJ·m-2处理下,3个阶段都有显著下降,分别减少了45.1%、82.2%和75.2%。

图6 UV-B处理下各侵染阶段稻瘟病菌生长量Figure 6 Growth of Magnaporthe grisea in different stages of infection under UV-B treatment
3 讨论
3.1 UV-B辐射处理下致病基因表达水平变化特征
UV-B辐射处理能使菌丝生长变得致密、紧凑,产孢量也随着辐射强度的增加而显著下降[5],在5 kJ·m-2处理时,Chitinase的表达一致。2.5 kJ·m-2辐射处理,在辐射60 min时,Chitinase的表达显著升高,可能是由于低辐射强度的UV-B让病菌对此做出了反应来保护自身,是病菌产生的防护措施[31],如加快细胞的合成与分解来减轻UV-B带来的伤害。黑色素广泛存在于各种真菌体内[16],黑色素基因表达的升高能降低病原菌受到的UV伤害[32],并且发现玉米大斑病黑色素基因在紫外辐射后也表现出相同的趋势,会在辐射处理60 min时,表达量上升至最高峰,随后下降[18],与本试验Chitinase表达趋势相似,辐射下Chi⁃tinase与玉米大斑病黑色素基因表达变化的相似,可能是由于激发了相似的代谢途径来进行自我保护。
而对于MGP1,不同辐射剂量对其表达不产生显著影响,仅在高辐射剂量表达才会有少量下调,推测MGP1对UV-B辐射耐受。细胞已经开发出许多修复或耐受机制来抵消UV或任何其他应激物造成的分子损伤[19],在光解酶的帮助下光还原是各种生物体中最重要且经常发生的修复机制之一[33],此外,切除修复可分别借助多种糖基化酶和聚合酶在几种生物体分子修复中也起重要作用[34]。诱变修复或二聚体旁路、重组修复、细胞周期检查点,细胞凋亡和某些替代性修复途径等机制也存在于各种生物体[22],推测在UV-B处理下可能激发了其中的修复机制,最终免于MGP1表达的改变,但其中可能存在的路径尚不明确。
MAGB则受2.5 kJ·m-2辐射强度的UV-B时有较5 kJ·m-2辐射强度更明显的变化,可能是由于MAGB对低辐射强度较敏感。致病基因的表达除取决于分生孢子的生理状态之外,还取决于辐射剂量[12]。CP⁃KA在两种辐射强度下有着一致趋势的表达变化,在5 kJ·m-2辐射强度处理CPKA的表达量整体低于2.5 kJ·m-2处理,随辐射剂量的增加,基因转录受到破坏,其表达受到抑制。CPKA编码合成致病性所必需的蛋白激酶A的催化亚基单位PKA-c,而cAMP途径依赖于蛋白激酶A的活性,这也可能导致cAMP途径受到影响[35],进一步影响稻瘟病菌的致病力,cAMP信号通路对外界压力胁迫如热激、紫外线辐射、电离辐射等都会做出应答[36]。结合本试验,得知只有高强度辐射处理时能抑制CPKA的表达,撤去辐射,放置6 h和24 h都能发现CPKA的表达恢复对照水平,从白化酵母菌中也能得出,辐射后的光复活和黑暗处理都能产生光还原[37]。
从Chitinase、MAGB、CPKA的表达变化不难发现,在去除UV-B辐射后,降低的表达量会有所回升,所以推测通过逆境筛选留存的稻瘟菌株有适应而产生的修复功能——光还原[37],所以持续或者更长时间的UV-B照射才能使稻瘟病菌降低或丧失致病力。从本文的试验结果可以看到,Chitinase、MAGB、CPKA在UV-B辐射处理下,表达都有显著变化,大致趋势都是在5 kJ·m-2处理强度时表达量相对于对照降低了,变化程度有所不同,已有的研究中就发现光响应基因中2.8%的基因是光敏感的,2%诱导和0.8%抑制[38]。
3.2 UV-B辐射下致病基因表达水平变化与稻瘟病菌侵染力的关系
试验结果印证了UV-B辐射能减轻稻瘟病害以及影响其致病力的假说,其中Chitinase在稻瘟病菌各生长期都有重要作用,说明UV-B对其影响可能贯穿整个生育阶段,低强度辐射时,其表达上升来减轻紫外辐射带来的伤害,随着辐射强度增强和时间的延长,表达下降以致稻瘟病整个生育阶段都有可能受到伤害和抑制,例如产孢量的减少、芽管、附着胞、侵染菌丝形成受阻等等;MAGB与CPKA的表达都影响着cAMP途径[35],该途径与稻瘟病菌的生长发育、孢子萌发、侵染结构形成与致病性都具有重要作用[39-41]。MAGB和CPKA基因的表达变化表示,适当的紫外辐射能让附着胞形成受阻以及侵染机械压力的不足或丧失。从试验结果可以得出,这4个基因在UV-B辐射处理下,其表达都有显著改变,其中5 kJ·m-2处理时,更加显著抑制了表达水平,这与试验结果5 kJ·m-2的UV-B辐射能更大程度减少芽管、附着胞和菌丝侵染,从而更大程度地减少稻瘟病害相一致。
虽然近年对稻瘟病菌致病力相关基因[42]和UV-B辐射对动植物真菌病害影响的研究已较为丰富,但是在分子水平验证UV-B辐射对稻瘟病菌影响的研究尚少,机理尚不清晰,UV-B调节基因的功能尚不清楚,还需进一步探索生物学细节来验证。如调查过度表达,基因破坏或引入反义RNA等实验[23]等,将有助于揭示稻瘟病菌在UV-B作用下分子层面的机理。
4 结论
(1)UV-B辐射能有效减轻水稻稻瘟病的病情指数,减少病害,控制好辐射剂量(如5 kJ·m-2辐射处理)能使病害程度显著降低。
(2)UV-B辐射能有效降低水稻稻瘟病病害的重要机理是UV-B辐射能使稻瘟病菌致病基因的表达发生变化,尤其在5 kJ·m-2辐射处理120 min时,能显著减少致病基因的表达,从而降低病菌入侵对应各个发育阶段的生长量来减少病害。
猜你喜欢
智慧农业导刊(2022年21期)2022-11-03
当代水产(2022年1期)2022-04-26
作物学报(2022年6期)2022-04-08
中国农业科学(2021年23期)2022-01-14
昆虫学报(2021年3期)2021-05-11
中国瓜菜(2020年8期)2020-09-26
农民致富之友(2020年15期)2020-05-25
河北渔业(2020年2期)2020-04-08
中国稻米(2020年1期)2020-03-05
价值工程(2018年25期)2018-09-26
