
北京山区元宝枫夜间液流活动特征及影响因素
2019-06-04 08:05蔡永茂陈立欣陈左司南张志强
生态学报 2019年9期
张 婕,蔡永茂,陈立欣,陈左司南,张志强,*
1 北京林业大学水土保持学院水土保持与荒漠化防治教育部重点实验室,北京 100083 2 北京市八达岭林场,北京 102112
研究表明,测定树干液流时,夜间液流很微弱,但并不为0,夜间液流对蒸腾的贡献也因树种而异[1]。夜间液流对于树木弥补水分亏缺和维持自身水分平衡有重要意义,有助于树体夜间自根系向上运输物质[2],为植物器官的夜间呼吸提供氧传递的机制[3]。尤其在旱季,植物无法从土壤吸收足够的水分,储存水对于蒸腾有着一定的启动作用[4],而夜间液流可以恢复树体水分储备,弥补树体白天根系吸水能力的不足,是储存水的重要来源,从而减轻根系吸水的压力,弥补土壤供水不足。此外夜间液流的存在会使得树木蒸腾耗水和从单木扩展到林分的蒸腾耗水的计算产生偏差,导致对植物正常需水的低估,影响水资源管理和决策。而研究树体夜间液流通量的特征以及影响因子有助于掌握林木的耗水特性和植被应对环境胁迫的调控机制,可以了解林木对周围环境水分的需求和利用效率[5],有助于为植被建设森林健康经营和挑选节水树种提供理论依据。
越来越多的研究表明,夜间液流一部分用于夜间蒸腾,主要由于树木不仅在白天维持着较高的蒸腾耗水,在夜晚也保持着较高的气孔导度和气孔蒸腾;也有研究认为另一部分夜间液流用于补充木质部白天蒸腾引起的木质部水分亏缺,这主要是由于树木存在的水容调节能力,使树体能够在夜间补充白天的蒸腾失水,避免树木体内产生气穴和栓塞[6]。
不同地区不同树种夜间液流动态变化特征以及影响因子有着一定差异。王艳兵等[1]对华北落叶松的研究发现夜间液流晴天变化幅度显著大于雨天,并且不同月份补水贡献率存在明显差异,表现为生长季末期>初期>中期;赵春彦等[7]对胡杨夜间液流通量的研究发现胡杨前半夜夜间累积液流量大于后半夜,且春季夜间累积液流量占整日累积液流量的比例大于夏季和秋季。大量研究表明,气象条件是影响植物液流变化的主要因素,Daley和Phillips[5]认为夜间树干液流与饱和水汽压差呈显著相关;Phillips等[8]发现桉属植物的夜间液流同风速呈显著相关关系;魏潇[9]则发现祁连山青海云杉夜间树干速率与饱和水汽压差、气温呈显著关系,与风速相关性不显著。此外树木形态因子(如树高、胸径、冠幅、边材面积等)也对液流活动产生重要影响[10]。树体是树木储存水的载体[3],储存水的利用又与胸径边材面积等树形因子呈极显著相关[11]。王华等[12]发现夜间液流通量与胸径树高冠幅有很强的回归关系,分别以三次曲线、指数曲线和三次曲线拟合效果最好。
元宝枫(Acertruncatum)是槭树科槭树属落叶乔木,我国吉林省以南至黄河中、下游各省均有分布,具有适应性强、本身耗水量少的特点,属于深根性树种[13-14]。目前, 有关树木夜间液流现象及其对蒸腾量评估的少量研究主要是针对人工林展开的,对于山区的林分夜间液流的差异研究较少,对于元宝枫蒸腾的研究也较少。山区元宝枫的夜间液流通量的调控机制尚未完全弄清,夜间液流通量的用途以及蒸腾耗水的精准预测还有待进一步研究。基于此,本文在北京市八达岭山区对元宝枫林分边材液流进行连续测定,并结合气象因子与土壤水分的连续变化,旨在回答以下问题: (1)元宝枫夜间液流通量的变化规律,(2)元宝枫夜间液流通量的驱动因子以及影响因素,(3)树干夜间液流通量对于蒸腾的贡献率。从而深入揭示树木蒸腾耗水规律和树木储水能力,为植被建设森林健康经营和合理配置水资源、挑选节水树种提供理论依据。
1 材料与方法
1.1 研究地概况
北京地处干旱半干旱区域[15],本实验的研究区位于北京市的北部山区八达岭国家森林公园内(40°17′N,115°55′E),距离北京城区60 km,森林公园总面积为2940 hm2。64%的面积由油松、华北落叶松、元宝枫等针、阔叶林覆盖,区内的乔木林多为20世纪50—70年代所营建,该区域多年平均温度10.8℃,多年降雨量651 mm,无霜期约为160 d,土壤类型为山地褐土,土壤层较薄,腐殖质层厚度为3 cm。
在八达岭森林公园选择生长状况具有代表性的元宝枫样地,样地面积为225 m2,林分密度为2356(株/hm2),样地基本信息见表1。

表1 样地基本情况统计表
1.2 林木边材面积的确定
为避免伤害样树而影响液流的测定,在样地外侧另选14棵不同胸径的树木,量取胸径和树皮厚度,使用生长锥钻取木栓芯量取边材厚度的方法确定边材面积。在树干1.3 m高处钻取树干,将钻得的组织进行染色处理,观察边材和心材分布的区域测量并记录统计边材厚度,以此确定边材面积。利用树木胸径与边材面积的显著相关性建立关系方程,依据这个关系方程计算出每个林分林木的边材面积。
1.3 树干液流通量的测定与计算
在样地内选择6株不同径阶、树木健康、树干通直、适合安装探针的元宝枫作为液流观测样树,采用热扩散法(TDP插针式植物茎流计)对树干液流进行连续观测,样树基本特征见表2。为避免对测定样树的伤害,利用生长锥取木芯,得到边材面积与胸径的关系式,反推出边材面积,以便选取合适长度的TDP探针。之后统一在树干1.3 m高处,用小刀刮去10—15 cm的树皮,根据边材厚度用特定规格的钻头打孔,两孔平行的同时与树干垂直,将10—30 mm的TDP探针插入对应孔,同时避免把探头弄歪和弄断。TDP热扩散液流探针与树干接触处用硅胶密封,在安插探针的部位用铝箔包住树干外部,并用细铁丝扎紧,防止雨水入渗透并减少太阳辐射的影响。数据采集系统外接12 V的电源,保证其持续稳定的供电。探针与CR1000数据采集器相连,以便采集连续数据,数据采集器记录两个探针之间的温差数据,记录数据为30 min的平均值,每10天下载一次数据。
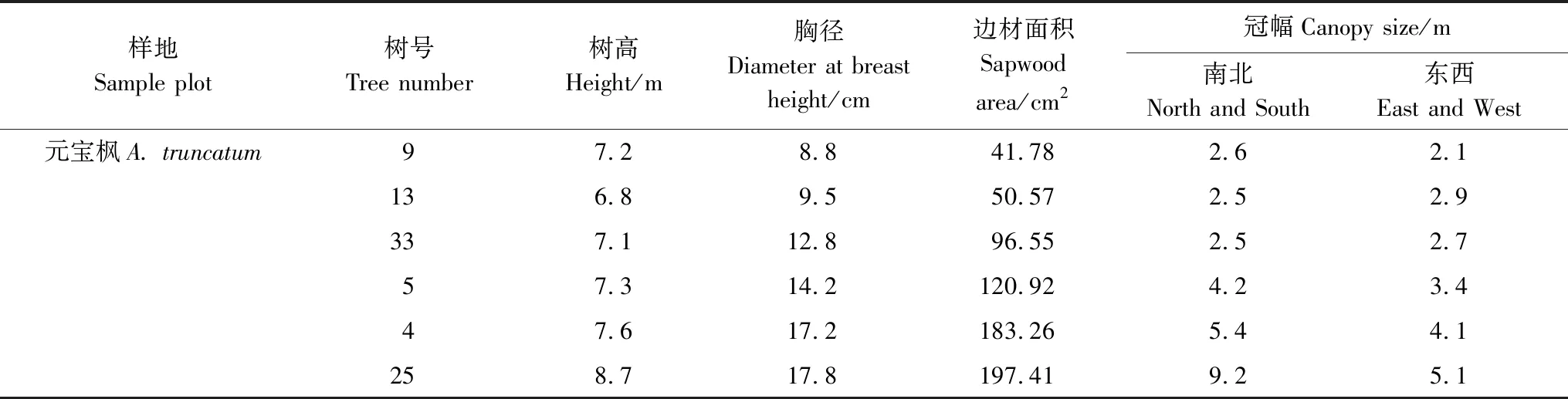
表2 样树基本特征统计表
根据Granier[16]建立的热扩散探针(TDP)上下探针的温度差与树干液流密切相关的原理,利用两者关系的经验公式将温度差转换为树干液流:
(1)
式中,Js为单株样木液流密度(g cm-2s-1),ΔT是加热探针和数据传感探针之间的温度差(℃),ΔTm是每1天中最大的温度差值(℃)。单株样木的日耗水量为:
Fs=Jsi·AS·1800
(2)
式中,FS为单株耗水量(g);AS为样木边材面积(cm2)。由于林分中树木胸径存在分异,故以胸径为尺度扩展因子推求林分尺度的日蒸腾量Ec(mm),林分蒸腾量计算公式为:
(3)
式中,As为林木边材面积(cm2),Aground为研究地面积(m2),Js为单位边材面积上的液流通量(g cm-2s-1)。
1.4 环境因子
选择林内空旷的平地,在样地周边安装自动气象监测站,对太阳辐射强度(Rn, W/m2),空气温度(T,℃),空气相对湿度(RH,%),光合有效辐射(PAR,W/m2),风速(W, m/s)等进行同步监测,饱和水汽压亏缺(VDP, kPa)可以采用大气温度和相对湿度实测值用计算,计算公式如下:
(4)
式中,T表示大气温度,RH为空气相对湿度。
另外,在矩形样地对角线布置土壤水分监测点,埋入土壤水分传感器(5TE)。八达岭林地由于地处山区,0—20 cm和20—40 cm的土层>2 mm 的石砾的体积比例分别为16%和13.2%[17],故测定深度为20 cm、40 cm。并用数据采集器EM50每隔30 min记录一次数据进行连续测定,仪器记录时间设为30 min,每10天下载一次数据。用铁铲挖至埋入土壤水分传感器的深度,取出充满土壤的环刀,将采集的土样带回实验室,测定包括田间持水量,土壤容重在内的土壤理化性质,采用土壤相对有效含水率REW对土壤水分状况进行分级,REW的计算公式如下:
(5)
式中,VWC(%)为土壤体积含水率,VWCmax(%)为研究时段内最大土壤含水率,即研究地的田间持水量(%),VWCmin(%)为研究时段内的最小土壤含水率。
1.5 数据分析
将一天中光合有效辐射为0的时段定义为夜间液流,将TDP温差数据导入Baseliner软件,以此实现原始数据的质量控制和原始数据的转换,从而将TDP温差数据转变为液流数据,之后用Excel以及SPSS进行数据处理。采用单因素方差分析的方法分析夜间液流通量动态变化,采用曲线参数估计法分析树干夜间液流通量与树木形态因子之间的回归关系,采用多元回归分析和相关分析的方法分析树干夜间液流通量与环境因子的关系。
2 结果
2.1 夜间液流速率变化特征
从7月份元宝枫样地液流速率与太阳辐射变化趋势可以看出(图1),树干液流的日变化呈典型的单峰曲线,从6:00开始空气温度不断升高,太阳辐射加强,蒸腾强度不断增大,12:00达到最大值,之后随着太阳辐射减小温度降低蒸腾随之减弱,并在夜间到达较低水平。但当19:00左右光合有效辐射变为0后,仍然可以观测到元宝枫存在明显的夜间液流。根据光合有效辐射的监测数据,将夜间液流计算时间段确定为19:00到次日5:00。

图1 7月份元宝枫液流速率及太阳辐射的变化Fig.1 The daily variations of sap flow rate and solar radiation of A. truncatum in July
观察不同月份的元宝枫树干夜间液流速率变化特征(图2)可以看出,7月份元宝枫夜间液流速率的变化平缓,液流通量没有明显的上升和下降点,基本保持在0.00002 g cm-2s-1左右。其余月份夜间液流通量在前半夜较后半夜明显活跃,夜间液流从19: 00开始呈下降的趋势,其中19:30到00:00夜间液流下降较为明显,之后趋于平缓,但仍可观测到明显的夜间液流,说明夜间液流的变化主要存在于前半夜。由于白天蒸腾比较强烈导致树木水分亏缺,日落后根系与土壤之间的水势差仍会存在[18],所以刚入夜夜间液流较高以弥补白天水分亏缺,之后树木水分亏缺程度减弱,夜间液流随之减小。
此外,5、6月份夜间液流液流速率明显高于7、8、9月份,且前半夜夜间液流速率变化的幅度也比7、8、9月份大,表现在液流速率起始点较大,下降的较为明显。生长季初期枝叶伸展,生长活动旺盛,可以说明此时树体利用一定夜间液流来应对生长季初期自身的水分需求。
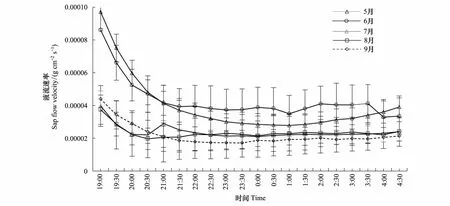
图2 元宝枫各月夜间液流速率变化Fig.2 The daily variations of nighttime sap flow rate of A. truncatum
2.2 夜间液流通量变化特征
可以看出夜间液流速率的变化规律表现为19:00—0:00逐渐下降,之后趋于平缓(图2),以0:00为界区分前半夜和后半夜,同时观察元宝枫夜间累积液流通量的月际变化(表3)。可以看出,夜间液流前半夜和后半夜所占的比例不同的月份之间也有一定差异(表3)。5月的夜间累积液流量最大,此时树木生长活动旺盛,新叶萌生,白天蒸腾量较大,6月份次之,最小的为生长季末期的9月份。同样可以看出,前半夜夜间液流高于后半夜,前半夜夜间累积液流量占夜间累积液流量的53.85%—64.10%,而后半夜占夜间累积液流量的35.9%—46.15%,平均夜间液流通量为5月>6月>8月>9月>7月。
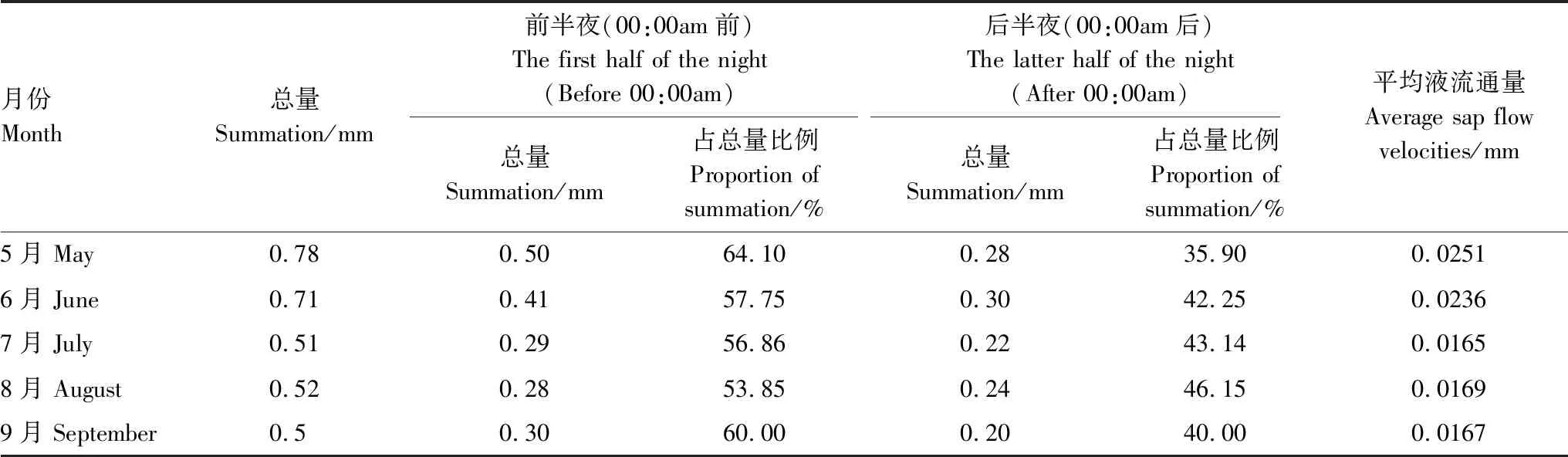
表3 元宝枫各月夜间累积液流通量变化
2.3 夜间液流通量的影响因子
2.3.1树木形态
采用单因素方差分析分析了处于相同的生长环境和气候条件下的6株元宝枫的夜间液流通量变化,方差同质性检验的结果为P<0.001,表明形态结构不同的树干夜间液流通量有显著差异。因此采用曲线参数估计的方法分别对树干夜间液流通量与胸径、树高和冠幅进行回归分析,通过曲线参数估计法,可以得出11种拟合模型,最后挑选出R2最高的模型。通过分析可知,树干夜间液流通量的与胸径树高冠幅之间的回归关系分别以三次曲线模型(R2=0.734,P=0.031),二次曲线模型 (R2=0.889,P=0.018),和三次曲线模型(R2=0.536,P=0.035)的拟合关系最好。
观察6株样木夜间液流通量与树木形态因子的变化关系(图3),可以看出夜间液流通量与胸径树高冠幅呈正比的关系。在一定范围内,胸径树高冠幅越大的样木,夜间液流通量越大,可以说明树木的形态因子也是影响树木夜间液流的重要因子。

表4 不同样木元宝枫夜间液流通量方差分析
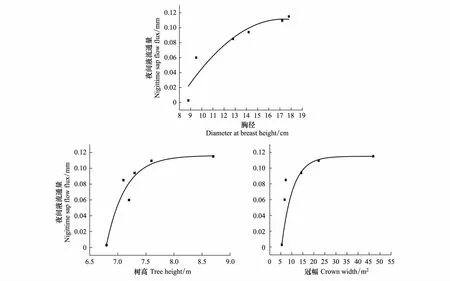
图3 元宝枫夜间液流通量与树形因子的变化Fig.3 Variations of nighttime sap flow flux at night and shape of tree factor of A. truncatum
2.3.2降雨以及土壤水分
观察降雨数据以及土壤水含量各月变化发现7月份降雨较多,表5的数据可以反映出,7月份土壤水分条件较好,土壤体积含水率较高,土壤相对有效含水率为0.58且大于0.4,此时元宝枫不存在干旱胁迫的情况,可见土壤水分不是夜间液流的限制因子。土壤体积含水率为7月>8月>9月>6月>5月,土壤相对有效含水率7月份最大,生长季初期较小,最小为5月份的0.12,此时生长季初期的元宝枫存在明显的干旱胁迫现象。通过对比各月土壤含水率变化,5、6月土壤水分环境较差,加之生长旺盛连续的蒸腾失水,树干水分亏缺严重。在生长季初期,一旦有所降雨,土壤水分条件改善,树体组织大量吸收水分会使得树体水势增大[19],从图中可以看出降雨当天以及之后的几天夜间液流总量会出现增长(图4),表现为生长季初期较为明显,可以看出此时夜间液流有一部分用于补水。
通过观察林地的降雨量与夜间液流通量的变化以及夜间液流通量与土壤含水量的变化关系(图4、图5),观察到5、6月份存在干旱胁迫的情况下,土壤含水量增大时夜间液流有不同程度的增多,可以发现降水对树干夜间液流的影响是通过影响土壤含水量实现的,土壤水分环境差时降雨后的几天夜间液流会增大,以弥补白天蒸腾造成的水分亏缺,维持树体自身的水量平衡。
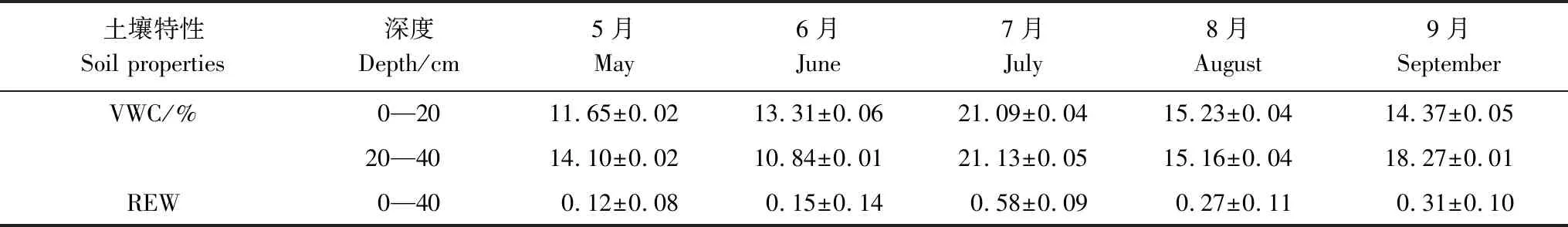
表5 样地各月土壤体积含水率及土壤相对有效含水率变化
VWC:土壤体积含水率 Soil moisture(%);REW:土壤相对有效含水率 Relative extractable soil water
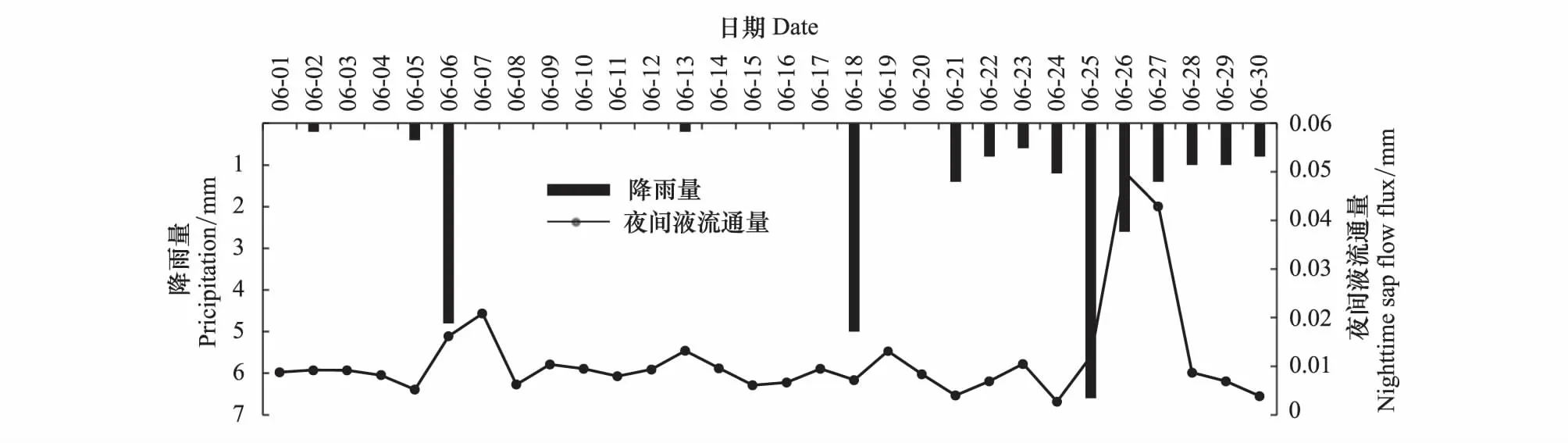
图4 元宝枫降雨量与夜间液流通量的变化Fig.4 Variations of pricipitation and nighttime sap flow flux of A. truncatum at night in June
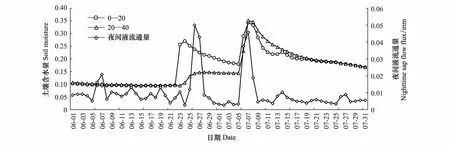
图5 元宝枫夜间液流通量与土壤含水量变化Fig.5 Variations of nighttime sap flow flux and soil moisture of A. truncatum
2.3.3环境因子
从图6中可以看出,夜间液流通量与饱和水汽压差,风速存在一定的相关关系,而饱和水汽压差又是温度和湿度共同作用的,可见温湿度也影响着夜间液流通量的变化。元宝枫夜间液流通量会随着叶片边界层水汽压与叶片气孔内腔水汽梯度变小,液流通量也减小,随气温的升高夜间液流通量不断升高,气温影响了气孔的开放,而随着空气湿度增大,元宝枫夜间液流呈减小趋势,此时风起到的作用是打开气孔以促进夜间蒸腾,环境因子一定程度上可以解释元宝枫夜间液流的变化,说明元宝枫在生长季存在少量的夜间蒸腾。
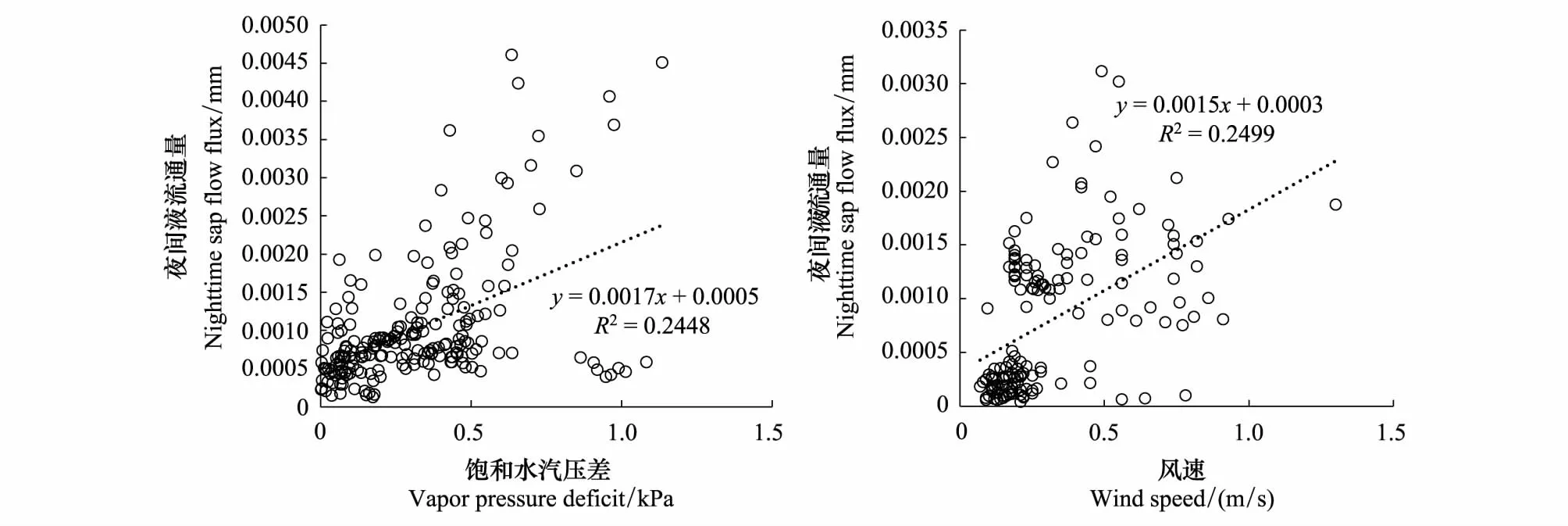
图6 元宝枫夜间液流通量与环境因子的相关关系 Fig.6 Correlation of nighttime sap flow flux at night and environmental factor of A. truncatum
将夜间液流总量与日间液流总量进行拟合(图7),发现夜间液流总量与日间液流总量呈显著相关,元宝枫二者的相系数为0.466(R2=0.217,P=0.00,n=154)。从植物生理的角度来看,日间蒸腾越强烈,植物出现自身水分亏缺的程度就越大,白天较强烈的补水需求会带动较为活跃的夜间液流活动,造成木质部含水量的增加和木质部的扩张[20]。

图7 元宝枫夜间液流通量与日间液流总量的相关关系 Fig.7 Correlation of nighttime sap flow flux and sum of diurnal sap flow of A. truncatum
将夜间液流通量与测定的各个环境因子以及土壤含水量和整日蒸腾进行相关分析,结果表明不同月份夜间液流通量的影响因子之间的相关关系略有不同(表6)。总体上看各月夜间液流通量与土壤含水量和整日蒸腾的相关性较为显著,其中7月份夜间液流通量与土壤含水量呈显著相关(R2=0.708),且是在P<0.01水平下显著。各月夜间液流均与日蒸腾相关性较高,可以看出7、8月份夜间液流通量与饱和水汽差、空气相对湿度、空气温度以及风速相关性较为微弱,此时的夜间液流更多的用于补水,而补水量取决于日蒸腾强度造成的树体水分亏缺程度以及土壤含水率决定的所需的补水能力,受环境因子的影响不大,可以认为夜间蒸腾很微弱。再结合7、8月份土壤水分条件较好的情况(表4),可以解释7、8月份夜间液流通量较低的现象。
观察生长季初期和末期,夜间液流通量同饱和水汽压差、空气温度以及风速呈现出一定的相关关系,但也不能完全解释夜间液流的变化,夜间液流通量与整日蒸腾量和土壤含水量相关系数也较大,生长季初期存在干旱胁迫的条件下白天蒸腾强烈,树体水分亏缺,对夜间液流的需求量大,风速和空气温湿度等环境因子改变气孔的张度和蒸腾机制,结合与日蒸腾量相关性较强的现象,认为此时夜间液流一部分用于补水的同时,还有一部分夜间液流用于夜间蒸腾。
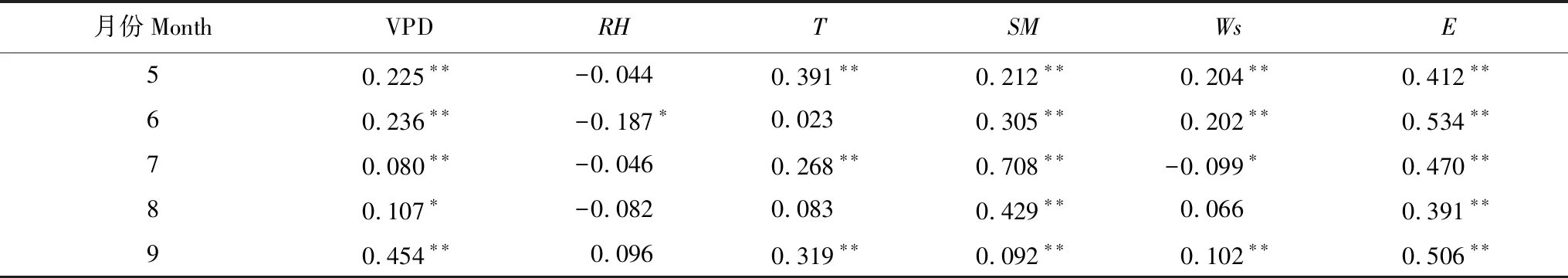
表6 元宝枫各月夜间液流通量与环境因子的Pearson相关系数
*为0.05水平上差异显著,**为0.01水平上差异显著;VPD:饱和水汽压差 Vapor pressure deficit(kPa);RH:空气相对湿度 Relative humidity(%);T:空气温度 Air temperature(℃);SM:Soil moisture(%);Ws:风速 Wind speed(m/s);E:整日蒸腾 Sum of diurnal sap flow(mm)
生长季初期和末期夜间液流一部分用于补水一部分用于蒸腾,而在土壤水热条件好的生长季中期元宝枫夜间液流更多的是用于补水。以0:00为界区分前半夜和后半夜,分别对夜间液流通量及可能的环境影响因子进行相关分析(表7)。可以发现,元宝枫呈现出一定的规律,总体上看夜间液流通量与饱和水汽压差、空气温度、空气相对湿度、土壤含水量、风速相关,其中前半夜夜间液流通量相较后半夜相关性较为显著,可以认为夜间蒸腾存在于前半夜。生长季初期存在干旱胁迫的条件下,夜间液流一部分用于夜间蒸腾一部分用于补水,夜间蒸腾多发生在前半夜,后半夜以补水为主;而在土壤水分较为充足时,元宝枫前半夜后半夜的夜间液流主要的作用都是补水。对于后半夜的夜间补水,表现为较为平缓的夜间液流,也可能用于皮孔蒸腾和角质蒸腾。可以发现元宝枫不同的生长环境不同月份的夜间液流的影响因子和用途有一定差异。
表7 前后半夜元宝枫夜间液流通量与各环境因子的相关性分析
Table 7 Correlation analysis of nighttime sap flow flux and environmental factor ofA.truncatumat the first half of the night and the latter half of the night

时段 TimeVPDRHTWsSM前半夜The first half of the night0.279∗∗-0.293∗∗ 0.175∗∗ 0.089∗∗0.145∗∗后半夜The latter half of the night0.078∗-0.0270.104∗0.0370.193∗∗
2.4 夜间液流占整日蒸腾的比例
对不同月份元宝枫夜间液流贡献率的方差分析结果为P=0.229>0.05(表8),表明各月夜间累积液流量占整日蒸腾的贡献率差异不显著,但可以观察到不同月份贡献率仍有变化。

表8 不同月份元宝枫夜间液流贡献率方差分析
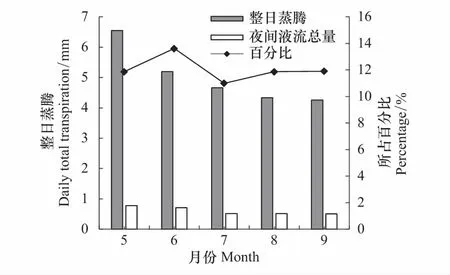
图8 元宝枫不同月份夜间液流通量占整日蒸腾的比例 Fig.8 The proportion of nighttime sap flow to daily total transpiration of A. truncatum

图9 土壤含水量与夜间液流贡献率变化 Fig.9 Variations of soil moisture and the contribution rate of nighttime sap flow to transpiration
从图8可以看出元宝枫各月夜间累积液流量占整日累积液流量的比例为6月>9月>5月>7月>8月,其中6月最高达14%,平均贡献率为12%。生长季初期夜间累积液流量占整日累积液流量的比例大,是因为5、6月相比7、8月份平均温度低,平均光合有效辐射低,使树木整日的蒸腾量较低,且夜间液流很大程度弥补白天水分亏缺,夜间补水量大,导致夜间累积液流占的比例较大,此时树干夜间补充的水分对树体应对干旱胁迫起到更大的作用。而夏季蒸腾强烈,白天液流通量比晚上大的多,导致7、8月份比例较低,白天蒸腾量越大,夜间树干水分补充对于整树的贡献率就小。
可以发现夜间蒸腾补水贡献率与土壤含水量变化有一致性(图9),在生长季初期,降雨过后引起的土壤含水量增大,此时夜间液流增大,夜间液流贡献率也随之增大,这种情况表现在存在干旱胁迫的条件下。而在水热条件较好的7月份,夜间液流并没有很高,夜间液流贡献率也不高,可见元宝枫在生长季初期气温较低水热条件差的情况下,夜间补水量对于树木维持自身正常生长以及保持自身水量平衡应对春季干旱胁迫起到了作用,而在水热条件较好的情况下占的比例是较少的。
将夜间液流通量对总蒸腾的贡献率与各环境因子进行相关分析(表9),可以看出元宝枫夜间液流通量的日蒸腾贡献率与白天蒸腾量以及累积太阳辐射有相关关系,且成负相关。白天较强的太阳辐射使得蒸腾明显增多,树干水分亏缺严重进而造成贡献率变小。这些环境因子对贡献率的作用从大到小依次为太阳辐射>白天蒸腾量>风速>空气湿度。
3 讨论
3.1 夜间液流通量的变化特征
本文发现夜间液流速率前半夜较后半夜更为活跃,总是在刚入夜后呈下降趋势,之后趋于平缓。这是由于白天蒸腾比较强烈导致树木水分亏缺,日落后根系与土壤之间的水势差仍会存在[18],所以刚入夜夜间液流较高以弥补白天水分亏缺,之后树木水分亏缺程度减弱,夜间液流随之减小。

表9 夜间液流通量贡献率与影响因子之间的相关分析
Rn:太阳辐射强度 Rolar radiation intensity(W/m2)
此外,5、6月份前半夜液流速率的下降幅度比7、8、9月份显著,这种现象与生长季枝叶伸展,生长活动旺盛有关。元宝枫夜间液流通量不同的月份之间有一定差异,表现为5月的夜间累积液流量最大,6月份次之,最小的为生长季末期的9月份。生长季初期树木生长活动旺盛,新叶萌生,白天的蒸腾量较大,导致树体产生较大水分亏缺,则需要较高的夜间液流以弥补白天的水分亏缺。对比生长季初期较高的夜间液流,生长季中期夜间液流通量并不高,这与当时的环境有很大关系,在土壤水分较好的生长季中期,虽然植物蒸腾需求大,但雨天光合有效辐射减弱,蒸腾作用降低,树干可以在白天充分吸收水分,导致树体水势下降不大,水分亏缺程度有所减弱,加之元宝枫本身植物组织中束缚水比例高,细胞原生质粘滞性以及原生质胶体的亲水性强,有利于树体保持自身水分[21],当土壤条件较好时,自身水分亏缺并不严重。而在9月份温度降低,树木生理活动减弱,树体水分亏缺较轻,结合土壤含水量相比生长季中期下降,土壤水分环境变差的情况,夜间液流通量会有所下降。
3.2 夜间液流的影响因素
夜间液流被认为是树干贮水量的重要组成部分,这意味着树木本身的贮水量可能是影响夜间液流的主要因素[22],由于树干贮水量是树木本身的特征,影响着树体水容的大小,同时储存水的利用又与胸径边材面积等树木形态因子呈极显著相关[11],因此夜间液流也会受到树形因子如胸径树高冠幅边材面积的影响。
Phillips等[23]研究发现每日的水分利用会随着直径变化大幅度增加,表现为直径0.34 m的榆科树种的日蒸腾为46 kg,而直径0.98 m的日蒸腾为750 kg,树形因子影响整日蒸腾进而影响夜间液流通量的变化,Goldstein等[11]也发现树干基部整日的液流量与边材面积和树高显著正相关。本研究中可以看出夜间液流通量随着胸径树高冠幅的增加不断增大,说明树形因子通过影响边材面积和整日蒸腾来影响夜间液流通量的变化,进而影响树木储存有效水[24],表现为越高大的的样木,夜间液流通量越大。
针对环境因子对夜间液流通量的影响,尹立河等[25]对陕西榆林地区旱柳和小叶杨夜间液流研究发现前半夜树干液流密度主要受饱和水汽压差的影响,后半夜则主要受风速的影响。本文发现除不同树木形态的元宝枫夜间液流通量有显著差异的同时,不同月份夜间液流通量的影响因子也略有不同。生长季的中期即7、8月份夜间液流通量与饱和水汽压差,空气相对湿度,温度以及风速相关性较为微弱,这与Fisher等发现的夜间树干液流与饱和水汽压差显著相关[26]的结果不同,这可能由于研究地的环境条件差异以及树种的不同导致。7、8月份降雨较多,土壤水分条件较好,夜间液流通量会更多的用于补水,补水量取决于土壤含水量和日蒸腾强度。5、6月份夜间液流通量同饱和水汽压差、温度以及风速呈现出一定的相关关系,夜间液流速率与夜间天气条件的相关性支持了气孔开放的假设[27],此时存在一定夜间蒸腾,驱动力为饱和水汽压差、温度、空气相对湿度、风速等环境因素的变化。但也不能完全解释夜间液流的变化,夜间液流通量与整日蒸腾量和土壤含水量相关系数也较大,认为此时较大的夜间液流活动是夜间蒸腾和补水作用共同造成的。且Donovan等[28]认为夜间液流的出现缘于冠层蒸腾导致的水分缺失以及夜间根系的吸水,可见用于补水的夜间液流通量驱动力源于日蒸腾强度造成的树体水分亏缺程度以及土壤含水率决定的所需的补水能力,夜间液流通量与土壤含水量呈现线性相关[29],土壤含水量低时夜间液流通量也比较低[26]。
本文发现生长季初期夜间液流通量既然用于夜间蒸腾又用于补水,生长季中期则主要用于补水。用于夜间蒸腾的夜间液流通量与饱和水汽压差、温度、空气相对湿度、风速相关,随着水汽压差的增加,叶片边界层水汽压与叶片气孔内腔水汽梯度增大,蒸腾增强,夜间液流通量呈上升趋势[30],而用于补水的夜间液流通量与土壤含水量和日蒸腾强度相关性较强,这与王华发现的较为干旱的情况下,环境因子对夜间液流通量的影响明显,雨季一定程度上会掩盖环境因子对夜间液流通量的影响[12]较为一致。其中前半夜夜间液流通量相较后半夜与环境因子的相关性较显著,可以认为夜间蒸腾主要存在于前半夜,而后半夜以补水为主,在气孔处于接近关闭状态下的液流活动更可能用于体内水分亏缺的补充[31]。
3.3 夜间液流占整日蒸腾的比例及用途
本文发现夜间累积液流通量占整日蒸腾的比例6月最高达14%,5个月的月平均贡献率为12%,生长季初期夜间累积液流量占整日累积液流量的比例大。沈振西等[18]发现半干旱半湿润地区的树种夜间液流通量占整日蒸腾的比例高,认为气候越干旱植物夜间液流的比例大,这也是植物适应水分限制环境的一种表现。
本研究表明元宝枫生长季初期和末期夜间补水量对日蒸腾的贡献率比生长季中期高,主要是生长季初期树木整日的蒸腾量较低,且夜间液流很大程度弥补白天水分亏缺,夜间补水量大,造成贡献率较高,这与王艳兵等人对华北落叶松夜间树干液流的研究结果一致。同时赵春彦等[7]也发现胡杨夜间累积液流量对整日的贡献率春季大于夏季和秋季,周翠鸣、赵平等对广州地区荷木夜间液流研究发现干季夜间补充的水分对整树蒸腾的作用更大[4]。王华等[12]发现平均气温低,水热条件差的月份,夜间水分补充量对于蒸腾的贡献率高,此时期内树干夜间补充的水分对植物体起更大的作用。夜间液流通量的日蒸腾贡献率生长季初期大于生长季中期,可见夜间累积液流通量对整日蒸腾的贡献率不同月份有一定差异,且干季比湿季贡献率更高。
而对于树干液流的用途,一部分人认为是树干夜间补水造成的,也有人认为是夜间蒸腾造成的。王华认为夜间液流密度前半夜和后半夜相比变化幅度大,水分补充主要时期在前半夜,树干水分补充足够以后观察到的液流是用于皮孔蒸腾和成熟叶片的角质层蒸腾,并且通过分析夜间液流与水汽压亏缺和风速的关系确定是否存在夜间蒸腾。本文发现元宝枫夜间液流通量同样用于蒸腾和补水,生长季初期存在干旱胁迫的条件下,夜间液流一部分用于夜间蒸腾又有一部分用来补水,生长季中期则主要用于补水,夜间树干液流与环境因子相关性不高,并且夜间蒸腾出现在前半夜,后半夜以补水为主。对于夜间液流的驱动机制,不同月份表现出不同特征,生长季初期水热条件差时,用于夜间蒸腾的夜间液流通量驱动力为饱和水汽压差、温度、空气相对湿度、风速等环境因素。Dawson等发现处于土壤含水量较高和不容易出现水分亏缺的环境中的植物,会表现出较高的夜间液流通量值[32],而蒸发需求又是夜间液流较强的驱动力[33],可见用于补水的夜间液流通量则取决于日蒸腾强度造成的树体水分亏缺程度以及土壤含水率决定的所需的补水能力。生长季中期时夜间液流通量主要是用于补水,驱动因子同样是土壤含水量和日蒸腾强度。
元宝枫各月夜间累积液流量对整日累积液流量的平均贡献率为12%,尽管夜间液流占全天蒸腾的比例较低,但是在进行大尺度估算时,也应考虑这一比例,如果估算植物日间蒸腾量时忽略这一比例,容易导致林分耗水的低估。
4 结论
(1)树干液流速率日变化为典型单峰曲线,元宝枫生长季存在明显夜间液流。且夜间液流前半夜较后半夜明显活跃,前半夜呈下降的趋势,后半夜趋于平缓,前半夜夜间累积液流量占夜间累积液流量的53.85%—64.10%,而后半夜占夜间累积液流量的35.90%—46.15%。
(2)5、6月份前半夜液流速率的下降幅度比7、8、9月份显著,5月的夜间累积液流量最大,平均夜间液流通量为5月>6月>8月>9月>7月。
(3)存在水分胁迫的条件下降雨之后和降雨之后的几天夜间液流会增大,土壤水分条件较好,土壤水分不再是夜间液流的限制因子时,夜间液流通量并不高。
(4)生长季初期和末期,用于夜间蒸腾的夜间液流通量同饱和水汽压差、空气温度以及风速呈现出一定的相关关系,用于补水的夜间液流通量取决于日蒸腾强度和土壤含水率。生长季中期夜间液流通量与饱和水汽压差,空气相对湿度,空气温度以及风速相关性较为微弱,补水量取决于日蒸腾强度造成的树体水分亏缺程度以及土壤含水率决定的所需的补水能力。
(5) 生长季初期夜间液流一部分用于补水一部分用于蒸腾,且夜间蒸腾存在于前半夜,后半夜以补水为主。而在土壤水分条件好的生长季中期元宝枫夜间液流更多的是用于补水。补水需求带动夜间液流活动,弥补白天水分亏缺,并作为储存水的重要来源。
(6)元宝枫生长季初期和末期夜间补水量对日蒸腾的贡献率比生长季中期高,平均贡献率为12%。
关于树干夜间液流通量仍有许多方面需要探索,例如元宝枫夜间液流通量对于总蒸腾量的估算造成多大的误差,其次存在干旱胁迫和土壤水分条件较好的两种情况下,夜间液流通量的差异到底有多大,如果夜间蒸腾和补水同时存在时,哪种起主导作用等问题,在这点上,国外有研究通过监测邻接冠层的树干位置的液流变化判断夜间水分补充是否用于夜间蒸腾,也有观测胸高处夜间液流密度值变化,判断水分补充还是夜间蒸腾,这些问题都有待进一步的研究。同时也可以对其夜间气孔导度和径向生长的变化进行监测,结合元宝枫植物生理特征方面的研究,深入了解元宝枫夜间蒸腾的机制[34]。
猜你喜欢
中国林业产业(2021年5期)2021-12-14
小猕猴学习画刊(2021年10期)2021-04-18
山东林业科技(2019年2期)2019-06-03
小雪花·成长指南(2019年12期)2019-02-10
吉林农业(2019年9期)2019-01-06
时尚北京(2018年1期)2018-01-22
都市丽人(2017年6期)2017-07-25
农村农业农民·B版(2016年12期)2017-01-11
中国质量与标准导报(2014年5期)2014-02-28
储能科学与技术(2014年5期)2014-02-27
