
烟蓟马趋光规律及不同波长色板田间诱捕效果
2019-06-11 00:32米娜张起恺王海鸿吴圣勇雷仲仁
中国农业科学 2019年10期
米娜,张起恺,王海鸿,吴圣勇,雷仲仁
烟蓟马趋光规律及不同波长色板田间诱捕效果
米娜,张起恺,王海鸿,吴圣勇,雷仲仁
(中国农业科学院植物保护研究所植物病虫害生物学国家重点实验室/农业农村部农产品安全生物性危害因子(植物源)控制重点实验室, 北京 100193)
【目的】在室内、温室和露地3种条件下,通过测定烟蓟马()对不同单色光及不同波长诱虫色板的行为反应,探究对烟蓟马具有最佳趋性的波长、光照强度,以及相关影响因子如性别、日节律、饥饿、温度、相对湿度对烟蓟马趋光行为的影响,为改进诱虫色板及诱虫灯等烟蓟马绿色防控技术和产品提供依据。【方法】首先,室内使用分辨率高、光谱测量范围广、稳定性好的单色仪测试装置测定烟蓟马对10种单色光的行为学趋性,筛选出烟蓟马趋性较强的单色光,并将初始光照强度通过中性密度滤光片衰减,探究光照强度对烟蓟马趋光行为的影响。其次,以烟蓟马趋光率为统计指标,探究性别、日节律、饥饿、温度、相对湿度对烟蓟马趋光行为的影响。根据室内试验结果,利用Dan Bruton虚拟波长及RGB值的函数关系,打印出相应波长的蓝色和黄色诱虫色板,涂胶制板。评价温室和露地条件下,烟蓟马对不同波长自制色板及不同厂家生产色板的趋性。【结果】室内试验结果显示,烟蓟马对450 nm的蓝色光趋性最强,趋光率高达75.34%,其次为562 nm的黄色光以及430 nm的蓝紫色光,趋光率分别为73.61%和64.03%。在430、450、562 nm单色光刺激下,烟蓟马雌虫的趋光性均强于雄虫。光强衰减试验结果表明,随着光照强度增加,烟蓟马趋性增强。在上午8:30—10:00时,烟蓟马对430、450、562 nm单色光最为敏感。饥饿4 h后,烟蓟马的趋光性最强,之后随着饥饿时间的延长,趋光性降低。温度为25—30℃时,烟蓟马对3种单色光趋性极显著高于对照组;15℃时,烟蓟马对单色光的刺激均不敏感。相对湿度为45%—60%时,烟蓟马对430、450、562 nm单色光的趋性均显著强于对照组,而在相对湿度为30%和90%时,烟蓟马对单色光的趋性与对照组均无显著差异。通过在温室和露地中应用不同波长色板及不同厂家生产的诱虫色板对烟蓟马的诱集效果比较,结果发现450 nm的蓝色自制诱虫色板和反射波长为440—470 nm的2#诱虫色板均对烟蓟马诱虫效果最佳,此试验结果与室内筛选的最佳波长高度吻合。【结论】性别、日节律、饥饿、温度和相对湿度对烟蓟马的趋光行为均有一定的影响。室内光谱光强和诱虫色板试验均表明烟蓟马对450 nm蓝色光以及波长在450 nm左右的蓝色诱虫色板有明显的趋性,450 nm左右的蓝色诱虫色板可用于监测和防治烟蓟马。
烟蓟马;趋光性;单色光;诱虫色板
0 引言
【研究意义】烟蓟马()又被称为葱蓟马、棉蓟马、瓜蓟马以及韭菜蓟马,主要通过吸取植物汁液和传播病毒危害植物,在世界各地广泛分布[1-2]。其寄主范围较广,据统计可造成上百种作物不同程度的减产[3]。近年来,随着国际贸易的频繁,烟蓟马逐渐成为世界性害虫[4],可造成超过50%的农业减产[5-6]。化学防治是现阶段防治烟蓟马的主要方式,但长期使用农药使烟蓟马已对多种杀虫剂产生抗药性,并且对环境产生一定影响[7-10]。目前,烟蓟马已对阿维菌素、多杀菌素、乙基多杀菌素、甲维盐和螺虫乙酯等新型选择性杀虫剂产生抗药性,抗性最高超过了21 000倍[11]。因此,探索绿色环保的烟蓟马防治技术迫在眉睫。利用昆虫趋光性这一特性来防治烟蓟马具有无农药残留、成本低及对环境友好等优点,前景广阔。【前人研究进展】昆虫普遍具有趋光性,趋光行为是昆虫的一种主要行为方式[12]。目前,国内外有关烟蓟马趋光行为的研究大多是通过诱虫灯、诱虫色板等方法来完成的。例如,利用灯光诱捕以及温室或露地悬挂黄蓝诱虫板,这些技术已广泛应用于蔬菜、果树、烟草等作物上监测烟蓟马的种群动态及防治[13-16]。但是有关蓟马对不同波长及颜色的行为趋性等研究结果并不一致。Hoddlea等[17]研究发现,白色诱虫色板对西花蓟马()和纹蓟马()的诱集效果最佳,鳄梨蓟马()对黄色的偏好性强于白色,3种蓟马均对蓝色色板无明显的趋性;吴青君等[18]研究发现,波长在438.2—506.6 nm的海蓝色对西花蓟马的诱集效果最好;Rodriguez-Saona等[19]测试了硬蓟马()对白色、黄色、蓝色3种颜色的趋性,结果显示硬蓟马对白色诱虫色板的趋性强于蓝色和黄色色板;Demirel等[20]研究发现,荧光黄色诱虫色板对烟蓟马有明显的吸引力,且蓝色和白色诱虫色板对蓟马的吸引力无显著差异;Gharekhani等[21]利用诱虫色板在不同作物上诱集烟蓟马的结果显示,烟蓟马对黄色和蓝色具有很强的趋性,且两者之间无差异。【本研究切入点】国内外市场上现有的诱虫灯及诱虫色板的波谱范围宽泛、颜色标准和量值不统一,这不仅严重影响试验结果,也弱化了其应用效果,使得通过诱虫色板或诱虫灯防治烟蓟马的技术受到了阻碍,因此亟需筛选烟蓟马最强趋性的波长和光强,并进一步探究不同因素对烟蓟马趋光行为的影响。【拟解决的关键问题】通过对烟蓟马进行不同单色光刺激,筛选出烟蓟马的最佳敏感光谱;通过光照强度的改变,总结烟蓟马随之变化的规律;明确影响烟蓟马趋光行为的因素;进一步运用Photoshop铜板纸打印特定波长诱虫色板,依据温室和露地试验探究烟蓟马趋性,明确趋性色板波长及相应的参数值,并与不同厂家生产的诱虫板进行比较,为开发和研制防治烟蓟马的诱虫灯和诱虫色板提供依据。
1 材料与方法
室内试验于2017年在中国农业科学院植物保护研究所完成,田间试验于2018年5—9月在中国农业科学院廊坊科研中试基地完成。
1.1 供试昆虫及材料
烟蓟马采自中国农业科学院廊坊科研中试基地的葱()上,在光照培养箱(MLR-351H,SANYO Electric Co.Ltd)中用菜豆()饲养,饲养温度为(25±1)℃,相对湿度60%—70%,光周期14L﹕10D。
不同厂家诱虫板包括佳多诱虫板:蓝色、黄色、绿色、白色、红色、粉色、黑色、灰色、青色、紫色;宝鸡广仁诱虫色板:黄绿色和姜黄色;中捷四方诱虫色板:蓝色和黄色。面积均为20 cm×25 cm,诱虫色板颜色确定依据《中国颜色命名》。
自制诱虫色板:应用Dan Bruton虚拟波长与RGB值的函数关系(http://www.efg2.com/Lab/Science And Engineering/Spectra. htm),将不同的虚拟波长转换为相应的RGB值(表1),利用Photoshop软件并采用高光铜版纸打印出相应的色板,色板尺寸为30 cm×20 cm。将粘虫胶(佳多科工贸有限责任公司)均匀地涂在色板两面。
1.2 试虫处理
室内趋光行为学试验选取5日龄烟蓟马成虫,置于行为反应箱的活动室中,为了使烟蓟马的复眼适应状况保持一致,在每次光刺激前,将烟蓟马放置暗室中适应2 h。
1.3 光谱测量
光谱仪选用美国ASD(Analytical Spectral Device)公司的ASD FieldSpec HandHeld野外便携式光谱仪(波段值325—1 050 nm)。测量时间选在11:00—14:00无风晴朗的午间,测定需先进行光谱仪优化、暗电流采集、BaSO4白板矫正和设定重复测定值20。光谱仪的探头垂直向下距诱虫色板约3 cm,诱虫色板的中心反射波长为反射率的峰值。
1.4 试验装置与行为反应方法
行为反应光路:高压氙灯光源HGILX150和单光栅单色仪HGIMS3012购买于北京衡工有限公司。氙灯为150 W德国进口灯泡,单色仪分辨率为0.1 nm,光谱范围是330—1 000 nm。中性密度滤光片购买于上海恒选仪器科技。

表1 11种诱虫色板及其对应的RGB值和虚拟波长
根据烟蓟马的行为特性并参考相关文献[22],设计趋光行为装置,并制作烟蓟马行为反应盒。烟蓟马行为反应盒为U型,主要分为3个部分,分别为避光室(30 cm×6 cm×7 cm)、趋光室(30 cm×6 cm×7 cm)和反应活动室(16 cm×5 cm×7 cm)。为了便于观察和计数,反应盒的上部为透明亚克力有机玻璃板,其他各部分均为黑色不透明的亚克力有机玻璃板。试验开始时,将试虫从反应活动室的小孔中放进去。接着将趋光室、避光室以及反应活动室上的开孔用配套的盖子和黑布盖上,以防试虫逃脱。单色光经过聚光镜后汇聚于趋光室,置于反应活动室的试虫可同时观察到趋光室和避光室的光,从而做出选择。
1.5 室内光谱行为反应测定
趋光行为反应:试验在黑暗无光的环境下进行,室内温度(27±1)℃,相对湿度(60±5)%。单色光的波长分别为340、380、430、450、483、498、562、582、605、620 nm,每次光刺激15 min。每次试验选取健壮的试虫45头左右为一组,重复3次。为了减小误差,每次试验采用的试虫均不重复,各波长刺激后,取出反应盒,快速统计趋光室和避光室内试虫的个数,计算其趋光率和避光率。选取光谱试验中具有较高趋性的3种单色光,测定不同性别烟蓟马对这3种单色光趋光性的差异。趋光率=趋光室内虫数/总虫数×100%。
光强行为反应:选取上述光谱行为反应试验中具有较高趋性的3种单色光,分别为430、450、562 nm。光强度用不同的中性滤光片调节,单色光的光强度范围以0.01、0.03、0.10、0.32、1.00倍数渐次增强,试验方法同趋光行为反应。
烟蓟马日光谱行为反应:设8:30—10:00、11:30 —13:00、14:00—15:30、17:00—18:30 4个处理,方法同趋光行为反应。
不同饥饿时间下烟蓟马的光谱行为反应:设饱食、饥饿4、8、12、16、24 h 6个处理,方法同趋光行为反应。
不同温度下烟蓟马的光谱行为反应:在15、20、25、30、35℃ 5种温度下,分别测定烟蓟马对430、450、562 nm 3种自制诱虫色板的趋性。试验使用Y形透明玻璃管,两臂及基部长10 cm,两臂之间的夹角为45°,直径为2 cm。试验前,先将烟蓟马和Y管置于光照培养箱内(MLR-351H,SANYO Electric Co. Ltd)15 min,以便烟蓟马能够先适应试验温度。试验时,将自制色板卷成直径为3 cm的圆筒套于Y管的一臂,另一臂以培养箱内的白光为对照。将烟蓟马放置于离Y管基部5 cm后,用实验棉塞住两臂和基部的开口,5 min后统计两臂内烟蓟马的数量。每次试验选取烟蓟马5头,重复8次。
不同相对湿度下烟蓟马的光谱行为反应:在30%、45%、60%、75%、90% 5种相对湿度下,分别测定蓟马对430、450、562 nm 3种自制诱虫色板的趋性。方法同不同温度下烟蓟马的光谱行为反应。
1.6 诱虫色板的温室和露地试验评价
1.6.1 10种诱虫色板对温室烟蓟马诱集效果 试验在烟蓟马危害严重的大葱日光温室蔬菜棚内开展。试验地区域种植的大葱要求在整个种植过程中不使用任何化学农药。在选择的试验地设置上述10种诱虫色板进行诱集试验,按照随机区组均匀排列。每种颜色重复3次且同种颜色位于不同的排,共计3排,每排10种颜色。第二天早晨8:00之前调查每块板上蓟马的数量,连续观察3 d。
1.6.2 不同波长自制诱虫色板的温室和露地试验评价 温室试验:选择对烟蓟马有较强趋性的蓝黄两种颜色,根据国际照明委员会(CIE)对颜色与波长之间对应关系的划分标准,将蓝色波段和黄色波段进一步划分,划分为虚拟波长分别为430、440、450、460、470、480、490 nm的蓝色色板和560、570、580、590 nm的黄色色板。将其打印成对应的色板,涂胶制板,悬挂于温室中。调查方法同1.6.1,每个处理重复4次。
露地试验:露地试验所用波长分别为430、440、450、460、470、480、490、560、570、580、590 nm。自制诱虫色板在试验田中分4排17列悬挂,避免同排或同列诱虫色板相同,交替悬挂诱虫色板。每排间距4 m,每列间距3.3 m。诱虫色板悬挂于高于地面60 cm处。调查方法同1.6.1。
1.6.3 不同生产厂家诱虫色板的温室和露地试验 温室试验:选取实验室现有的4种黄色诱虫色板和3种蓝色诱虫色板进行烟蓟马诱集。3种蓝色诱虫色板分别编号为1#、2#、3#,其中1#蓝色诱虫色板购买于北京中捷四方有限公司,2#和3#蓝色诱虫色板购买于河南佳多有限责任公司。4种黄色诱虫色板分别编号为4#、5#、6#、7#,其中4#和7#诱虫色板购买于宝鸡广仁生物科技有限公司,5#购买于河南佳多有限责任公司,6#购买于北京中捷四方有限公司。调查方法同1.6.1,每个处理重复4次。
露地试验:将上述3个不同厂家的4种不同黄色诱虫色板以及2个厂家3种蓝色诱虫色板分别设为一个处理,共设 7个处理,重复4次,即每种颜色使用4张,共计28张。调查方法同1.6.1。
1.7 数据处理与分析
数据采用SPSS13.0软件进行处理,用ANOVA方法进行方差分析,若有显著性,采用Duncan法比较显著性差异(<0.05)。在进行方差分析前,对每组数据进行方差齐性、正态分布检验和处理,以确定该组数据满足方差分析的需求。对只有两个处理数据的差异性分析采用独立样本T检验。试验数据采用Origin 8.5软件作图。
2 结果
2.1 室内光谱行为反应测定
2.1.1 趋光行为反应 在340—620 nm单色光的刺激下,烟蓟马均能产生一定的趋光行为反应。由图1可以看出,烟蓟马趋光反应曲线呈双峰型,主峰为450 nm的蓝色光,趋光率高达75.34%,次峰为562 nm的黄色光,趋光率为73.61%。烟蓟马对这两种单色光的趋性显著高于340、380、483、498、582、605、620 nm的单色光。烟蓟马对605和620 nm的红色光趋性较弱,其趋光率显著低于其他8种单色光。
由图2可知,无论是雌虫还是雄虫,在 430 nm 蓝紫色光、450 nm 蓝色光和 562 nm 黄色光的刺激下,雌虫的趋光率均高于雄虫趋光率。烟蓟马雌虫对450 nm的蓝色单色光趋性最强,趋光率高达78.12%。在430 nm蓝紫色光和450 nm蓝色光刺激下,雌虫的趋光率均显著高于雄虫的趋光率。而在562 nm黄色光的刺激下,烟蓟马雌雄成虫趋光率无显著差异。
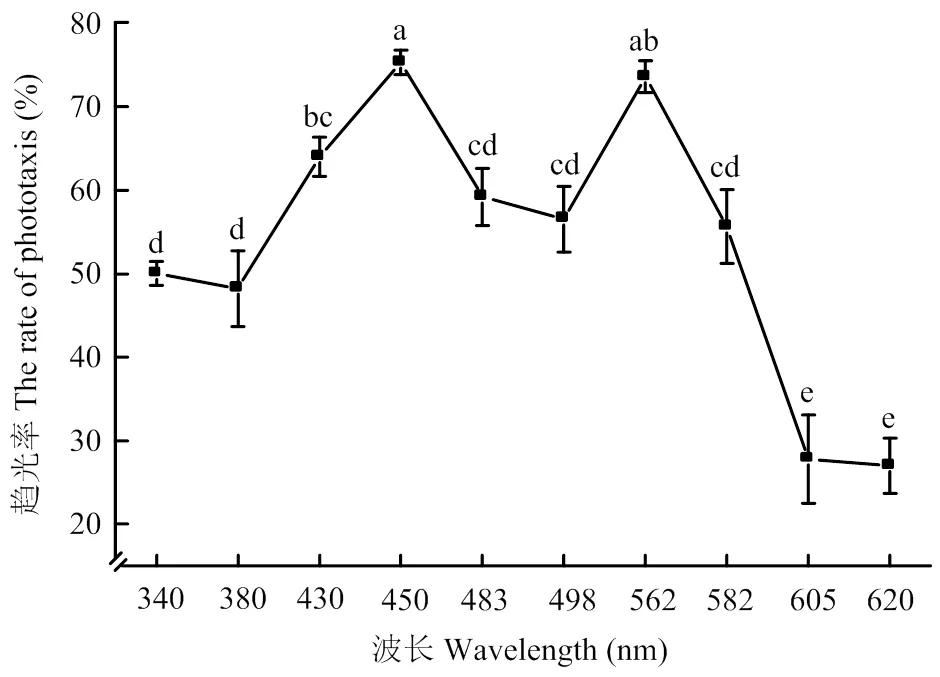
数值为平均值±标准误 Data are mean±SE;不同小写字母表示处理间显著差异(P<0.05)Different lowercase letters indicate significant differences among treatments (P<0.05)。下同The same as below
2.1.2 光强行为反应 随着光照强度的增加,烟蓟马对上述3种高趋性单色光的趋性均表现为增强。在430 nm蓝紫色光和450 nm蓝色光下,烟蓟马的趋光行为反应曲线相似,即随着光照强度的增强,烟蓟马的趋光行为反应也随之增强。在光强衰减至初始光强的0.01—0.03倍时,趋光率增加缓慢,后期均有大幅度的增加。而在562 nm的光刺激下,光强衰减至初始光强的0.01—0.03倍时,其趋光行为增加幅度最大,后期增加相对较为缓慢(图3)。

图2 不同性别烟蓟马的趋光率

图3 烟蓟马在不同光照强度下的趋光率
2.1.3 日节律行为反应 烟蓟马对同种单色光在不同时间段内的趋性差异显著。在1 d内,烟蓟马在8:30—10:00和14:00—15:30内对光较为敏感,活动能力比较强。在8:30—10:00对562 nm黄色单色光趋性最强,趋性反应率高达79.19%,显著高于其他3个时间段。在8:30—10:00时,烟蓟马对430 nm蓝紫色光和450 nm蓝色光的趋光反应率显著高于11:30—13:00和17:00—18:30时,与14:00—15:30无显著差异(图4)。

图4 烟蓟马日节律反应
2.1.4 不同饥饿时间下烟蓟马的光谱行为反应 测定饱食、饥饿4、8、12、16、24 h后烟蓟马对3种单色光的趋性结果表明,经4 h饥饿处理后,烟蓟马对3种单色光的趋光性增强。随着饥饿时间的延长,除450 nm的蓝色光外,烟蓟马的趋光率均下降。其中,烟蓟马对562 nm的黄色光趋光性最强,趋光率高达80.03%,显著高于饱食、饥饿8、12、16、24 h时的趋光率。经12 h饥饿处理后,烟蓟马对450 nm蓝色光的趋性增强(图5)。
2.1.5 不同温度下烟蓟马的光谱行为反应 在430 nm单色光刺激下,烟蓟马对不同温度的趋光率为30℃>25℃>35℃>20℃>15℃。在450 nm单色光刺激下,烟蓟马对不同温度的趋光率为25℃>30℃>35℃>20℃>15℃。在562 nm单色光刺激下,烟蓟马对不同温度的趋光率为30℃>25℃>20℃>35℃>15℃。烟蓟马在温度为25℃时对450 nm的趋性最强,趋光率高达87%,极显著高于对照组。在温度为35℃时,烟蓟马对450 nm和562 nm单色光的趋光率均显著高于对照组。在温度为25℃和30℃时,烟蓟马对这3种单色光的趋光率极显著高于对照组(图6)。
2.1.6 不同相对湿度下烟蓟马的光谱行为反应 由图7可知,在430 nm的单色光刺激下,相对湿度为45%和60%时,烟蓟马对此单色光的趋性极显著高于对照组。在相对湿度为75%时,烟蓟马对单色光的趋性显著高于对照组。在相对湿度为30%和90%时,烟蓟马对单色光的趋性与对照组无差异。在450 nm的单色光刺激下,烟蓟马在相对湿度为60%时趋光反应率最高,趋光率高达76.46%,相对湿度75%次之,两者均极显著高于对照。在相对湿度为30%和90%时,烟蓟马对单色光的趋性与对照组无显著差异。在562 nm单色光刺激下,相对湿度为45%、60%和75%时,烟蓟马对单色光的趋性极显著高于对照组,在相对湿度为30%和90%时,其趋性与对照间无显著差异。
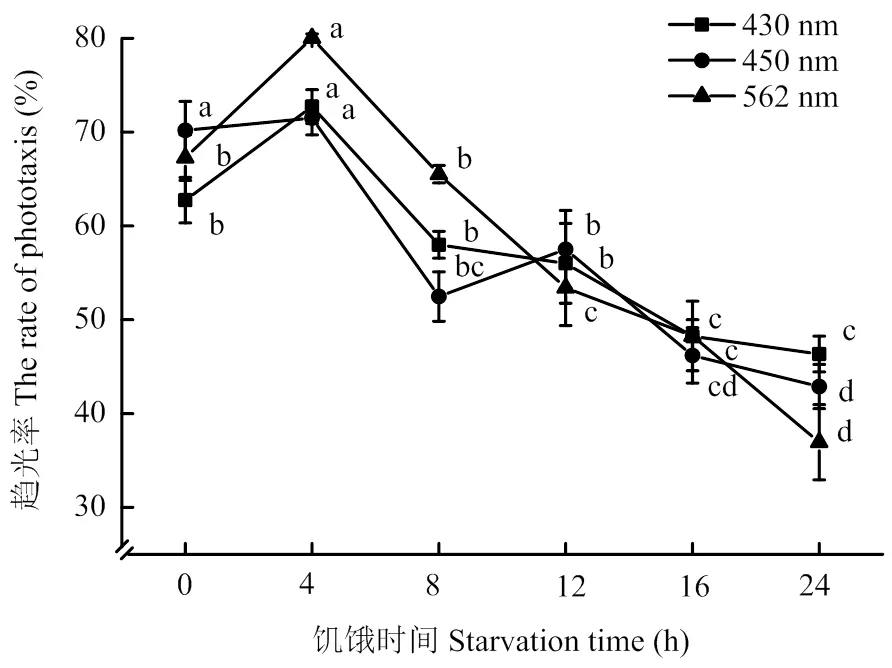
图5 不同饥饿时间下烟蓟马的趋光率
2.2 诱虫色板的温室试验评价
2.2.1 10种诱虫色板对温室烟蓟马诱集效果 温室试验表明,10种颜色的诱虫色板对烟蓟马的诱集效果不同,诱集数量从小到大依次为红色(波长630—780 nm)<粉色(波长630—780 nm)<黑色(无)<绿色(波长500—550 nm)<白色(波长390—780 nm)<紫色(波长375—400 nm)<黄色(波长560—600 nm)<灰色(波长400—415 nm)<青色(波长475—500 nm)<蓝色(波长440—493 nm)。蓝色诱虫色板诱集对烟蓟马的吸引力最强,平均诱集量达到(234±4)头/板,其次为青色,平均诱集量(87±11)头/板(图8)。红色、粉色以及黑色诱虫色板对烟蓟马无吸引力,平均诱集量不足10头。说明烟蓟马对蓝色更加具有趋性。经野外高光谱辐射仪ASD FieldSpec Pro FR2500测定,该板的波长为440—493 nm,中心波长为466 nm。

* P<0.05;** P<0.01;ns:差异不显著no significant difference。图7同 The same as Fig. 7

图7 不同相对湿度下烟蓟马的趋光率

图8 10种诱虫色板的平均诱集量
2.2.2 不同波长自制诱虫色板的温室和露地试验评价 根据室内烟蓟马光谱行为学试验以及温室10种诱虫色板诱集试验结果表明,烟蓟马对蓝色和黄色的诱虫色板趋性较强。为了进一步明确蓝色和黄色光谱中对烟蓟马有最佳趋性的颜色的虚拟波长,将蓝色和黄色光波段划分为若干个波长,各虚拟波长所对应的RGB值以及相对应的颜色如表1所示。
温室试验结果表明,烟蓟马对430—460 nm的蓝色诱虫色板的趋性显著高于560—590 nm的黄色诱虫色板。其中,烟蓟马对虚拟波长为450 nm蓝色诱虫色板的趋性最强,平均诱集量为(288±33)头/板。其次是波长为430 nm的蓝色诱虫色板,平均诱集量为(264±28)头/板,以及460 nm的蓝色诱虫色板,平均诱集量为(258±22)头/板,这3种蓝色诱虫色板间无显著差异。470—490 nm 3种诱虫色板的诱集效果相对较差,平均诱集量都不足百头,显著低于波长为430—460 nm的4种诱虫色板。不同虚拟波长的黄色诱虫色板对烟蓟马的诱集无显著差异。在黄色波段内,570 nm的黄色诱虫色板对烟蓟马的诱集效果最好,平均诱集量为(70±4)头/板(图9)。
露地试验结果表明,不同波长的诱虫色板对烟蓟马的诱集效果不同,430—470 nm的蓝色诱虫色板的诱集量显著高于480—490 nm蓝色色板和560—590 nm的黄色色板。在蓝色波段中,450 nm的诱虫色板的诱集效果显著高于430、470、480、490 nm的诱虫色板,与440和450 nm的蓝色诱虫色板无显著差异。在黄色波段中,不同波长的黄色诱虫色板无显著差异,570 nm的黄色诱虫色板诱集效果最好(图9)。
温室和露地试验结果均表明,450 nm的蓝色自制诱虫色板对烟蓟马的诱集效果最佳。
2.2.3 不同生产厂家诱虫色板的温室和露地试验评价 温室结果表明,不同生产厂家的诱虫色板对烟蓟马的诱集效果呈现显著差异。2#蓝色色板对烟蓟马的诱集效果显著高于1#和3#蓝色色板,最高平均诱集量达到(257.56±29.98)头/板,且显著高于其他6种诱虫色板。3种蓝色色板均诱集到上百头烟蓟马。在黄色诱虫色板中,5#黄色诱虫色板的诱集量最高,显著高于7#诱虫色板,与4#和6#诱虫色板无显著差异(表2)。
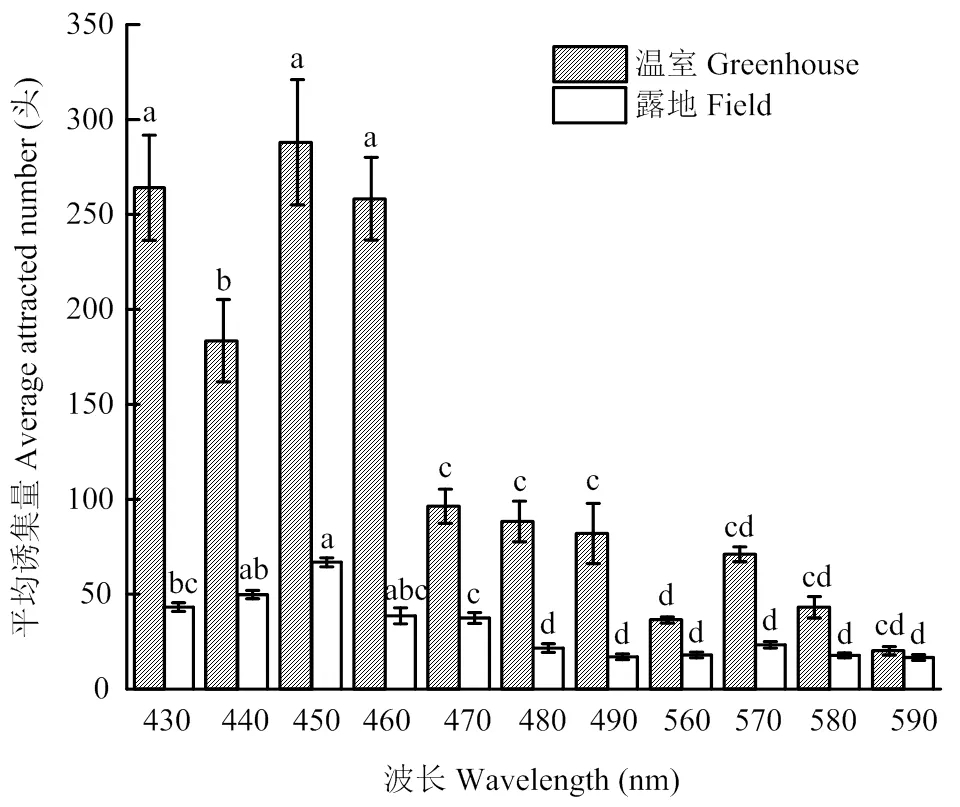
图9 不同虚拟波长下诱虫色板诱集烟蓟马的平均诱集量
露地试验结果表明,黄色和蓝色诱虫色板对烟蓟马的诱集作用存在显著差异,且蓝色诱虫色板的诱集量显著高于黄色诱虫色板。其中诱虫色板对烟蓟马的诱集数量大小依次为2#>1#>3#>5#>6#>4#>7#。2#蓝色诱虫色板对烟蓟马的诱集效果最好,平均诱集数量为(99.19±3.18)头/板,显著高于其他2种蓝色色板和4种黄色色板(表2)。

表2 不同生产厂家诱虫色板平均诱集量
不同生产厂家诱虫色板诱集试验结果表明,无论是在温室还是露地,蓝色色板均能诱集到较多的烟蓟马,诱集量显著高于黄色诱虫色板的诱集量,且波长在440—470 nm的蓝色色板诱集效果最好,这与室内筛选的最佳波长一致。
3 讨论
波长和光照强度是影响烟蓟马趋光的重要因子。本研究结果表明,450 nm的蓝色光对烟蓟马的吸引力最强,趋光率高达75%,562 nm的黄色光趋光率次之,为74%,430 nm蓝紫色光的趋光率64%,研究结果与陈祯[23]室内测定结论一致,即烟蓟马对450 nm的蓝色光有很强的趋性。室内趋光行为测定显示烟蓟马对黄色光有较强的趋性,但在田间试验并没有得到验证,这可能与烟蓟马所处的小环境或生理状况等有关,不能简单的将昆虫的趋光性与温室或露地的同种昆虫对光、颜色偏好等同[24]。活动能力较强的昆虫其复眼有较强的自我调节能力,通常在行为上表现比较耐光[25-29],其表现为在光的刺激下,昆虫行为随着光照强度的变化而变化。本研究发现,在562 nm的黄色光刺激下,即使在很弱的光照强度下,烟蓟马的趋光性也在增加,这更加能证明烟蓟马复眼对光具有调节能力,也可能与烟蓟马常常隐秘于暗处的生活习性有关。Makabe等[30]对烟蓟马的复眼进行电生理敏感光谱测定,结果显示烟蓟马对362 nm的紫外光以及532 nm的绿色光最为敏感,与本试验结果不一致,原因可能是视觉信号在加工传导时引起了视觉偏移。昆虫在接收到光信号后,经过神经传导、整合、加工的过程受到外界环境的影响,易造成行为反应结果与电生理测定结果不一致。如西花蓟马电生理测定峰值与趋光行为光谱峰值相差20 nm;铜绿丽金龟()对单色波选择的行为测试和复眼电生理不完全一致,峰值相差5 nm。复眼电生理测定采用胞外和胞内电位的方法,光信号未经神经输出,因此不能清楚地说明昆虫的感光生理机制。显然,趋光行为测定的方法更能直观反应昆虫对光和色的敏感性。
探究影响烟蓟马趋光行为因子是绿色高效防控其发生和危害的必要工作。本试验室内结果表明,烟蓟马雌虫的趋光性高于雄虫,这可能与雌虫生理节律有关,雌虫需要不断的补充营养物质、产卵以及寻找繁殖地[31]。无论是雌虫还是雄虫,450 nm的蓝色光均可以引诱到较多的烟蓟马。Davidson等[32]报道西花蓟马在饥饿4 h后飞行能力增强。本研究也表明短时间的饥饿可以增强昆虫的活动能力,烟蓟马饥饿4 h后对单色光的敏感性增强,但后期随着饥饿时间的延长和能量的消耗,对单色光的趋性降低。对烟蓟马在不同时间段趋光行为影响研究表明,烟蓟马在上午8:30—10:00对单色光的趋性最高,中午11:30—13:00减弱,下午15:00—16:30又稍加增强,临近黄昏时,趋光性再次降低,Smith等[33]在葱上也观察到相似的结果。烟蓟马具有栖花性,大多数的植物花朵开放于早晨,再加上烟蓟马经过一晚的饥饿,造成烟蓟马在早晨对单色光敏感。黄昏时,烟蓟马趋光性下降的原因尚不明确。烟蓟马在中午和下午的趋光行为可能受到温度和相对湿度的双重影响。温度为25—30℃时,烟蓟马的趋光性显著增强。最适于烟蓟马趋光的是相对湿度为60%—75%。此外,有关烟蓟马趋光性行为影响因子的研究仅在室内条件下开展,其数据还需要在田间进一步验证。
本质上讲,物质的颜色是通过光反射而得到的,研究昆虫的趋光性在一定的程度上就是研究昆虫趋色性[34]。但是,目前关于烟蓟马趋性的精确颜色及波长尚未定论。Kirk[35]对烟蓟马进行田间试验发现白色诱虫色板的诱集量是黄色和蓝色色板诱集量的两倍;Lu[36]报道在温室条件下,烟蓟马对蓝色以及淡蓝色诱虫色板的趋性要高于黄色和白色诱虫色板,这与本试验的结果一致。应用虚拟波长与RGB值的函数关系,将颜色/波长定量化来研究昆虫趋光行为是一种便捷且精确的研究方法,可实现诱虫色板相关参数数字化、规范化。此方法已运用在瓜实蝇()[37]、橘小实蝇()[38]、椰心叶甲()[39]、豆大蓟马()[40]、花蓟马()[41]、茶黄蓟马()[42]等,以评价对昆虫具有最佳趋性的颜色/波长。本研究表明,在黄色波段内,570 nm的黄色诱虫板对烟蓟马的吸引作用最强,其RGB值为R=225、G=255、B=0。在蓝色波段内,烟蓟马对450 nm的蓝色诱虫板(R=0、G=70、B=255)具有最佳的趋性,这与室内光谱趋性试验的结果一致。
目前,国内外关于昆虫趋光性的试验仅在单一的室内或室外条件下开展,且相关影响因子不明,本研究通过室内单色光试验得出,烟蓟马对450 nm的蓝色光表现出了最强的趋向反应,温室和露地试验也验证了450 nm的蓝色诱虫色板对烟蓟马有强烈的吸引作用。研究结果为研发蓝色诱虫色板以及光源诱捕器高效防治烟蓟马提供了理论支持。有关烟蓟马趋光行为影响因子的研究有助于改进和提高诱集防治技术,增强防治效果。
4 结论
光波长对烟蓟马成虫的趋光性有着较大的影响,光照强度的变化亦影响着烟蓟马的趋光行为。昆虫自身的感光机能和外界环境决定其趋光性,互作关系比较复杂。烟蓟马的趋光行为不仅受到内在因素如性别、饥饿程度的制约,也受到外在因素如光波长、光照强度、日节律、温度和相对湿度的影响。实验室、温室及露地试验结果表明,烟蓟马对450 nm的单色光以及450 nm蓝色诱虫色板有最佳的趋性,可用于烟蓟马的监测与防治。
[1] Rueda A, Badenes-Perez F R, Shelton A M. Developing economic thresholds for onion thrips in Honduras., 2007, 26: 1099-1107.
[2] 谢永辉, 李正跃, 张宏瑞. 烟蓟马研究进展. 安徽农业科学, 2011, 39(5): 2683-2685, 2785.
XIE Y H, LI Z Y, ZHANG H R. Research advances on the onion thrips ()., 2011, 39(5): 2683-2685, 2785. (in Chinese)
[3] den Belder E, Elderson J, van den Brink W J, Schelling G. Effect of woodlots on thrips density in leek fields: a landscape analysis., 2002, 91(1/3): 139-145.
[4] Muvea A M, Meyhofer R, Subramanian S, Poehling H M, Ekesi S, Maniania N K.Colonization of onions by endophytic fungi and their impacts on the biology of., 2014, 9(9): e108242.
[5] DIAZ-MONTANO J, FUCHS M, NAULT B A, FAIL J, SHELTON A M. Onion thrips (Thysanoptera: Thripidae): A global pest of increasing concern in onion., 2011, 104(1): 1-13.
[6] 韩云, 唐良德, 吴建辉. 蓟马类害虫综合治理研究进展. 中国农学通报, 2015, 31(22): 163-174.
HAN Y, TANG L D, WU J H. Researches advances on integrated pest management of thrips (Thysanoptera)., 2015, 31(22): 163-174. (in Chinese)
[7] Martin N A, Workman P J, Butler R C. Insecticide resistance in onion thrips () (Thysanoptera: Thripidae)., 2003, 31(2): 99-106.
[8] MacIntyre Allen J K, Scott-Dupree C D, Tolman J H, Harris C R. Resistance ofto pyrethroid and organophosphorus insecticides in Ontario, Canada., 2005, 61(8): 809-815.
[9] Herron G A, James T M, Rophail J, Mo J. Australian populations of onion thrips,Lindeman (Thysanoptera: Thripidae), are resistant to some insecticides used for their control., 2008, 47: 361-364.
[10] Herron G A, Langfield B J, Tomlinson T M, Mo J. Dose-response testing of Australian populations of onion thripsLindeman (Thysanoptera: Thripidae) further refines baseline data and detects methidathion and likely imidacloprid resistance., 2011, 50(4): 418-423.
[11] Lebedev G, Abo-Moch F, Gafni G, Ben-Yakir D, Ghanim M. High-level of resistance to spinosad, emamectin benzoate and carbosulfan in populations ofcollected in Israel., 2013, 69(2): 274-277.
[12] 靖湘峰, 雷朝亮. 昆虫趋光性及其机理的研究进展. 昆虫知识, 2004, 41(3): 198-203.
JING X F, LEI C L. Advances in research on phototaxis of insects and the mechanism., 2004, 41(3): 198-203. (in Chinese)
[13] 孙兴全, 刘晓平, 陆军. 棉花烟蓟马的发生与综合防治措施. 安徽农学通报, 2008, 14(24): 109.
SUN X Q, LIU X P, LU J. Investigations on the occurrence and measurement of comprehensive prevention ofLindeman(Thysanoptera: Thripidae) in cotton., 2008, 14(24): 109. (in Chinese)
[14] Mautino G C, Bosco L, Tavella L. Integrated management of(Thysanoptera: Thripidae) on onion in north-western Italy: basic approaches for supervised control., 2012, 68(2): 185-193.
[15] Khaliq A, Khan A A, Afzal M, Tahir H M, Raza A M, Khan A M. Field evaluation of selected botanicals and commercial synthetic insecticides againstLindeman (Thysanoptera: Thripidae) populations and predators in onion field plots., 2014, 62: 10-15.
[16] CHEN Z, GE Y, LIU X, KUANG R. Effect of colored sticky cards on non-target insects., 2015, 16(5): 983-987.
[17] Hoddlea M S, Robinsona L, Morgan D. Attraction of thrips (Thysanoptera: Thripidae and Aeolothripidae) to colored sticky cards in a California avocado orchard., 2002, 21(5): 383-388.
[18] 吴青君, 徐宝云, 张友军, 张治军, 朱国仁. 西花蓟马对不同颜色的趋性及蓝色粘板的田间效果评价. 植物保护, 2007, 33(4): 103-105.
WU Q J, XU B Y, ZHANG Y J, ZHANG Z J, ZHU G R. Taxis of western flower thrips to different colors and field efficacy of the blue sticky cards., 2007, 33(4): 103-105. (in Chinese)
[19] Rodriguez-Saona C R, Polavarapu S, Barry J D, polk d, jornsten r, oudemans p v, liburd o e. Color preference, seasonality, spatial distribution and species composition of thrips (Thysanoptera: Thripidae) in northern highbush blueberries., 2010, 29(11): 1331-1340.
[20] Demirel N, Yildirim A E. Attraction of various sticky color traps toLindeman (Thysanoptera: Thripidae) andPaoli (Homoptera: Cicadellidae) in cotton., 2008, 5(6): 389-394.
[21] Gharekhani G H, Ghorbansyahi S, Saber M, Bagheri M. Influence of the colour and height of sticky traps in attraction of(Lindeman) (Thysanoptera, Thripidae) and predatory thrips of family Aeolothripidae on garlic, onion and tomato crops., 2014, 47(18): 2270-2275.
[22] 范凡, 任红敏, 吕利华, 张莉萍, 魏国树. 光谱和光强度对西花蓟马雌虫趋光行为的影响. 生态学报, 2012, 32(6): 1790-1795.Fan F, Ren H M, Lü L H, Zhang L P, Wei G S. Effect of spectral sensitivity and intensity response on the phototaxis of(Pergande)., 2012, 32(6): 1790-1795. (in Chinese)
[23] 陈祯. 昆虫趋光特性与粘虫色板应用的生态影响——以烟田害虫及其天敌昆虫的趋光性研究为例[D]. 昆明: 云南大学, 2016.
CHEN Z. Insect phototaxis and ecological impacts of application of colored sticky cards in tobacco field[D]. Kunming: Yunnan University, 2016. (in Chinese)
[24] Prokopy R J, Owens E D. Visual detection of plants by herbivorous insects., 1983, 28: 337-364.
[25] 魏玮. 两种草蛉成虫复眼显微结构及其趋光行为的研究[D]. 保定: 河北农业大学, 2009.
WEI W. Study on microstructure of the compound eye and phototactic behavior ofTjeder andBrauer (Neuroptera: Chrysopidae)[D]. Baoding: Hebei Agricultural University, 2009. (in Chinese)
[26] 张海强, 朱楠, 范凡, 魏国树. 大草蛉成虫复眼的外部形态及其显微结构. 昆虫学报, 2007, 50(5): 454-460.
ZHANG H Q, ZHU N, FAN F, WEI G S. External morphology and microstructure of the compound eye ofRamber (Neuroptera: Chrysopidae)., 2007, 50(5): 454-460. (in Chinese)
[27] 江幸福, 张总泽, 罗礼智. 草地螟成虫对不同光波和光强的趋光性. 植物保护, 2010, 36(6): 69-73.
JIANG X F, ZHANG Z Z, LUO L Z. Phototaxis of the beet webwormto different wavelengths and light intensity., 2010, 36(6): 69-73. (in Chinese)
[28] 陈晓霞, 闫海燕, 魏玮, 乔玮娜, 魏国树. 光谱和光强度对龟纹瓢虫成虫趋光行为的影响. 生态学报, 2009, 29(5): 2349-2355.
CHEN X X, YAN H Y, WEI W, QIAO W N, WEI G S. Effect of spectral sensitivity and intensity response on the phototaxis of(Thunberg)., 2009, 29(5): 2349-2355. (in Chinese)
[29] 闫海燕. 龟纹瓢虫(Thunberg)感光、趋光机制的研究[D]. 保定: 河北农业大学, 2006.
YAN H Y. Study on mechaniam of photosensitity and phototaxis of(Thunberg)[D]. Baoding: Hebei Agricultural University, 2006. (in Chinese)
[30] MAKABE T, FUTAMURA T, Takayosh N, WAKAKUWA M, ARIKAWA K. Phototaxis of western flower thrips,and onion thrips,and the possibility of controlling thrips using ultraviolet-emitting trap in the greenhouse of Satsuma Mandarin ()., 2014, 58(3): 187-195.
[31] SONTIGUN N, SUKONTASON K L, KLONG-KLAEW T, SANIT S, SAMERJAI C, Somboon P, Thanapornpoonpong S, Amendt J, Sukontason K. Bionomics of the oriental latrine fly(Fabricius) (Diptera: Calliphoridae): temporal fluctuation and reproductive potential., 2018, 11: 415.
[32] Davidson M M, Butler R C, Teulon D A J. Starvation period and age affect the response of female(Pergande) (Thysanoptera: Thripidae) to odor and visual cues., 2006, 52(7): 729-736.
[33] Smith E A, Shields E J, Nault B A. Impact of abiotic factors on onion thrips (Thysanoptera: Thripidae) aerial dispersal in an onion ecosystem., 2016, 45(5): 1115-1122.
[34] 雷朝亮, 荣秀兰. 普通昆虫学. 北京: 中国农业出版社, 2003: 142.
LEI C L, RONG X L.. Beijing: China Agriculture Press, 2003: 142. (in Chinese)
[35] KIRK W D J. Ecologically selective coloured traps., 1984, 9: 35-41.
[36] Lu F M. Color preference and using silver mulches to control the onion thrips,Lindeman., 1990, 10(3): 337-342.
[37] Xue H w, Wu W j. Preferences of(Diptera: Tephritidae) to different colors: a quantitative investigation using virtual wavelength., 2013, 56(2): 161-166.
[38] Wu W Y, Chen Y P, Yang E C. Chromatic cues to trap the oriental fruit fly,., 2007, 53(5): 509-516.
[39] 陈俊谕, 马光昌, 陈泰运, 符悦冠. 椰心叶甲对虚拟波长下不同颜色的选择行为. 热带作物学报, 2014, 35(5): 962-966.
CHEN J Y, MA G C, CHEN T Y, FU Y G. Preferences ofto different virtual wavelengths colors., 2014, 35(5): 962-966. (in Chinese)
[40] 唐良德, 韩云, 吴建辉, 李鹏, 付步礼, 邱海燕, 刘奎. 豆大蓟马室内对不同颜色及光波的趋性反应. 植物保护, 2015, 41(6): 169-172.
TANG L D, HAN Y, WU J H, LI P, FU B L, QIU H Y, LIU K. Preference of(Thysanoptera: Thripidae) to different colors and light-waves in lab., 2015, 41(6): 169-172. (in Chinese)
[41] 张玲. 新疆苜蓿田蓟马种类、发生规律与温度对花蓟马生长发育影响的研究[D]. 乌鲁木齐: 新疆农业大学, 2007.
ZHANG L. Study on species, the occurrence of thrips on alfalfa in Xinjiang and the influence of temperature on the growth and development to[D]. Urumqi: Xinjiang Agricultural University, 2007. (in Chinese)
[42] 王琛, 朱文静, 符悦冠, 解志峰, 韩冬银, 张方平, 牛黎明. 茶黄蓟马嗜好颜色筛选及监测效果测定. 环境昆虫学报, 2015, 37(1): 107-115.
WANG C, ZHU W J, FU Y G, XIE Z F, HAN D Y, ZHANG F P, NIU L M. Screening of preferred color and field evaluation for monitoringHood (Thysanoptera: Thripidae)., 2015, 37(1): 107-115. (in Chinese)
(责任编辑 岳梅)
The Phototaxis Behavior ofand Trapping Effect of Different Wavelength Sticky Cards in the Field
MI Na, Zhang QiKai, WANG HaiHong, WU ShengYong, LEI ZhongRen
(State Key Laboratory for Biology of Plant Diseases and Insect Pests, Institute of Plant Protection, Chinese Academy of Agricultural Sciences/Key Laboratory of Prevention and Control of Biological Hazard Factor (Plant Origin) for Agrifood Safety and Quality, Ministry of Agriculture and Rural Affairs, Beijing 100193)
【Objective】The objective of this study is to explore the optimum light wavelength, light intensity and related factors, such as sex, daily rhythm, starvation time, temperature and relative humidity ofthrough behavioral responses study under the different conditions of laboratory, greenhouse and field, and to provide a theoretical support for improving the control techniques and products of, such as color sticky cards and light traps. 【Method】Firstly, the behavioral responses ofto 10 different wavelengths of monochromatic light were conducted by using monochromator testing device with high resolution, wide spectrum measurement range and high stability under laboratory condition. Monochromatic light with high response rate was screened and the initial light intensity was attenuated by neutral density filter to explore the effect of light intensity on the behavioral responses of. Secondly, the effects of sex, daily rhythm, starvation time, temperature and relative humidity on phototaxis ofwere also examined by using the rate of phototaxis as statistical index. Based on laboratory experiments, using the relationship between Dan Bruton’s virtual wavelength and RGB values, the blue and yellow sticky cards of the corresponding wavelengths were printed and the plates were glued. The tendency ofto the self-made sticky cards of different wavelengths and sticky cards produced by different manufacturers was evaluated in the greenhouse and in the open field.【Result】The results of laboratory experiment showed that phototaxis behavioral response rate was the highest on blue light at 450 nm, and the rate of photaxis was as high as 75.34%, followed by the yellow light at 562 nm and the blue-purple light at 430 nm, the rate of photaxis was 73.61% and 64.03%, respectively. Under the stimulation of 430, 450 and 562 nm monochromatic light, the phototaxis rate of femalewas higher than that of male. The light intensity attenuation test of the three monochromatic light showed that the tendency ofincreased with light intensity. At 8: 30-10: 00 am, thewas most sensitive to monochromatic light at 430, 450, and 562 nm. After 4 hours of starvation, the phototaxis rate ofwas the strongest, and then decreased with the prolongation of starvation time. The phototaxis rate ofto three monochromatic light was significantly higher than that of the control at 25-30℃. at 15℃,was not sensitive to monochromatic light stimulation. Under the relative humidity of 45%-60%, the phototaxis rate ofwas significantly stronger than that of the control at 430, 450 and 562 nm. However, there was no significant difference with the control under the relative humidity of 30% and 90%. The trapping effect onwas evaluated by using different wavelength sticky cards and different manufacturers in the greenhouse and field, and the results showed that the blue self-made sticky cards and the No.2 sticky cards with reflection wavelength of 440-470 nm had the best effect. The results of this test were in agreement with the optimum wavelength of laboratory screening. 【Conclusion】Sex, daily rhythm, starvation, temperature and relative humidity all have certain influence on phototaxis of. Comprehensive analysis on laboratory, greenhouse and field revealed thathas obvious tendency to monochromatic light and sticky cards at 450 nm. The blue trap sticky card of about 450 nm can be used to monitor and control.
; phototaxis; monochromatic light; color sticky card
10.3864/j.issn.0578-1752.2019.10.006
2018-12-05;
2019-01-04
国家重点研发计划(2017YFD0200900)、国家现代农业产业技术体系专项资金(CARS-23-D-08)
米娜,E-mail:3385856294@qq.com。通信作者雷仲仁,E-mail:leizhr@sina.com
猜你喜欢
植物保护(2022年5期)2022-10-13
杭州电子科技大学学报(自然科学版)(2022年3期)2022-06-08
今日农业(2021年11期)2021-11-27
阅读(科学探秘)(2021年8期)2021-09-01
青年歌声(2021年2期)2021-03-05
青年歌声(2020年11期)2020-11-24
陕西农业科学(2019年12期)2019-06-16
河南农业科学(2019年4期)2019-04-23
中国科学院大学学报(2019年1期)2019-01-21
商周刊(2018年13期)2018-07-11
