
毛竹入侵对森林植物和土壤的影响研究进展
2019-07-08 07:13周本智姜丽娜曹永慧葛晓改杨振亚
生态学报 2019年11期
童 冉,周本智,姜丽娜,曹永慧,2,葛晓改,2,杨振亚,2
1 中国林业科学研究院亚热带林业研究所,杭州 311400 2 钱江源森林生态系统国家定位观测研究站,杭州 311400 3 南京林业大学,南京 210000
森林生态系统作为陆地生态系统的主体部分,正在经历着火灾[1]、极端气候事件[2]、植物入侵[3]等危害。目前,对于植物入侵的研究大都集中于对草本、灌木类植物,而对于乔木类植物入侵研究较少。毛竹(Phyllostachysedulis(Carrière)J.Houzeau)作为重要的笋材两用经济树种,早在20世纪90年代,人们就认识到种植毛竹对农村经济的巨大贡献,这也是毛竹在短时间内得到大面积种植的重要原因[4]。为了追求利益最大化,在种植初期,竹农对于毛竹向原有森林扩张持乐观态度。但随着时代的发展和进步,廉价劳动力日渐缺乏,导致种植毛竹的这种强集约型经济模式被逐渐摒弃。在管理强度大幅降低之后,毛竹扩张问题愈发严重,而随着人们对生态环境保护的意识空前加强,越来越多的科技工作者率先认识到毛竹入侵原有森林所带来的巨大生态效益损失,开始积极开展相关研究工作。
毛竹是亚热带地区分布最为广泛的竹种,其人工林栽培面积正在逐年增加,笋材两用的特性为林农带来了极佳的经济效益。另外,随之而来的生态效益也逐渐增大,这与毛竹庞大的地下系统能够很好地起到含蓄水源、固定山地边坡的作用以及巨大的地上部分生物量又是森林碳汇的重要组成部分有关[5]。与此同时,这种人为导致的毛竹林纯化对周边森林进行入侵而产生的问题愈发明显,已经严重威胁到森林的物种多样性和生态系统稳定性[6],甚至在日本某些气候条件适宜的地区,由于极强的繁殖能力和高的光合作用速率等特性成为了外来入侵物种,面积的不断扩张也给生态系统服务功能造成了影响[7]。本文结合国内外研究进展及初期的阶段性研究,归纳和总结了毛竹林纯化及外来引种导致的入侵对原有森林产生的负面影响和潜在影响,并结合自然状态下毛竹扩张策略等从侧面进行了补充阐述。
植物和土壤是森林对外来侵害最为敏感的部分,两者对毛竹入侵的响应能够体现森林生态系统自身调控的适应能力。结合国内外毛竹入侵森林的相关研究,本文系统地综述了毛竹入侵森林生态系统的机制,植物生存能力、物种多样性、植被碳储量,以及土壤理化性质,土壤微生物群落结构和功能等对毛竹入侵响应与适应性的研究结果,在此基础上指出当前研究存在的问题,并提出未来的研究展望。这将有助于理解森林植物、土壤与入侵植物的相互关系以及植物、土壤对植物入侵的响应与适应对策,对系统了解植物入侵,预测周边环境对植物入侵的响应与适应具有一定的意义。
1 毛竹入侵森林的机制研究
在易于受到人为干扰的半自然森林发生植物入侵的可能性很大,这与半自然森林结构与功能不够完整有关,而大量研究表明植物入侵与物种本身的特性也有很大关系[8-9]。毛竹地下根状茎竹鞭极强的繁衍和空间拓展能力是其入侵周边森林的先决条件,有利于毛竹纯林在与针叶林、阔叶林、针阔混合林竞争中取得优势。Okutomi等[10]总结了阔叶林完全被毛竹纯林的过程,发现旺盛的根状茎入侵是最先开始的。毛竹在入侵过程中根系具有形态可塑性,即通过增加特殊根长、减少侧根节间长度,已达到向周围森林扩展的目的[11]。竹笋的伸长在很大程度上不受光环境的影响,这表明笋的伸长很可能依赖于无性系根系的碳水化合物的供给,新梢无性系对树荫的耐受性是其入侵完整林分的关键因素[12]。幼龄竹秆能够穿透林分树冠迅速形成新的树冠,产生强烈的遮光效应[10]。毛竹茎干快速生长的特性使其对碳水化合物的需求量极大[13-14],间接使入侵林木逐渐失去赖以生存的水、光条件,从而形成了竹进林退的局面。白尚斌等[15]用毛竹茎叶、根和凋落物等浸取液处理苦槠、青冈种子进行种子萌发效应的测定,发现高浓度浸提液对两树种发芽速度有显著的延缓作用,对苦槠根系活力有显著的抑制作用,说明毛竹具有潜在的化感作用,可能有利于其入侵周边森林,从而危害周边森林环境。改变土壤元素矿化进程也可能是毛竹成功入侵的重要策略[16]。
毛竹在入侵森林的同时,自身也进化出新的适应策略,以更好的应对外界不断变化的环境条件。刘希珍等[17]在对毛竹入侵阔叶林过程中叶功能性状特性的研究发现,叶表面积与叶干物质含量之间呈显著负相关关系,N与P呈显著正相关关系,Liu等[18]对毛竹入侵杉木林进行了研究,发现N和P的含量随着毛竹比例的增加呈现相同的变化趋势。另外,两者都认为毛竹年龄是影响叶功能性状采取不同适应策略的关键因素[17-18]。毛竹根系在入侵森林过程中同样也作出了积极的适应,刘骏等[19]对毛竹向常绿阔叶林扩张过程中细根策略的研究发现,毛竹在与阔叶树竞争的过程中,细根分布趋向于上层土壤,且比根长显著增加,通过这种广布、精确的细根竞争策略实现资源获取能力的提高,达到种群扩张的目的。毛竹胸径和生物量、叶面积指数和凋落物数量和质量等群落结构指标在入侵不同类型森林时的变化特征不同,毛竹向阔叶林扩展时,通过增大个体胸径和叶面积指数来适应竞争环境,扩展前沿较多的凋落物量和较高的C、N、P含量,增加了养分归还潜力,会进一步促进毛竹的生长;毛竹向杉木林扩展时,通过增加叶面积指数来适应竞争环境,随着毛竹林的扩展毛竹胸径增加、凋落物C∶P、N∶P增加,毛竹生长受到P的制约减弱[20]。Saroinsong等对落叶阔叶林附近的毛竹林分进行了为期12年的监测,发现在入侵过程中研究区内竹秆密度和地上生物量增大,而阔叶树密度和物种数量都减小[21]。综上所述,毛竹在入侵森林生态系统时,地下和地上部分发生着协同改变,又进一步而影响到周边森林的植物和土壤。
2 毛竹入侵对植物的影响
2.1 影响植物生存能力
植物在对原生地群落入侵的过程中,最先也是最直接受到攻击的是原有植物。入侵植物通过掠夺养分和水分、争取更充足的光照、分泌化学物质等方式影响原生地植物的生存能力,对植物正常生长产生威胁。刘烁等[22]发现毛竹入侵会使杉木林变得更为稀疏,光照强度增强,但理论上并未影响到成年杉木的正常生长,而由于杉木幼苗喜荫的特性,导致幼苗更新受阻,可能是导致杉木林死亡退化的重要原因。Okutomi K等[10]对毛竹入侵阔叶树原因分析时,发现毛竹竹秆能够轻易对临近的阔叶树造成机械损害。毛竹各器官潜在的化感作用也可能使林木种子萌发困难,根系难以伸长[15]。
2.2 降低植物物种多样性
植物物种多样性是指植物群落中的丰富性和异质性,即群落中所含物种的多寡和群落中各个种的相对密度[23]。植物入侵通过减少原有群落的植物物种多样性,对食物网进行破坏,进而对生态系统结构和功能造成损害[24]。林倩倩等[25]发现毛竹入侵造成了常绿阔叶林、针叶林、针阔混交林等森林类型树种立木数的降低。另外,不同生活型植物物种多样性对毛竹入侵的响应不同。白尚斌等[6]对天目山自然保护区进行了为期7年的长期定位观测实验,发现毛竹入侵对针阔混交林植物物种多样性产生了不利影响,具体表现在乔木层和灌木层的物种丰富度、Simpson指数和Pielou均匀度指数显著降低,而草本层的物种丰富度显著提高。乔木层植物物种多样性受毛竹入侵影响最大,欧阳明等[26]、杨怀等[27]在研究毛竹向常绿阔叶林扩张时均发现乔木层的Shannon-Wiener指数发生了大幅下降。毛竹入侵导致周围森林生物多样性的降低,进而加剧毛竹入侵形势,影响森林生态功能的稳定。
2.3 减少植被碳储量
植被在全球碳循环中具有重要的作用,可以通过光合作用在极短时间尺度上实现对碳循环的调节[28]。森林植被碳库占全球陆地的77%,在维持全球碳平衡以及调节气候方面发挥着不可替代的作用[29]。而当植物入侵发生时,一般会对原有森林碳循环产生影响,进而导致植被碳储量变化,这将是影响森林碳储量增减的重要因素[30]。Fukushima等[31]在对毛竹入侵日本东京次生阔叶林的研究发现,植物地上生物量与地下生物量的比值随毛竹优势度的增加而降低,尤其是在入侵初期。杨清培等[32]在研究大岗山毛竹入侵常绿阔叶林生态系统时发现植被碳储量出现大幅降低,由93.26 t/hm2减少到82.06 t/hm2,毛子龙等[33]同样发现在薇甘菊入侵深圳湾红树林生态系统后,植被碳储量也出现了大幅降低。另外,杨清培等[32]的研究发现植被的年固碳量有所增加,由常绿阔叶林的4.15 t hm-2a-1增加到毛竹林的4.76 t hm-2a-1,提高了14.18%,与其他种类植物入侵一般导致年固碳量降低不同,这可能与毛竹叶片数量庞大、光合作用速率高有关系。
3 毛竹入侵对土壤理化性质和微生物的影响
3.1 对土壤物理性质的影响
植物在入侵周边森林过程中,由于入侵物种和所取代物种根系之间存在生理特性的差异,导致土壤物理性质发生改变,如土壤含水量、容重、孔隙度、密度等,而这种变化往往有利于加快植物入侵的进程。赵雨虹等[34]通过设置4种不同的林分类型对毛竹入侵常绿阔叶林过程中土壤物理性质进行了研究,发现两种混交林林分土壤密度大于阔叶林,而土壤孔隙度变小,持水能力变弱。宋庆妮等[16]则发现竹阔混交林相较于常绿阔叶林,在0—10 cm和10—30 cm两个土层的土壤容重都分别显著减少。Shinohara等[35]对毛竹林和临近常绿阔叶林的土壤含水量进行了对比,发现在0—60 cm的土层范围内,竹林土壤体积含水量始终高于阔叶林,说明毛竹入侵可能会导致土壤含水量的变化。综合来看,由于原有常绿阔叶林属于高等级植物群落类型,土壤物理性质在长时间的群落进化过程中趋于稳定,所以当发生毛竹入侵时,往往会破坏原有的土壤结构和功能,对原有森林造成不利影响,而当毛竹入侵完成时,土壤物理性质又趋于稳定,向有利于毛竹生长的方向发展。
3.2 对土壤化学性质的影响
土壤化学性质包括土壤酸碱性、有机质含量、养分周转、元素循环等,是森林生态系统物质循环和能量流动的重要保障,也是对外界环境变化反应较为敏感的部分。土壤酸碱度是土壤胶体固液相性质的综合表现,一般用pH值表示,是对外界环境影响反应敏感的土壤化学性质,能够对植物的生长发育产生影响[36]。外来植物入侵会导致土壤酸碱性的变化,Li 等[37]对毛竹入侵阔叶林后土壤化学性质进行研究,发现混交林土壤pH远高于毛竹纯林和未侵入森林。Umemura等[38]同样发现毛竹入侵日本扁柏形成混交林的土壤pH值要显著高于未被侵入森林,且pH值与土壤交换性钙离子含量之间呈现显著正相关关系,而与钾、镁离子之间相关性不大,说明毛竹可能是通过调节土壤中交换性钙离子含量对pH值进行调整,进而对其他土壤过程和土壤化学性质产生影响。
植物入侵往往能增加生态系统中土壤有机质含量,其中土壤有机碳占很大一部分。赵雨虹等[35]对毛竹入侵常绿阔叶林过程中土壤有机碳含量变化研究时,发现土壤有机碳含量先增大后降低,大小关系:毛竹纯林<常绿阔叶林<入侵混交林。而Wang等[39]却发现毛竹入侵日本柳杉成熟森林过程中土壤有机碳含量大小顺序依次是:毛竹纯林<入侵混交林<日本柳杉林,表明毛竹增加了有机物腐殖化进程,这可能是与毛竹高活性炭加快了凋落物的分解有关。从两者的研究结论可以看出,入侵混交林土壤有机碳含量是否高于原有森林可能与入侵林分类型有关,其中凋落物类型可能是造成差异的主要原因。毛竹纯林土壤有机碳含量一般都小于入侵森林,杨清培等在进行毛竹扩张对常绿阔叶林生态系统碳储特征影响的研究同样得出了这样的结论[32]。吴家森等[40]对毛竹入侵森林对土壤肥力影响时,发现各活性有机碳占总有机碳的百分率也随着毛竹的扩张而增大。
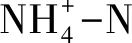
土壤磷元素缺乏对我国亚热带地区林木生长起着严重的限制作用,包括针叶林、常绿阔叶林、真阔混交林等,所以对毛竹入侵森林边缘地带磷元素动态的研究是很有必要的[42]。Wu等[43]对毛竹入侵日本柳杉人工林后的土壤磷元素进行了24个月的动态观测,发现混交林土壤全磷含量显著低于毛竹纯林和日本柳杉人工林,而毛竹纯林和日本柳杉人工林的土壤有效磷却显著降低。混交林酸性磷酸酶活性显著高于另外两种林分,土壤微生物生物量磷同样高于另外两种林分,但并不显著。在两年的测定中,以上4个土壤指标在混交林中都呈现季节性波动,而在另外两种林分中却保持相对稳定。
3.3 对土壤微生物群落与功能的影响
土壤微生物作为森林生态系统重要的组成部分,能够对土壤生态系统养分流动和能量流动起到主导作用,在维持系统稳定性和可持续性方面具有重要意义[44]。当外界环境发生变化时,土壤微生物种类多样性[45]、生物量[46]、群落结构[47]、功能多样性[48]等都会发生相应的变化。由于土壤微生物结构组成相对简单的特性,当发生外界植物入侵等类型的生态扰动时,会进行快速而灵敏的应答反应[49]。李永春等[50]在对毛竹入侵天目山原始阔叶林对土壤真菌群落的影响时发现,硝态氮和pH是影响真菌群落结构和数量差异的主要因素,且毛竹入侵显著影响了阔叶林原有的生化过程,这与Chang等[51]研究了毛竹入侵对日本柳杉人工林真菌群落影响的结论相同。王奇赞等[52]和Xu等[53]分别用不同的方法分析毛竹入侵天目山常绿阔叶林土壤细菌群落结构造成的影响,前者发现毛竹入侵并未显著改变土壤细菌结构和多样性的显著改变,而后者却得出毛竹入侵显著影响到土壤微生物群落,而两者共同认为,土壤中未知的细菌种类增多,这可能是由于试验方法和计量的不同,造成了结论不尽相同。Lin等[54]对毛竹入侵日本柳杉人工林造成的影响进行研究,发现竹林土壤细菌群落多样性最多样,其次是过渡带,杉木人工林的多样性最低,说明毛竹入侵邻近的日本柳杉人工林,增加了土壤细菌多样性,进而改变了土壤微生物群落。
4 存在问题和研究展望
4.1 丰富毛竹入侵管控手段
目前,越来越多的人意识到毛竹入侵对森林生态系统造成的负面影响,迫切需要投入了更多的人力和物力进行毛竹入侵的管控。传统的手段有在入侵林地内将毛竹进行清除,但往往效果不好。蔡亮等[55]发现在其研究区内,毛竹竹鞭一般只有向前或向下的生长趋势,当遇到岩石、断面、水流和强根时并不会向侧面生长,进而提出挖沟、灌水的初步管控策略。加大采笋量、砍伐量同样也可以对毛竹长势造成一定的影响。日本人T Uemura等提出在经营性毛竹人工林时,进行平行条状采伐,能够在有效利用森林资源的同时,很好的起到防控作用。以上方法普遍存在周期长、见效慢、投入高的缺点,针对这些,有人提出了竹类除草剂的研制,这将是未来一段时间的攻坚难点。最后,笔者大胆设想,在自然界中是否存在与毛竹能够和谐生存,互不干扰正常生长,且对毛竹与邻近森林之间起到阻隔作用的树种,这将是生物防治中生物替代的重要手段。
4.2 着重不同类型毛竹—森林临界面演替长期监测
由于生殖方式存在差异,毛竹入侵往往是通过地下竹鞭的无性繁殖渗透到周边森林,具有一定的趋向性,而其他外来植物依靠自身风媒、虫媒传播等特性实现跨越式、散点式的侵入。毛竹这种入侵方式必然使毛竹与侵入森林之间形成临界面,而这个临界面自身所具有的物种组成、群落结构、微环境等特性决定了毛竹入侵的难易和速度。不同的森林类型又给对临界面研究增加了复杂性,其中是否存在特性和共性是以后研究的重点,而这需要我们投入更多精力进行临界面演替的长期监测。另外,利用遥感图像变化等先进手段从大尺度上投入到长期监测是很有必要的[56]。
4.3 明确毛竹入侵阶段划分
生物入侵的阶段划分一直存在争议,这往往是由于研究目的不同导致的。刘健等[57]将植物入侵划分为传播到达、定居建群和扩散入侵3个阶段,而这往往适用于外来植物入侵。除毛竹非原生种外的日本和其他亚热带地区外,中国大部分地区仅仅有扩散入侵这个最终阶段。针对竹类入侵自身特性,有人提出了以下4个阶段:地下渗透、地上成竹、竞争排外和优势维持[58]。祁红艳[59]认为毛竹氮磷根际效应要强于其他植物,只是并未阐明是地下渗透阶段还是竞争排外阶段。由于入侵森林类型不同,是否会造成入侵阶段周期延长或缩短,将是下一步研究的重点。另外,针对不同入侵阶段特性,提出具有针对性的防控措施也是很有必要的。
4.4 加强毛竹入侵生态后效研究
在已经完成毛竹入侵地区,我们不能只把注意力集中到负面影响,因为毛竹一旦入侵成功,其强大的生命力使其难以被彻底根除。此时,我们应该把更多注意力放到对生态后效的研究。毛竹入侵导致乔木、灌木物种多样性减少,反而增加了林下草本植物物种多样性,这是否有利于在已发生入侵的森林进行草本植物的种植,如药用植物沙参、决明等[60]。杨清培等[32]发现毛竹入侵后森林生态系统植被碳储量大幅降低,而年固碳量却有所升高。宋庆妮等[61]对毛竹入侵后凋落物水文功能进行评价后,发现常绿阔叶林转变为毛竹林后,凋落物的水文生态功能是增强的。毛竹庞大的地下根鞭系统具有极强的生物力学性质,对低海拔多雨地区山地边坡的稳定同样发挥着重要的作用,但是否优于原有林木尚无人研究。因此,对毛竹入侵生态效益评价将是今后的研究重点之一。
猜你喜欢
当代水产(2022年8期)2022-09-20
昆明医科大学学报(2022年2期)2022-03-29
食品安全导刊(2021年20期)2021-08-30
意林·少年版(2020年13期)2020-08-02
汉语世界(2020年3期)2020-06-19
东坡赤壁诗词(2020年2期)2020-06-04
河南科学(2020年3期)2020-06-02
绿色中国(2019年18期)2020-01-04
农家科技下旬刊(2017年8期)2017-11-13
科技创新导报(2016年21期)2016-12-17
