
鸦胆子素A诱导非小细胞肺癌细胞株H460凋亡及机制的初步研究
2019-08-20 02:23李聪王燕赵晓丽王真蒋建东
中国医药生物技术 2019年4期
李聪,王燕,赵晓丽,王真,蒋建东
鸦胆子素A诱导非小细胞肺癌细胞株H460凋亡及机制的初步研究
李聪,王燕,赵晓丽,王真,蒋建东
100050 北京,中国医学科学院北京协和医学院药物研究所(李聪、王燕、蒋建东),医药生物技术研究所(赵晓丽、王真)
考察鸦胆子素A(BA)对人非小细胞肺癌细胞系 H460 细胞凋亡的影响和作用机制。
用 BA 对 H460 细胞进行处理,采用 MTT 法检测细胞增殖情况,并利用 Annexin V-FITC/PI 双染法检测 BA 对 H460 细胞凋亡的影响。Hoechst 33342 染色和免疫荧光染色观察细胞凋亡时染色质的凝集以及 DNA 损伤标志蛋白 γ-H2AX 表达情况。Western blot 和RT-PCR 检测 H460 细胞内凋亡相关蛋白表达水平。
MTT 结果表明,与对照组相比,BA 能显著抑制 H460 细胞增殖。对 A549、H1299、H1355 三株非小细胞肺癌细胞系进行抑制增殖作用检测,结果显示 BA 对三株细胞系均有不同程度抑制作用。BA 作用 48 h 后,各浓度 BA 处理 H460 细胞凋亡率均高于对照组,具有时间依赖性,差异具有统计学意义。BA 处理组可见明显的染色质凝集,细胞核碎片化,DNA 损伤标志蛋白 γ-H2AX 表达明显增加。Western blot 结果显示,BA 作用 48 h 后,促凋亡蛋白 Bax 表达升高,抗凋亡蛋白 Bcl-2 表达降低,与对照组相比,Bax/Bcl-2 比值显著升高。同时与对照组相比,凋亡通路中的 cleaved caspase-9、cleaved caspase-3 表达明显增加,并引起凋亡标志蛋白 PARP 切割。RT-PCR 结果显示,BA 处理 H460 细胞后,与对照组相比,Bcl-2 基因表达显著降低。
BA 能抑制人非小细胞肺癌细胞株 H460 细胞增殖,显著诱导其凋亡,该作用与 BA 引起 DNA 损伤并激活线粒体凋亡通路有关。
鸦胆子素; 细胞凋亡; 癌,非小细胞肺; H460 细胞
肺癌是世界上发病率、死亡率最高的恶性肿瘤之一,其中非小细胞肺癌(non-small-cell lung cancer,NSCLC)占到 85%。统计显示,NSCLC 患者 5 年生存率极低(约 15%),原因是患者早期症状不明显,约有 75% 的患者发现时已处于中晚期[1],平均生存期仅为 4 个月[2]。目前的治疗方法主要是手术治疗、放化疗、免疫治疗、分子靶向治疗以及药物辅助治疗等[3-4]。手术方法和放化疗是临床常用治疗手段,但具有一定局限性;免疫治疗和分子靶向治疗,应用时间尚短,其安全性和有效性仍需进一步认证[5-6]。目前,从天然产物中发现抗肿瘤活性化合物是新药发现的主要途径,植物来源的紫杉醇、长春新碱等已广泛应用于临床治疗,因此寻找新的活性高、毒副作用小的药物迫在眉睫。近年来,研究人员尝试从天然产物中发现能有效抑制恶性肿瘤的活性成分用于非小细胞肺癌的治疗。
鸦胆子素 A(bruceine A,BA)又称鸦胆子苦素 A,是从植物鸦胆子[(Linnaeus) Merrill]果实中提取分离的苦木内酯类化合物。中医认为,鸦胆子味苦、性寒,有小毒,归大肠、肝经,有清热、解毒、截疟、止痢和腐蚀赘疵之功效[7]。在我国南部地区,鸦胆子多用作抗疟药,临床常将鸦胆子油乳剂用于肺癌及肺癌脑转移、肝癌和消化道肿瘤的辅助治疗。有研究表明鸦胆子油乳剂具有诱导细胞凋亡达到人急性髓性白血病的潜力[8],但关于鸦胆子中的单体成分 BA 是否具有抗肿瘤作用,目前尚无实验数据。本实验拟利用 H460 细胞这一 NSCLS 细胞株,针对 BA 进行体外抗肿瘤作用研究,主要观察其抑制H460 细胞增殖及诱导凋亡作用,同时探讨 BA 作用下 H460 细胞发生凋亡的可能分子机制。
1 材料与方法
1.1 材料
1.1.1 实验试剂 鸦胆子素 A(CAS 25514-31-2,HPLC ≥ 98%)购自南京春秋生物工程有限公司;人非小细胞肺癌细胞株 H460 购自美国 ATCC;胎牛血清(fetal bovine serum,FBS)购自美国 Gibco 公司;RPMI1640 细胞培养基、细胞培养用磷酸盐缓冲液(PBS)、0.25% 胰蛋白酶、10000 U/ml 青霉素 G-链霉素均购自美国 Hyclone 公司;RNase A、四甲基偶氮唑蓝(MTT)购自美国 Amresco 公司;PVDF 膜、ECL 化学发光显色液购自美国 Millipore 公司;Bradford 蛋白定量试剂、5 ×上样缓冲液由普利莱基因技术有限公司提供;Annexin V-FITC/PI 细胞凋亡检测试剂盒购自北京四正柏生物科技有限公司;Hoechst 33342 和 β-actin(A1978)抗体购自美国 Sigma 公司;细胞裂解液、Western blot 所用一抗购自美国 Cell Signaling Technology 公司;辣根过氧化物酶标记的二抗购自北京中杉金桥生物技术有限公司;其他无机试剂均由北京化工厂提供。
1.1.2 仪器 CKX41 倒置显微镜购自日本 Olympus 公司;37081 荧光显微镜购自德国 Carl Zeiss 公司;Mikro 200R 低温离心机购自德国Hettich Zentrifugen公司;Centrifuge 5424 高速离心机购自德国Eppendorf 公司;165-8001 蛋白垂直电泳仪、170-3940 蛋白半干转膜仪、Model 680 酶标仪均购自美国 Bio-Rad 公司;Amersham Imager 600 凝胶成像仪购自美国 GE 公司;FACS Calibur 流式细胞仪购自美国 BD 公司。
1.2 方法
1.2.1 细胞培养 将非小细胞肺癌细胞株 H460 细胞常规复苏后用RPMI1640 细胞培养基培养,在使用前加入 10%(v/v)FBS、100 U/ml 青霉素 G和 100 μg/ml 链霉素,细胞置于 37 ℃和 5% CO2培养箱中培养。
1.2.2 MTT 实验 取对数生长期的细胞,消化计数,以 5 × 103/孔(100 μl)接种于 96 孔板,培养 24 h 后,加入用培养基配置的不同浓度药物 100 μl,每个浓度设置 3 个平行孔,并设不加药物的溶剂对照组和无细胞的空白对照组。加药作用 48 h 后,每孔避光加入 20 μl MTT(5 mg/ml),37 ℃、5% CO2条件下继续培养 4 h,吸净孔内液体,每孔加 150 μl DMSO,在微量振荡器上轻轻振摇 7 min,用酶标仪于 570 nm 波长处测定吸光度值。以不加药物的溶剂对照组细胞活力作为 100%,计算细胞增殖率。
1.2.3 流式细胞仪检测细胞凋亡 取对数生长期的细胞以 3 × 105/孔接种于六孔板中,待细胞贴壁后用药物处理细胞 48 h,同时设置空白对照组。消化收集细胞(包括悬浮细胞和贴壁细胞),用预冷的 PBS 洗涤两次,根据 Annexin V-FITC/PI 染色试剂盒说明书操作染色后,上流式细胞仪检测。实验重复 3 次。
1.2.4 Hoechst 33342 染色 取对数生长期细胞消化计数,以 1 × 105/孔接种于六孔板,待细胞贴壁后分别用 0.5、5、20 μmol/L 浓度的 BA 处理细胞 48 h后,弃去培养基,加入用新鲜培养基配制的2 μg/ml Hoechst 33342 染色液,1 ml/孔。避光染色 15 min,PBS 清洗两次后,加入新鲜培养液,于荧光显微镜观察细胞核内染色质凝集现象并拍摄图片。
1.2.5 免疫荧光染色 六孔板中加入无菌盖玻片,取对数生长期的细胞接种于六孔板中,待细胞贴壁后用 0.5、5、20 μmol/L BA 处理细胞 48 h,制作细胞爬片。将爬好细胞的玻片用 PBS 洗3 次,4% 多聚甲醛固定 15 min,PBS 洗 3 次,0.5% Triton X-100 通透 20 min,吸干表面 PBS,滴加山羊血清室温封闭 1 h,吸掉封闭液直接滴加稀释好的一抗,4 ℃孵育过夜。次日,PBST 浸洗 3 次,吸干多余液体后滴加稀释好的荧光二抗(FITC),避光孵育 1 h,随后 PBST 洗 3 次,吸干残余液体染核,封片。荧光显微镜下观察并采集图像。
1.2.6 Western blot 检测蛋白水平变化 取 H460 细胞以 3 × 105/孔的密度接种于六孔板中,待细胞贴壁后加药物处理细胞 48 h,收集细胞(冰上操作),PBS 清洗,收集细胞沉淀。根据细胞沉淀的体积,加入细胞裂解液。冰上裂解 30 min,12 000 r/min、4 ℃离心 15 min,收集上清即为蛋白提取液,根据 Bradford 法测得的蛋白浓度,按照 20 μg/管分装后上样,根据蛋白分子量大小用 10% 和 15% SDS-PAGE 进行电泳,采用半干转膜法将目的蛋白转移到 PVDF 膜上,用 5% 脱脂奶粉封闭 1 h,漂洗,加一抗 4 ℃孵育过夜,次日取出 PVDF 膜,洗去未结合的抗体,加入二抗溶液,室温下反应 1 h,洗膜,用 ECL 免疫印迹试剂盒进行显色,通过 Amersham Imager 600 凝胶成像系统捕获图像。
1.2.7 RT-PCR 检测基因表达 H460 细胞消化计数后接种于六孔板,待细胞贴壁用 BA 处理,48 h 后,Trizol 裂解提取 RNA,测定浓度后,逆转录为 cDNA,随后进行 RT-PCR,体系为 20 μl。结果以 GAPDH 为内参进行分析。引物序列如下:Bcl2:forward 5' ACGACTTCTCCCGCCGCTACC 3';reverse 5' ACAATCCTCCCCCAGTTCACCC 3';GAPDH:forward 5' GGATGATGTTCTGGAAGAGC C 3';reverse 5' AACAGCCTCAAGA TCATCAGC 3'。
1.3 统计学处理
2 结果
2.1 BA 化学结构及其对 H460 细胞增殖的影响
图 1A 为 BA 的化学结构。不同浓度 BA 处理细胞 48 h 后,与对照组相比,细胞活力明显下降,低浓度即可显著抑制细胞增殖(图 1B)。给予 0.5 μmol/L BA 分别作用 12、24、48、72 h 后,结果显示,随时间增加 BA 显著抑制 H460 细胞增殖,该效应具有时间依赖性(图 1C)。增加 H1299、H1355、A549 三株非小细胞肺癌细胞系,检测 BA 对其增殖影响,结果可见,BA 能不同程度抑制这 4 种细胞增殖,与对照组相比具有显著差异(图 1D)。

图 1 BA 化学结构及其对细胞增殖的影响(A:BA 化学结构;B:不同浓度BA 对H460 细胞增殖影响;C:不同作用时间对H460 细胞增殖影响;D:BA 对不同细胞系增殖影响;与对照组相比,*P < 0.001)
Figure 1 Chemical structure of BA and the proliferation inhibition of cells induced by BA (A: Chemical structure of BA; B: Effects of different concentration of BA for 48 h on proliferation of H460 cells; C: A time-dependent manner with 0.5 μmol/L BA; D: Effect of BA on proliferation of different cell lines, cells were treated with BA for 48 h;*< 0.001 vs. control)
2.2 BA 对细胞凋亡的影响
利用流式细胞术检测 BA 对 H460 细胞凋亡的影响。不同浓度 BA(0.5、5、20 μmol/L)处理 H460 细胞,其凋亡率分别为(31.77 ± 1.81)%、(34.31 ± 1.53)%、(47.45 ± 6.18)%,明显高于对照组(3.16 ± 0.85)%(图 2A)。5 μmol/L BA 作用 H460 细胞 24、48、72 h 后,H460 细胞凋亡率分别为(20.44 ± 3.71)%、(30.29 ± 1.79)%、(47.56 ± 1.31)%,与对照组相比具有显著性差异(图 2B)。提示 BA 可显著诱导 H460 细胞凋亡。
2.3 BA 引起细胞核染色质凝集
Hoechst 33342 进行细胞核染色,经荧光显微镜观察可见,对照组细胞核完整,呈卵圆形,未见凋亡细胞(图 3A)。BA 处理 48 h 后,随药物浓度增加,细胞核出现固缩、碎片化等典型的细胞凋亡形态学改变(如图中箭头处所示),随 BA 浓度升高,凋亡细胞数目增多(图 3B ~ D)。
2.4 BA 对 γ-H2AX 表达的影响
采用免疫荧光检测细胞内 γ-H2AX 表达情况,结果如图 4 所示,不同浓度BA 处理 H460 细胞 48 h 后,细胞内 γ-H2AX 蛋白(图中为绿色荧光)表达增加,且具有剂量依赖性。
2.5 BA 对细胞凋亡相关蛋白表达的影响
Western blot 结果(图 5A)显示,BA 处理H460 细胞 48 h 后,cleaved caspase-9 和 cleaved caspase-3 表达增加,以及 PARP 蛋白切割,提示 BA 诱导 H460 凋亡可能是经由线粒体通路,通过激活 caspase-9 诱导细胞凋亡。随后,针对凋亡发生机制展开进一步研究发现,促凋亡蛋白 Bax 随 BA 浓度的升高表达量增加,抗凋亡蛋白 Bcl-2 表达减少,与对照组相比,Bax/Bcl-2 比值明显升高(图 5B),此外,图 5C 中 Bcl-2 mRNA 表达显著降低(< 0.01),揭示 BA 通过调节 Bax 和 Bcl-2 表达诱导细胞凋亡。

图 2 BA 对 H460 细胞凋亡的影响(A:浓度依赖性;B:时间依赖性;C:凋亡率直方图;与对照组相比,*P < 0.001)
Figure 2 Apoptotic cell death of the H460 cells was induced by BA (A: A dose-dependent manner; B: A time-dependent manner; C: The corresponding histograms of A and B;*< 0.001 vs. control)

图 3 Hoechst 33342 染色观察BA作用H460 细胞48 h 后细胞核的变化(× 100)(A:空白对照组;B:0.5 μmol/L;C:5 μmol/L;D:20 μmol/L;标尺= 100 μm)
Figure 3 The effect of BA on the H460 cells at 48 h was viewed using Hoechst 33342 staining (× 100) (A: Control; B: 0.5 μmol/L; C: 5 μmol/L; D: 20 μmol/L; Bar = 100 μm)
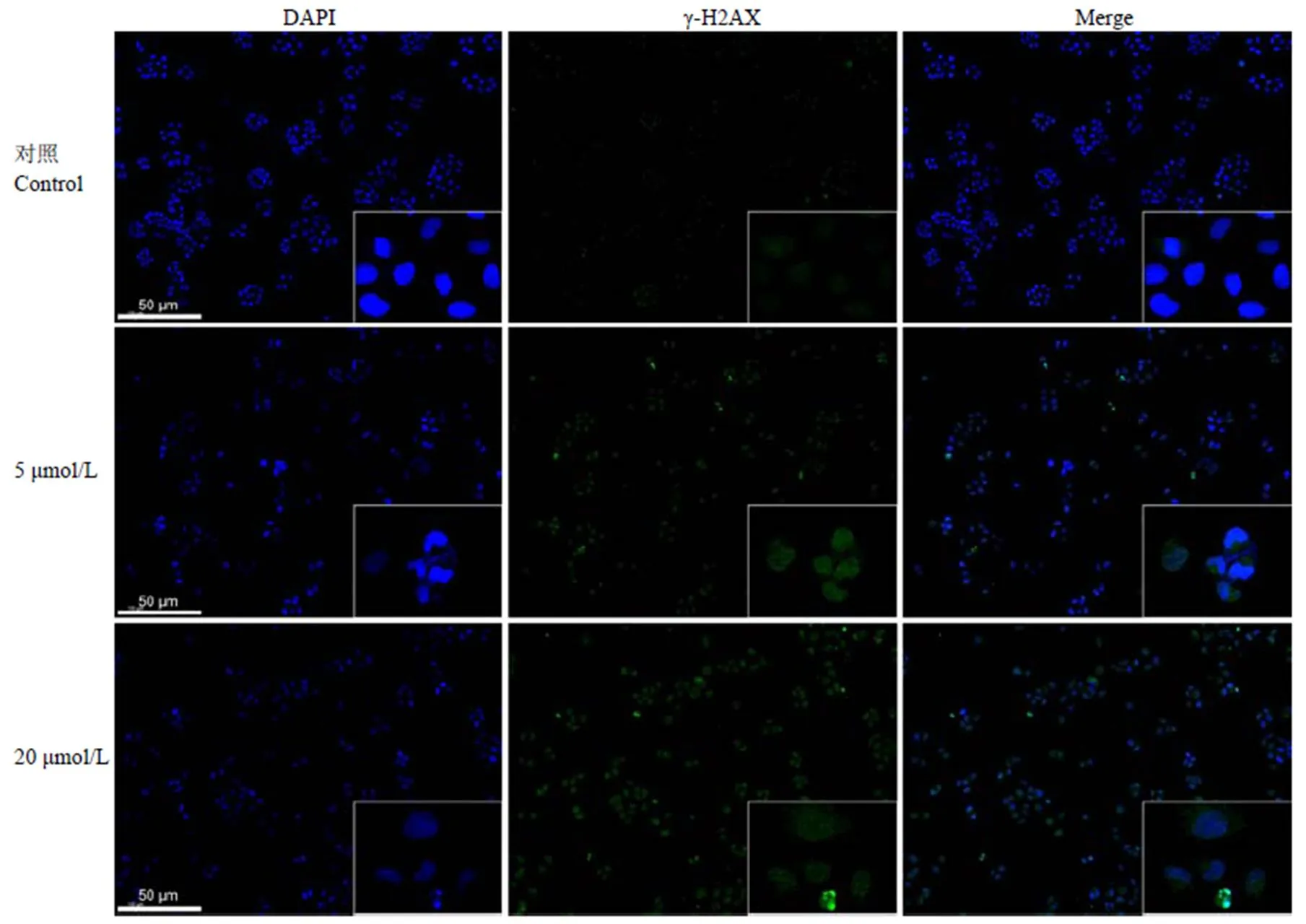
图 4 免疫荧光检测BA 对细胞内γ-H2AX 蛋白含量的影响
Figure 4 The effect of BA on the expression of γ-H2AX in the H460 cells
3 讨论
临床上,鸦胆子油乳剂常用于肺癌、肝癌、消化道癌症的辅助治疗。BA 是鸦胆子果实中提取的单一化合物,目前关于 BA 的研究较少,多局限于提取分离以及治疗犬类疾病[9-10],对于其药理作用尤其是抗癌作用及机制研究方面尚无报道[11-13]。针对单体化合物 BA 诱导 H460 细胞凋亡作用及机制的研究将为非小细胞肺癌的临床治疗提供研究基础。
本实验中,采用不同浓度 BA 处理 H460 细胞 48 h,MTT 结果显示 BA 能显著抑制 H460 细胞增殖(< 0.001)。另选择三株非小细胞肺癌细胞系 H549、H1299、H1355,检测 BA 抑制其增殖情况,结果显示 BA 对不同细胞系均有显著抑制作用,其中对 A549 和 H1355 细胞抑制作用更强。利用 Annexin V-FITC/PI 双染检测 H460 细胞凋亡率,结果显示随药物浓度增加,细胞凋亡率明显升高。Hoechst 33342 染色发现,BA 能引起 H460 细胞染色质凝集,细胞核碎片化。免疫荧光结果同样印证,BA 能诱导 H460 细胞凋亡,表现为 DNA 双链断裂标志蛋白γ-H2AX含量增加。γ-H2AX 属于组蛋白家族成员之一,为磷酸化激活状态,是检测 DNA 损伤的标志蛋白[14],其表达量增加,说明 BA 处理 H460 细胞可能引起细胞 DNA 损伤,但是目前尚不清楚 DNA 损伤是否是 BA 诱导 H460细胞凋亡的机制,有待进一步实验验证。

图 5 BA对H460 细胞凋亡相关蛋白表达的影响(A:BA 对凋亡相关蛋白表达影响;B:Bax/Bcl-2 比值;C:BA 对Bcl-2 mRNA 表达的影响;与对照组相比,**P < 0.01,*P < 0.001)
Figure 5 The effects of BA on apoptosis-related protein expression in H460 cells (A: Expression of apoptosis-related proteins; B: Ratio of Bax/Bcl-2 expression; C: Bcl-2 mRNA expression in the H460 cells;**< 0.01 and*< 0.001 vs. control)
细胞凋亡即细胞的程序性死亡,主要有两条途径调节:第一种是外源性或细胞质途径,由 FAS 死亡受体激活;第二种是内源性或线粒体途径,线粒体内的细胞色素 C 释放,激活 caspase-9 引起级联反应[15]。两种途径最终都汇合到共同的半胱天冬氨酸蛋白酶途径,活化 caspase-3 级联反应,降解细胞内结构和功能蛋白,引起细胞死亡[16]。实验证明 BA 能诱导 H460 细胞凋亡,为检测其作用机制,对凋亡通路蛋白(caspase-9、caspase-3)及其作用底物 PARP 表达量进行 Western blot 检测,结果显示 cleaved caspase-9 和 cleaved caspase-3 表达水平随 BA 浓度增加而升高,提示 BA 诱导 H460 细胞凋亡可能与激活线粒体途径相关。Bcl-2 家族蛋白是线粒体凋亡通路的关键调节因子,主要分为两类,促凋亡蛋白:Bax、Bak、Bad、Bcl-Xs等和抑制凋亡蛋白:Bcl-2、Bcl-XL、Bcl-W 等[17]。两者通过促进或抑制细胞色素 C 的释放发挥调节作用[18]。Western blot 检测 Bax 和 Bcl-2 表达水平发现,随给药浓度增加,Bax 表达升高,Bcl-2 表达降低,提示 BA 通过调节 Bax/Bcl-2 比值诱导H460 凋亡。此外,RT-PCR 结果提示,BA 能抑制 Bcl-2 mRNA 表达,其结果与 Western blot 结果一致。
综上所述,BA 能在体外诱导 H460 细胞凋亡,抑制其增殖,其作用机制主要是通过升高Bax/Bcl-2 比值,激活线粒体通路中的 caspase-9,活化下游caspase-3,最终引起细胞凋亡。此外 BA 诱导 H460 细胞发生 DNA 损伤也可能为其潜在的抗肿瘤机制之一。BA 作为一种天然的单体化合物,其对 H460 细胞的作用及作用机制仍需深入研究。本实验只是验证了其对 H460 细胞体外作用情况,后续实验会进一步扩大 BA 抑瘤谱,针对 EGFR 突变细胞系和对 EGFR 具有抗药性细胞系进行进一步研究,并针对 Bcl-2 家族调节蛋白上游通路进行检测,同时开展体内动物实验,为 BA 应用于非小细胞肺癌的临床治疗提供参考依据。
[1] Oser MG, Niederst MJ, Sequist LV, et al. Transformation from non-small-cell lung cancer to small-cell lung cancer: molecular drivers and cells of origin. Lancet Oncol, 2015, 16(4):e165-e172.
[2] Inoue A, Yoshida K, Morita S, et al. Characteristics and overall survival of EGFR mutation-positive non-small cell lung cancer treated with EGFR tyrosine kinase inhibitors: a retrospective analysis for 1660 Japanese patients. Jpn J Clin Oncol, 2016, 46(5):462-467.
[3] Reck M, Rabe KF. Precision diagnosis and treatment for advanced non-small-cell lung cancer. N Engl J Med, 2017, 377(9):849-861.
[4] Kumarakulasinghe NB, van Zanwijk N, Soo RA. Molecular targeted therapy in the treatment of advanced stage non-small cell lung cancer (NSCLC). Respirology, 2015, 20(3):370-378.
[5] Kumar SS, Higgins KA, McGarry RC. Emerging therapies for stage III non-small cell lung cancer: stereotactic body radiation therapy and immunotherapy. Front Oncol, 2017, 7:197.
[6] Molina JR, Yang P, Cassivi SD, et al. Non-small cell lung cancer: epidemiology, risk factors, treatment, and survivorship. Mayo Clin Proc, 2008, 83(5):584-594.
[7] Bawm S, Matsuura H, Elkhateeb A, et al. In vitro antitrypanosomal activities of quassinoid compounds from the fruits of a medicinal plant, Brucea javanica. Vet Parasitol, 2008, 158(4):288-294.
[8] Zhang H, Yang JY, Zhou F, et al. Seed oil of Brucea, javanica induces apoptotic death of acute myeloid leukemia cells via both the death receptors and the mitochondrial-related pathways. Evid Based Complement Alternat Med, 2011, 2011:965016.
[9] Sun Z, Cao Y, Zhai LZ. Java brucea and Chinese herbal medicine for the treatment of cholesterol granuloma in the suprasellar and sellar regions: A case report and literature review. Medicine (Baltimore), 2017, 96(5):e5930.
[10] Pan L, Chin YW, Chai HB, et al. Bioactivity-guided isolation of cytotoxic constituents of Brucea javanica collected in Vietnam. Bioorg Med Chem, 2009, 17(6):2219-2224.
[11] Yan Z, Guo GF, Zhang B. Research of Brucea javanica against cancer. Chin J Integr Med, 2017, 23(2):153-160.
[12] Kim SH, Liu CY, Fan PW, et al. The aqueous extract of Brucea javanica suppresses cell growth and alleviates tumorigenesis of human lung cancer cells by targeting mutated epidermal growth factor receptor. Drug Des Devel Ther, 2016, 10:3599-3609.
[13] Huang Z, Yang G, Shen T, et al. Dehydrobruceine B enhances the cisplatin-induced cytotoxicity through regulation of the mitochondrial apoptotic pathway in lung cancer A549 cells. Biomed Pharmacother, 2017, 89:623-631.
[14] Zhang F, Zhang T, Qu Y, et al. Replication-dependent γ-H2AX formation is involved in docetaxel-induced apoptosis in NSCLC A549 cells. Oncol Rep, 2010, 24(5):1297-1305.
[15] Ghobrial IM, Witzig TE, Adjei AA. Targeting apoptosis pathways in cancer therapy. CA Cancer J Clin, 2005, 55(3):178-194.
[16] Reed JC. Bcl-2 and the regulation of programmed cell death. J Cell Biol, 1994, 124(1-2):1-6.
[17] Takeda K, Stagg J, Yagita H, et al. Targeting death-inducing receptors in cancer therapy. Oncogene, 2007, 26 (25):3745-3757.
[18] Mahdavi S, Khodarahmi P, Roodbari NH. Effects of cadmium on Bcl-2/ Bax expression ratio in rat cortex brain and hippocampus. Hum Exp Toxicol, 2018, 37(3):321-328.
Bruceine A induces apoptosis in non-small-cell lung cancer cell line H460 by regulating mitochondria-related apoptosis signaling
LI Cong, WANG Yan, ZHAO Xiao-li, WANG Zhen, JIANG Jian-dong
Institute of Materia Medica (LI Cong, WANG Yan, JIANG Jian-dong), Institute of Medicinal Biotechnology (ZHAO Xiao-li, WANG Zhen), Chinese Academy of Medical Sciences & Chinese Academy of Medical Sciences, Beijing 100050, China
We aim to investigate whether bruceine A (BA) induces apoptosis in human non-small-cell lung cancer cell (NSCLC) H460 cells and to learn the mechanism.
The proliferation of NSCLC cell lines, including H460, A549, H1299 and H1355, was detected by the MTT assay after treatment with BA. Effect of BA on apoptosis in the H460 cells was measured by FACS analysis with Annexin V-FITC/PI staining. Hoechst 33342 and immunofluorescence (IF) staining were used to observe the chromatin condensation and the expression of the DNA damage marker protein γ-H2AX, respectively. Western blot and real-time PCR were used to detect the expression of apoptosis-related protein and mRNA in the H460 cells, respectively.
Based on MTT result, the proliferation was significantly inhibited in H460 cells after treatment with BA for 48 h. The proliferation inhibition of other NSCLC cell lines A549, H1299 and H1355 upon BA treatment was detected, and BA inhibited the proliferation of the three cell lines in a dose-dependent manner. Significant induction of apoptosis was observed in a time-dependent fashion upon BA treatment in the H460 cells. Also, remarkable nuclear condensation was observed following BA treatment in H460 cells using Hoechst 33342 staining. DNA damage marker γ-H2AX was increased by BA as evidenced by IF staining. Furthermore, BA increased and decreased the expression of Bax and Bcl-2, respectively, and cleaved caspase-9, caspase-3 and PARP protein, as demonstrated by Western blot. Also, Bcl-2 gene expression in the H460 cells was significantly decreased after BA treatment.
BA inhibits the H460 cells proliferation by inducing apoptosis, which may be mediated through the mitochondria-dependent apoptotic signaling pathways.
BRUCEINE; Apoptosis; Carcinoma, non-small-cell lung; H460 cell
JIANG Jian-dong, Email: jiang.jdong@163.com; WANG Zhen, Email: wangzhen@imb.pumc.edu.cn
10.3969/j.issn.1673-713X.2019.04.002
国家自然科学基金(500101206)
蒋建东,Email:jiang.jdong@163.com;王真,Email:wangzhen @imb.pumc.edu.cn
2019-03-06
猜你喜欢
西北民族大学学报(自然科学版)(2022年2期)2022-07-06
中国临床医学影像杂志(2022年2期)2022-05-25
中国药学药品知识仓库(2022年1期)2022-03-23
科学与生活(2021年16期)2021-11-25
昆明医科大学学报(2021年4期)2021-07-23
纺织科技进展(2021年4期)2021-07-22
兰州理工大学学报(2021年3期)2021-07-05
中国市场(2017年5期)2017-03-15
医学研究杂志(2015年9期)2015-07-01
医学研究杂志(2015年12期)2015-06-10
