
黄绿绿僵菌侵染褐飞虱的电镜观察
2019-08-29 07:17张珏锋陈建明钟海英
浙江农业学报 2019年8期
张珏锋,陈建明,李 芳,钟海英,吴 鸿
(1.中国林业科学院 亚热带林业研究所,浙江 杭州 311400; 2.浙江省农业科学院 植物保护与微生物研究所,浙江.杭州 310021)
褐飞虱[Nilaparvatalugens(Stål)]是我国及东南亚水稻产区的主要害虫之一,属“r对策型”、迁飞性害虫,多年来给我国水稻生产造成极大危害[1]。由于化学杀虫剂在田间被多用、滥用,近年来褐飞虱的抗药性日益增强,而且褐飞虱在对抗性水稻的为害过程中,容易产生新的致害性种群[2]。新的致害性种群的出现,使得大面积推广种植的抗性品种水稻,如IR系列、Mudgo、ASD7等失去褐飞虱抗性。多因素累加,致使当下褐飞虱的田间防控困难重重。
绿僵菌(Metarhiziumspp.)是一种广泛应用于农林害虫防治的无性真菌[3-4],具有强大的粘附寄主的功能,能够以独特的方式侵入昆虫体壁,即使在坚硬的表面如玻璃等上都可产生附着胞。绿僵菌的附着胞具有穿刺作用的结构,它可以沿气孔开口处或直接刺透寄主表皮进入寄主内部[5-7],也可通过消化道、伤口等途径侵入虫体[8-9],使寄主感染微生物而死,对刺吸式害虫的防治具有积极效果。同时,绿僵菌还具有田间再侵染的能力,为褐飞虱的生物防治提供了可能,有望成为化学杀虫剂的良好替代物。但绿僵菌也具有起效时间长、对环境温湿度要求高等缺陷,田间防效往往远不及实验室效果,如从自然界感染绿僵菌的棕榈象(Rhynchophorusferrugineus)上分离获得的菌株仅在实验室条件下有效[10]。相对于活动的虫态,活体真菌对静止虫态作用时间更长,作用成功概率也更大。因此,明确绿僵菌是否能够通过侵染褐飞虱卵巢实现继代传播,以及是否能够侵染稻株内卵块从而致使卵块无法孵化,对褐飞虱的生物防治具有应用价值。有关绿僵菌对寄主昆虫体壁侵染的过程,已在小菜蛾(Plutellaxylostalla)[11-12]、棕榈象[13]、榛实象甲(Curculionucum)[14]、褐飞虱[15]上有过报道,但关于绿僵菌侵染寄主卵巢、卵块的相关报道较少。
本试验应用扫描电镜结合半薄切片观察研究了黄绿绿僵菌(Metarhiziumflavoviride)对褐飞虱从体表侵染至体内定殖、萌发的过程,以及入侵寄主卵巢的现象,可直观地表现黄绿绿僵菌对褐飞虱的侵染能力和侵入方式,旨在为评价该菌的杀虫作用和应用前景提供基础资料和参考。
1 材料与方法
1.1 供试材料
供试菌株为中国林业科学院林业研究所提供的黄绿绿僵菌。
供试褐飞虱在浙江省农业科学院植物保护与微生物研究所人工气候培养室内繁殖10代以上,选取发育状况一致的褐飞虱成虫作为接菌对象。
1.2 黄绿绿僵菌孢子悬浮液的配制
将供试菌株转接到PDA平板培养基上,(25±2)℃、无光条件下培养10 d,待真菌充分产孢后,用灭菌的接种环轻轻将该菌株的分生孢子粉刮到盛有5 mL 0.05%(质量分数)Tween-80的三角瓶中,充分振荡30 min后按比例稀释,血球计数板计数,配制成1.0×108mL-1的孢子悬浮液。
1.3 褐飞虱接菌方法
取褐飞虱成虫10头,用手握式喷雾器均匀喷施1.0×108mL-1的孢子悬浮液后接入45日龄TN1水稻苗,外罩玻璃罩,以喷施0.05%(质量分数)Tween-80的褐飞虱为对照。处理后试虫置于(28±2)℃、相对湿度85%和光/暗周期14 h/10 h的恒温培养箱中饲养,72 h后取样:部分解剖取卵巢,并做冷冻切片,观察卵巢侵染状态;部分用于制作半薄切片,观察体壁侵染情况。持续观察处理过的褐飞虱,其间如有死亡褐飞虱,取样用于扫描电镜观察。
1.4 水稻植株接菌方法
取45日龄稻株洗净,剪除叶片后将茎秆剪成5~7 cm小段,ddH2O冲洗3次,晾干后用小号缝衣针轻扎,模仿褐飞虱取食,用手握式喷雾器在模仿取食处均匀喷施1.0×108mL-1的孢子悬浮液。将处理后的水稻茎秆放入铺有3层湿润滤纸的培养皿中,置于28 ℃培养箱内,以喷施0.05%(质量分数)Tween-80的茎秆为对照,72 h后取样、切片,用于电镜观察。
1.5 褐飞虱卵块接菌方法
取初孵褐飞虱雌雄成虫各1头,接入置有45日龄TN1稻株的盆钵中产卵,外罩玻璃罩,48 h后移去褐飞虱雌雄成虫,于茎秆部喷施1.0×108mL-1的孢子悬浮液10 mL,以喷施0.05%(质量分数)Tween-80的稻株为对照。将带卵稻株置于(28±2)℃、相对湿度85%和光/暗周期14 h/10 h的恒温培养箱中培养,72 h后解剖稻株取卵块用于扫描电镜观察。取对照稻株中卵块10枚,喷施黄绿绿僵菌后置于灭菌水琼脂平板上,以喷施0.05%(质量分数)Tween-80的卵块为对照,(28±2)℃培养箱中培养72 h后用于扫描电镜观察。
1.6 绿僵菌对褐飞虱卵块孵化率及幼虫、成虫历期的影响
取初孵褐飞虱雌雄成虫各1头,喷施1.0×108mL-1的孢子悬浮液,作为处理(T),以喷施0.05%(质量分数)Tween-80作为对照(CK)。处理后的试虫接入45日龄TN1水稻苗,放入(25±2)℃、相对湿度85%和光/暗周期14 h/10 h的恒温培养箱中产卵,每处理10个重复,每24 h换置一批稻苗。72 h后移去褐飞虱,后期检查卵块孵化率及各虫态发育历期。
1.7 扫描电镜处理方法
所取样品用3%(体积分数)戊二醛隔夜固定12 h后,用锇酸固定12 h,磷酸缓冲液浸洗3次,每次30 min,然后依次用体积分数分别为50%、70%、80%、90%、100%的乙醇逐级梯度脱水,每级12 h,其中用100%(体积分数)乙醇脱水2次[16]。醋酸异戊酯置换2次,每次停留30 min。样品置于室温下自然干燥后,用导电胶将样品粘于扫描电镜样品台上,然后经离子溅射法喷金,在TM-1000 HICHMI型扫描电子显微镜下进行观察并拍照。
1.8 半薄切片处理方法
取褐飞虱样品置于卡诺固定液中固定12 h后,依次于体积分数分别为50%、70%、95%的乙醇中逐级脱水,每级脱水3 h,脱水后材料按塑料包埋剂(Technovit7100)的说明书中方法进行包埋,之后将样品置于预渗透液(等体积的体积分数为95%的乙醇与base liquid Technovit7100混合)中,12 h后转入渗透液(1 g硬化剂1溶于100 mL base liquid Technovit7100),渗透6~12 h后在小包埋孔中摆好,后置于新鲜包埋液(硬化剂1与base liquid Technovit7100的质量体积比为1∶15)中,聚合2 h以后,于Leica Ultracut R型超薄切片机切片,厚度1 μm,切好的片子用小铜网捞片,直接转到滴有水的干净载玻片上,电热台展开、烘干后,用加拿大树胶封片。
2 结果与分析
2.1 黄绿绿僵菌分生孢子在褐飞虱体表的侵染过程
由扫描电镜观察结果可知:黄绿绿僵菌侵染褐飞虱时,在寄主褐飞虱体表不同结构区的入侵行为存在一定差异。黄绿绿僵菌最先从寄主腹部侵染(图1-A),推测可能是由于褐飞虱腹部褶皱处较多,易于菌体入侵。菌体在褐飞虱腹部定殖后慢慢延伸至寄主背部褶皱凹陷处(图1-B),再逐步侵染至背部体壁光滑处,以菌丝体形式包围寄主虫体后继续延伸侵染至寄主褐飞虱的翅(图1-C、D)、复眼(图1-E)、触角(图1-F)等部位。黄绿绿僵菌侵染褐飞虱以后,在寄主表皮以孢子(图1-G)和菌丝体(图1-H)2种形态存在。已有的研究表明,绿僵菌以附着孢和菌丝体的方式侵入寄主虫体,侵染过程中附着胞的产生受寄主表皮营养条件、pH值等的影响。在本试验中我们也观察到,黄绿绿僵菌分生孢子附着于寄主体表后,可利用体表的营养物质萌发产生芽管,形成附着胞入侵(图1-C),当未找到侵入点时,则形成菌丝在寄主体表横向生长(图1-D),寻找体表较薄弱部位侵入。
2.2 黄绿绿僵菌分生孢子对稻株的侵染观察
观察黄绿绿僵菌处理的模拟褐飞虱取食水稻茎秆部切片可见:横切面的表皮、基本组织和维管束完整,排列整齐(图2-A),与对照处理(图2-B)相比无明显差异,未发现有明显外力破坏痕迹。同时,镜检也未在稻茎体腔内发现有类似于黄绿绿僵菌孢子或菌丝体的物质。
如图3所示,黄绿绿僵菌溶液处理稻株中解剖获得的卵块(图3-A)与对照稻株内解剖获得的卵块(图3-B),在电镜观察下两者外观形态相似,体表光滑,无明显菌丝黏附,但在处理稻株内获得的卵块体表可观察到少量的黄绿绿僵菌孢子(图3-C)。黄绿绿僵菌处理的离体褐飞虱卵块72 h后即被大量菌丝体包被(图3-E),且卵块表面可观察到黄绿绿僵菌菌丝体及分生孢子(图3-F)。由以上结果推测:黄绿绿僵菌并不能在水稻植株内定殖,但可定殖于离体的褐飞虱卵块,因而喷施于稻株表面的黄绿绿僵菌在稻株表面存在产卵痕时有可能通过产卵痕侵染产于稻株叶鞘组织内的褐飞虱卵块。

A,绿僵菌对褐飞虱表皮的侵染(腹部);B,绿僵菌对褐飞虱表皮的侵染(背部);C,褐飞虱翅上的黄绿绿僵菌孢子;D,褐飞虱翅上的黄绿绿僵菌菌丝体;E,褐飞虱复眼处孢子;F,褐飞虱触角处孢子;G,形成穿透板;H,体表布满菌丝体。A, Invasion behavior of M. flavoviride on the cuticle of N. lugens(Abdomen); B, Invasion behavior of M. flavoviride on the cuticle of N. lugens(Back); C, M. flavoviride conidia on the wing of N. lugens; D, Mycelia of M. flavoviride on the wing of N. lugens; E, Conidia on compound eye; F, Conidia on antenna; G, Penetrating plate; H, Extensive growth of hyphae.图1 寄主体表不同结构区黄绿绿僵菌的入侵行为Fig.1 Invasion behavior of M. flavoviride on different surface topography of host
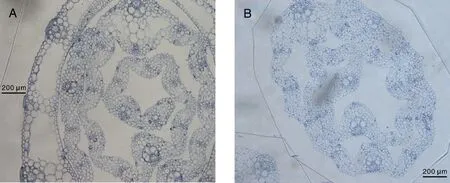
图2 试验处理(A)和对照(B)稻株茎秆的显微观察Fig.2 Observation of semithin section of stem of Oryza sativa under both treatment (A) and control (B)
2.3 黄绿绿僵菌分生孢子对褐飞虱体壁的侵染
电镜下观察黄绿绿僵菌侵染72 h的褐飞虱表皮组织可见,相对于健康体壁细胞(图4-A),被侵染的褐飞虱表皮组织内大量菌丝穿透形成大小、形状各异的菌丝段和菌丝取代表皮组织,且组织内细胞器因菌丝穿透遭到破坏,细胞排列松弛,部分变成空泡(图4-B、C)。
解剖死亡24 h之内的褐飞虱,虫体腹部呈硬块状(图5-A),电镜下可见孢子堆积(图5-B)。推测黄绿绿僵菌通过体表薄弱处侵入寄主体内后,借助寄主的营养快速繁殖,导致菌体大量堆积在褐飞虱腹部,引起寄主死亡。
2.4 黄绿绿僵菌分生孢子在褐飞虱卵巢的侵染过程
对黄绿绿僵菌侵染的褐飞虱卵巢进行镜检可见,内部分布着大量形似黄绿绿僵菌孢子的物质(图6-A、B),但由于褐飞虱体内存在可垂直卵传的共生类酵母菌,因而仅通过镜检无法判断图6中黄绿绿僵菌是否可以侵染寄主卵巢。

A,处理稻株内卵块;B,对照稻株内卵块;C,处理稻株内卵块(箭头1、2所指为黄绿绿僵菌孢子);D,对照处理离体卵块;E,黄绿绿僵菌处理离体卵块;F,卵块表面分布的黄绿绿僵菌菌丝体。A, Eggs in M. flavoviride treated rice plant; B, Eggs in control rice plant; C, Eggs in M. flavoviride treated rice plant (Arrow 1, 2 referred to M. flavoriride spores); D, Isolated egg under control; E, Isolated egg treated by M. flavoviride; F, Mycelium on the surface of the isolated egg.图3 不同处理褐飞虱卵块的电镜观察Fig.3 Scanning electron microscope observation of N. lugens eggs under different treatments

A,对照处理;B,黄绿绿僵菌分解的体壁;C,体壁内分布的孢子。A, Control cuticle; B, Cuticle decomposed by M. flavoviride; C, Conidia distributed in cuticle.图4 褐飞虱体壁被黄绿绿僵菌侵染前后对比Fig.4 Comparison of the cuticle of N. lugens before and after infection of M. flavoviride

A,腹部;B,堆积的孢子。A, Abdomen; B, Accumulated spores.图5 死亡褐飞虱腹部电镜观察Fig.5 Electron microscopic observation of dead N. lugens
2.5 黄绿绿僵菌对褐飞虱卵块孵化率及幼虫、成虫历期的影响
由图7可知,处理褐飞虱种群的卵块孵化率集中在61.10%~72.67%,黄绿绿僵菌处理时间的延长并未降低寄主的卵块孵化率,且与对照褐飞虱种群的孵化率相比也无显著差异。表1的数据表明,不同处理的褐飞虱若虫历期和成虫历期并无显著差异,说明黄绿绿僵菌处理并未对褐飞虱若虫与成虫历期造成影响。由上述结果推测,短期(24~72 h)黄绿绿僵菌处理还不足以对寄主体内的卵块产生影响。

箭头所指为黄绿绿僵菌孢子。The arrows in the picture referred to M. flavoviride spores.图6 黄绿绿僵菌侵染后的褐飞虱卵巢半薄切片观察Fig.6 Observation of semithin section of N. lugens ovary after M. flavoviride infection
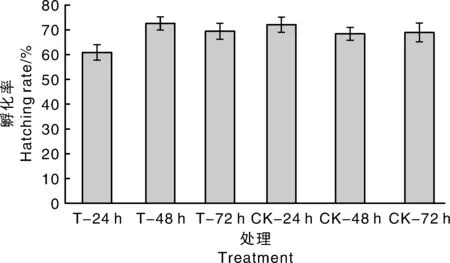
图7 不同处理的褐飞虱卵块孵化率Fig.7 Hatching rate of N. lugens under different treatments
表1 不同处理的褐飞虱若虫历期和成虫历期
Table 1 Adult and nymph duration ofN.lugensafter different treatments
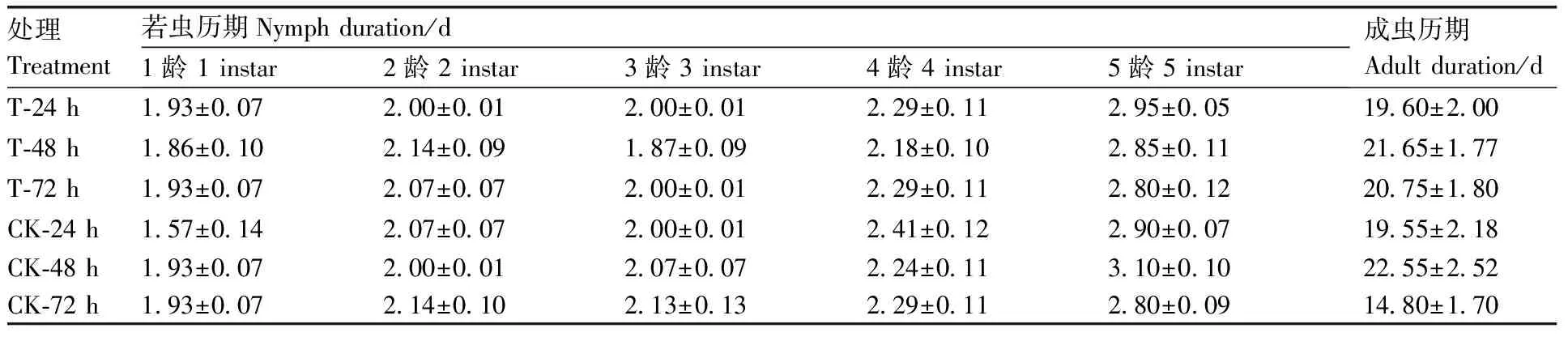
处理Treatment若虫历期Nymph duration/d1龄 1 instar2龄 2 instar3龄 3 instar4龄 4 instar5龄 5 instar成虫历期Adult duration/dT-24 h1.93±0.072.00±0.012.00±0.012.29±0.112.95±0.0519.60±2.00T-48 h1.86±0.102.14±0.091.87±0.092.18±0.102.85±0.1121.65±1.77T-72 h1.93±0.072.07±0.072.00±0.012.29±0.112.80±0.1220.75±1.80CK-24 h1.57±0.142.07±0.072.00±0.012.41±0.122.90±0.0719.55±2.18CK-48 h1.93±0.072.00±0.012.07±0.072.24±0.113.10±0.1022.55±2.52CK-72 h1.93±0.072.14±0.102.13±0.132.29±0.112.80±0.0914.80±1.70
3 讨论
绿僵菌与传统的化学杀虫剂相比,具有环境友好、人畜无害、效果持续时间长等优点,但真菌杀虫剂的有效成分均为活体细胞,其田间防效及稳定性受环境因子,如温湿度[17]、紫外线[18-19]、化学药剂[20]等的影响,使得生防真菌的应用受到作物生长环境和季节的限制。生防真菌具有独特的体壁侵染方式,以寄主虫体体壁作为侵染的第一道屏障。这在生防真菌的抗逆及害虫侵染过程中发挥着重要作用。
本试验所用的黄绿绿僵菌菌株在实验室条件下能识别褐飞虱表皮,破坏表皮组织,使表皮层疏松以利菌丝体穿透,从而形成大小、形状各异的菌丝段和菌丝取代表皮组织,成功附着建立寄生关系[21-22]。菌丝体同时侵入寄主各器官组织,破坏寄主组织与器官,直至整个虫体充满菌丝和菌丝段,导致寄主褐飞虱死亡。
绿僵菌的生长周期较长,易受田间环境因子的影响,且在菌丝体侵入破坏寄主组织与器官之前并不影响寄主的取食、产卵等行为,虽可侵染褐飞虱,但田间防治效果并不明显。因此,明确绿僵菌是否能够侵染褐飞虱卵巢致其无法正常产卵,以及是否能够感染稻株内卵块导致其无法孵化,对褐飞虱的田间防治更具应用价值。这是因为相对于活动的虫态,活体真菌对静止虫态的作用时间更长,作用成功概率也更大。目前,关于真菌侵染寄主卵块的相关报道较少,Toledo等[23]曾在蜡蝉(Peregrinusmaidis)雌成虫被白僵菌(Beauveriabassiana)和绿僵菌(Metarhiziumanisopliae)侵染48 h后的卵巢管和卵母细胞中发现菌丝体,说明48 h之后绿僵菌即可侵染蜡蝉的生殖系统。本试验中,黄绿绿僵菌处理72 h的褐飞虱卵巢切片可见大量形似黄绿绿僵菌孢子的物质,但由于褐飞虱自身携带的类酵母共生菌可垂直卵传,且形态与黄绿绿僵菌相似,因此单凭镜检无法判断绿僵菌是否可以侵染褐飞虱卵巢,且黄绿绿僵菌处理72 h之后的褐飞虱所产卵块孵化率,以及成虫、若虫历期都与对照种群无显著差异。此外,试验结果表明,黄绿绿僵菌菌体可定殖于褐飞虱卵块体表。结合以上试验结果推测,黄绿绿僵菌可以侵染褐飞虱生殖系统并定殖于卵块,但72 h还不足以使其菌丝体侵入寄主褐飞虱的生殖系统并对其生殖行为产生影响。这与Toledo等[23]的试验结果存在差异,可能是由于蜡蝉与褐飞虱的表皮细胞壁结构、组成成分等存在差异,导致黄绿绿僵菌在寄主褐飞虱的体表定殖、产生附着胞、侵染所需时间较长。绿僵菌对褐飞虱卵块的侵染对于褐飞虱的田间防控具有重要意义。李茂业等[15]曾报道绿僵菌可通过稻株产卵痕侵染褐飞虱卵块。本试验也发现,喷施于稻株表面的黄绿绿僵菌在稻株表面存在产卵痕时有可能通过产卵痕侵染产于稻株叶鞘组织内的褐飞虱卵块,但观察到的卵块体表孢子数量较少,推测并不能对卵块存活率产生影响,此外,也未观察到卵块变色死亡的现象。绿僵菌的稳定性易受环境因子影响,且水稻不同生长期植株的含水量、营养成分、表皮结构等都存在差异,推测以上原因共同造成了绿僵菌对稻株侵染的不稳定性。
本研究结果表明,黄绿绿僵菌可侵染褐飞虱及其卵块,但侵染所需时间较长,且易受外界因子影响。因此,黄绿绿僵菌对褐飞虱的田间防控,在使用上需结合田间气候变化趋势、褐飞虱虫龄等因素综合考虑,适时施用。同时,进一步提高黄绿绿僵菌的侵染效率,加强黄绿绿僵菌侵染能力方面的研究,对于提高黄绿绿僵菌对褐飞虱的田间防效具有现实意义。
猜你喜欢
华南农业大学学报(2022年5期)2022-08-02
湖北农业科学(2021年22期)2021-12-13
特种经济动植物(2021年12期)2021-12-13
ELLE世界时装之苑(2021年4期)2021-04-07
湖北农业科学(2020年17期)2020-11-09
农家科技下旬刊(2020年5期)2020-06-19
农业机械学报(2020年2期)2020-03-09
椰城(2018年9期)2018-11-15
农家顾问(2016年5期)2016-05-14
小溪流(画刊)(2015年9期)2016-03-03
