
茂兰喀斯特森林退化区凋落物的分解动态
2019-09-10 07:22王敏容丽俞国松李芹
广西植物 2019年8期
关键词:喀斯特
王敏 容丽 俞国松 李芹


摘要: 為了解亚热带气候型的茂兰喀斯特森林退化区次生林和灌木林的凋落物分解动态过程,该研究采用分解袋法,对茂兰喀斯特森林退化区不同类型的凋落物在不同坡位的分解状况进行了为期18个月的观测,并通过分析凋落物分解时的失重量和失重率的动态变化,比较了次生林和灌木林的凋落叶的失重率变化,探讨了不同坡位对凋落物分解的影响。结果表明:各种类型凋落物的分解速率和失重率在退化区内存在明显的差异, 落叶>常绿叶>枯枝(P<0.05),三种凋落物整体变化趋势在分解过程中大致相同, 它们在早期都快速分解,中期分解变慢,后期开始加速;落叶在次生林与灌木林中的前期分解速率基本同步,后期为灌木林落叶>次生林落叶,而常绿叶在灌木林与次生林中的分解速率则表现为基本同步;利用回归方程对凋落叶分解50%和95%所需时间进行估测,得出落叶和常绿叶在灌木林中分解50%和95%所需时间少于次生林的;在不同坡位,三种凋落物分解速率的总体趋势为中坡>上坡;三种凋落物的C含量波动性较大,但总体变化趋势是随分解时间的增加而减少,随着分解时间增加,N含量增加,而C/N比则降低。
关键词: 茂兰, 喀斯特, 凋落物, 分解动态, 养分释放
中图分类号: Q948文献标识码: A文章编号: 1000-3142(2019)08-1081-11
Abstract: In order to study the dynamic process of litter decomposition of secondary forest and shrubbery in Maolan karst forest degradation area of subtropical climate type, the decomposition bag method was used to observe the litter decomposition of different types in different slopes in Maolan karst forest degradation area for 18 months. By analyzing the dynamic changes of weight loss and weight loss rate during the decomposition of litters, comparing the changes of weight loss rate of litters in secondary forest and shrubbery and the effects of different slope positions on litters decomposition. The results showed that there were significant differences in the decomposition rate and weight loss ratio of all types of litters in the degradation zone, fallen leaves>often leaves>deadwood(P<0.05), the overall change trend of the three litters was roughly the same in the decomposition process, they all decomposed rapidly in the early stage, slowed down in the middle stage and accelerated in the later stage. The decomposition rate of leaves in secondary forest and shrubbery was basically synchronized at the earlier stage, deciduous shrub>secondary forest leaves in the later, while the decomposition rate of the common green leaves in secondary forest and shrubbery was basically synchronous. The regression equation was used to estimate the time required for the decomposition of litter leaves by 50% and 95%, and it was concluded that the time required for the decomposition of deciduous leaves and evergreen leaves in shrubbery by 50% and 95% was less than that of secondary forests. At different slope positions, the general trend of decomposition rate of three kinds of litters is mesoslope>upslope. The C content of the three litters fluctuates greatly, but the overall change trend is to decrease with the increase of decomposition time, with the increase of decomposition time, the N content increases, while the C/N ratio decreases.
Key words: Maolan, karst forest, litter, decomposition of dynamic, nutrient release
森林凋落物是指生态系统内产生的生物成分返回森林表面,为分解者提供了物质和能量来源,用于维持生态系统功能的所有有机物质的总称(王凤友,1989)。凋落物分解是陆地生态系统物质循环和能量流动的重要组成部分,也是维持生态系统功能的主要过程之一(Swift et al., 1979; Berg & McClaugherty, 1989; Sun et al., 2004; 张浩和庄雪影,2008)。对森林凋落物的研究可追溯到19世纪,最早研究森林凋落物的是德国E.Ebermayer(1876),他对森林凋落物的生产和化学组成进行了研究。随后, 国外许多学者如Berg et al.(1993)、Edmonds & Thomas(1995)、Singh et al.(1999)等关于全球森林凋落物分解和养分释放进行了大量报道。唐仕姗等(2014)研究了中国森林生态系统凋落物分解速率的控制因子和分布特征。
我国西南地区喀斯特生态环境极其脆弱,贵州喀斯特森林生态系统属于亚热带森林生态系统,其生境与相同气候条件下常态地貌植被生境相比,有明显不同,主要表现为岩石裸露率高、土层浅薄、土被不连续、土壤富钙、偏碱性等(杨安学和彭云,2007)。朱守谦等(1995)研究了贵州喀斯特森林生态系统生境特征,得出喀斯特森林生态系统具有多层空间,浅薄的土壤层、土被不连续、地表贮水能力低以及岩石渗漏性强等;这样的生境条件下发育的植被需要更多的养分才能维持其生长,因而研究喀斯特退化区的凋落物分解对喀斯特森林生态系统的养分循环和能量转换具有重要意义。近年来,已有不少学者对喀斯特森林凋落物进行了研究,王庆玲(2009)对黔中喀斯特地区次生林凋落物生态功能做了研究,钱正敏等(2009)研究了茂兰喀斯特森林凋落物动态,朱双燕等(2009)和卢晓强等(2014)先后分析了喀斯特区原生林和次生林凋落物养分归还的特征,曾昭霞等(2011)研究了桂西北喀斯特区原生林与次生林凋落物量及其降解,俞国松等(2011)和熊红福等(2013)先后对喀斯特地区不同演替阶段植物群落的凋落物量、组成特征及季节动态变化进行了研究,陈元等(2017)研究了黔西北喀斯特山区植物群落凋落物动态,赵畅等(2018)研究了茂兰喀斯特原生林不同坡向及分解层的凋落物现存量和养分特征。但是,对喀斯特森林退化区凋落物分解动态方面的研究尚不多见。本研究通过对茂兰喀斯特森林退化区内不同类型凋落物的分解动态进行为期18个月的调查,探讨了喀斯特森林退化区凋落物的失重率和分解速率,对比了三种类型凋落物在不同林分中的分解变化以及在相同林分中不同坡位的分解变化,分析了凋落物养分释放特征,初步阐述了产生这些差异的原因,为喀斯特森林退化区的综合治理和生态环境的保护提供科學依据。
1材料与方法
1.1 研究区与样地概况
贵州省茂兰喀斯特自然保护区是国家级自然保护区,位于贵州省黔南布依族自治州荔波县南部,接壤广西壮族自治区。地理位置为107°52′10″—108°05′40″ E,25°09′20″—25°20′50″ N。区内地形为高原山地,喀斯特地貌十分典型,整个保护区呈东北—西南向。该区最高的海拔达1 078.5 m,最低海拔为432 m,平均海拔在802 m以上;成土母岩主要为中下石炭纪白云岩和石灰岩。样地位于茂兰保护区西北部拉桥的缓冲区,选取的喀斯特次生林是处于演替中间阶段的森林群落,而灌木林是演替初级阶段的群落。群落植被概况见表1。由于坡体下部受到人为干扰退化为草坡,故样地选在次生林和灌木林的上坡位和中坡位。
25°18′50″ N林分层次结构分化明显,乔木层、灌木层比较发达,高5~12 m,乔木层覆盖率在80%以上;灌木层高2~3 m,盖度10%左右,地表有少量藤刺、蕨类、地衣苔藓等分布,林下枯枝落叶层厚1~2 cm25°18′35″ N林分垂直结构单一,无或有少量乔木,主要以灌木层为主,高度2~4 m,覆盖度在80%以上,林下覆盖的枯枝落叶层厚为1~2 cm南天竹、圆果化香树、香叶树、荔波鹅耳枥、齿叶黄皮、多脉青冈茂兰喀斯特地区属于中亚热带季风气候,较湿润,冬无严寒,夏无酷暑。年平均温度为18.5 ℃,年平均积温为6 210.8 ℃;年降水量为1 215~1 400 mm,雨季集中在5—9月,全年阴雨天气较多,日照时数相对较少;年相对湿度为77.99%。由图1可知,其气候特征表现为雨热同期,夏季高温多雨,而冬季低温少雨。夏季的平均降雨量、平均气温及平均湿度为全年最高,其中7月份的平均降雨量为271 mm,平均湿度达79%,8月平均气温最高为27 ℃。冬季的平均降雨量、平均气温及平均湿度为全年最低,其中1月份的平均降雨量为12.1 mm,平均湿度为68%,平均气温最低为7 ℃,表现出了冬干夏湿的气候特征。
1.2 数据收集与处理
1.2.1 凋落物的收集于次生林和灌木林中分别收集上坡和中坡凋落物,在设置的每个样地内按照“梅花形”五点法布置5个凋落物收集器,收集器由孔径为0.2 mm × 0.2 mm的尼龙网制成,放置于离地面50 cm的高处,接收面积1 m × 1 m。将全部收集好的凋落物分成树叶和枯枝,树叶按其自身属性分成落叶和常绿叶,将分好类的三种凋落物各自均匀混合,带回实验室,于60 ℃的烘箱中烘干至恒重,分别称取10 g的凋落物装入尼龙网分解袋中(孔径为1 mm,规格为20 cm × 20 cm)。
1.2.2 分解袋的收集2016年7月分别将分解袋沿同一等高线放置于次生林和灌木林的上坡位和中坡位,各坡位分别放入81个分解袋,紧贴地表,覆上少许枯枝落叶,以模拟凋落物分解的自然状态。每隔2个月,分别随机回收每种分解样品各3袋,将回收的凋落物进行室内处理,附着在剩余凋落物表面的沉淀物用清水快速清洗,把分解袋中的根系去掉,放在80 ℃的烘箱中烘干至恒重,测定其干重。实验时间从2016年9月至2018年1月。
1.3 计算公式
1.4 凋落叶N、C元素测定
挑选烘干后的凋落物样品,分别将落叶、常绿叶与枯枝样品磨细,过60目筛后,放置于广口瓶中备测。采用元素分析仪(PE2400-Ⅱ)测定凋落叶N,C含量。
1.5 数据分析方法
采用EXCEL软件和SPSS21.0统计分析软件对数据进行计算和处理,采用Origin9.1软件对数据进行绘图处理。
2结果与分析
2.1 不同类型凋落物的分解动态
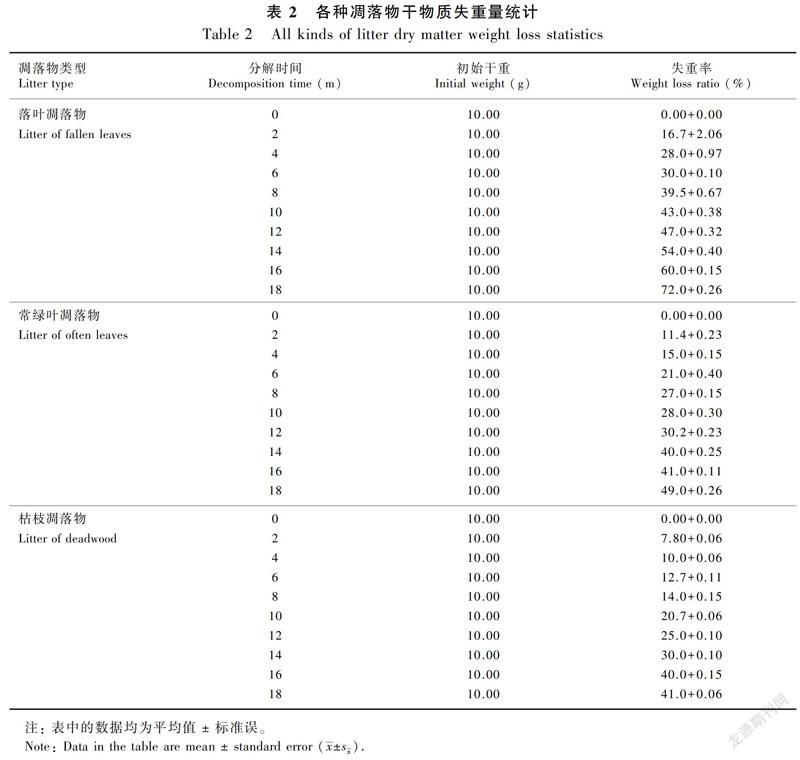
在分解过程中,不同类型的凋落物在不同的分解阶段失重程度各不相同。三种凋落物的失重率总体趋势均随着分解时间的增加而增加,但在不同分解阶段,凋落物的月失重率则有所不同,同一个阶段,不同类型的凋落物失重率也不相同。从表2可以看出,在分解的前2个月,落叶的失重率为16.7%+2.06%,常绿叶为11.4%+0.23%,枯枝为7.80%+0.06%,在分解的第10个月,落叶的失重率为43.0%+0.38%,常绿叶为28.0%+0.30%,枯枝为20.7%+0.06%,在分解的第18个月,落叶的失重率为72.0%+0.26%,常绿叶为49.0%+0.26%,枯枝为41.0%+0.06%。从而得出,三种凋落物分解速率的排序为落叶>常绿叶>枯枝。
2.2 凋落物月失重率在分解过程中的变化
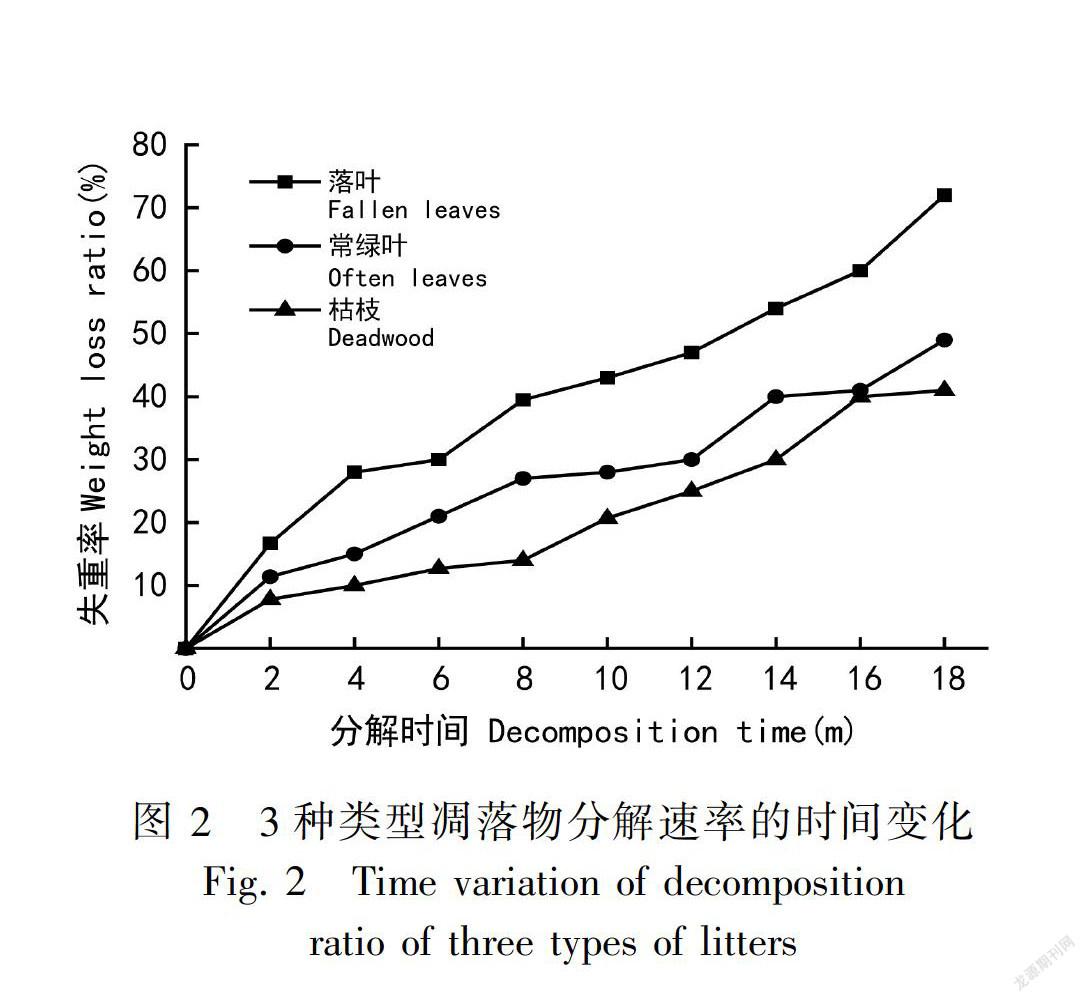
从三种凋落物分解动态(图2)来看,随着分解时间增加,凋落物失重率趋于增加,但不同分解阶段的凋落物月失重率不同。落叶凋落物在前4个月内分解速度非常快,失重率达28.0%,在4~6个月时分解速度变慢,6~8个月时开始加快,8~16个月时分解速度稍慢于前2个月,最后2个月又变快,分解到第18个月时失重率达到了72%;常绿叶和枯枝凋落物在前2个月分解速度快,失重率分别为11.4%和7.8%,常绿叶在分解2~8个月时分解速度慢于前2个月,8~12个月时其残留量下降不明显,在12~14个月时分解速度又变快,在14~16个月时常绿叶凋落物残留量下降幅度变化不大,16~18个月时下降速度再次加快;枯枝凋落物剩余量在2~8个月时下降速度缓慢, 从第8个表 2各种凋落物干物质失重量统计月开始分解速度加快。落叶凋落物的最高峰值失重率发生在分解的前4个月、第8个月和第18个月,而常绿叶的高峰值出现在第8个月和第14个月,枯枝在2~8个月之间,失重率变化不明显,分解速度缓慢,其峰值出现在第2个月和第16个月。总体来看,分解最快的是落叶,其次是常绿叶,最后是枯枝。
2.3 次生林与灌木林凋落叶的失重率变化
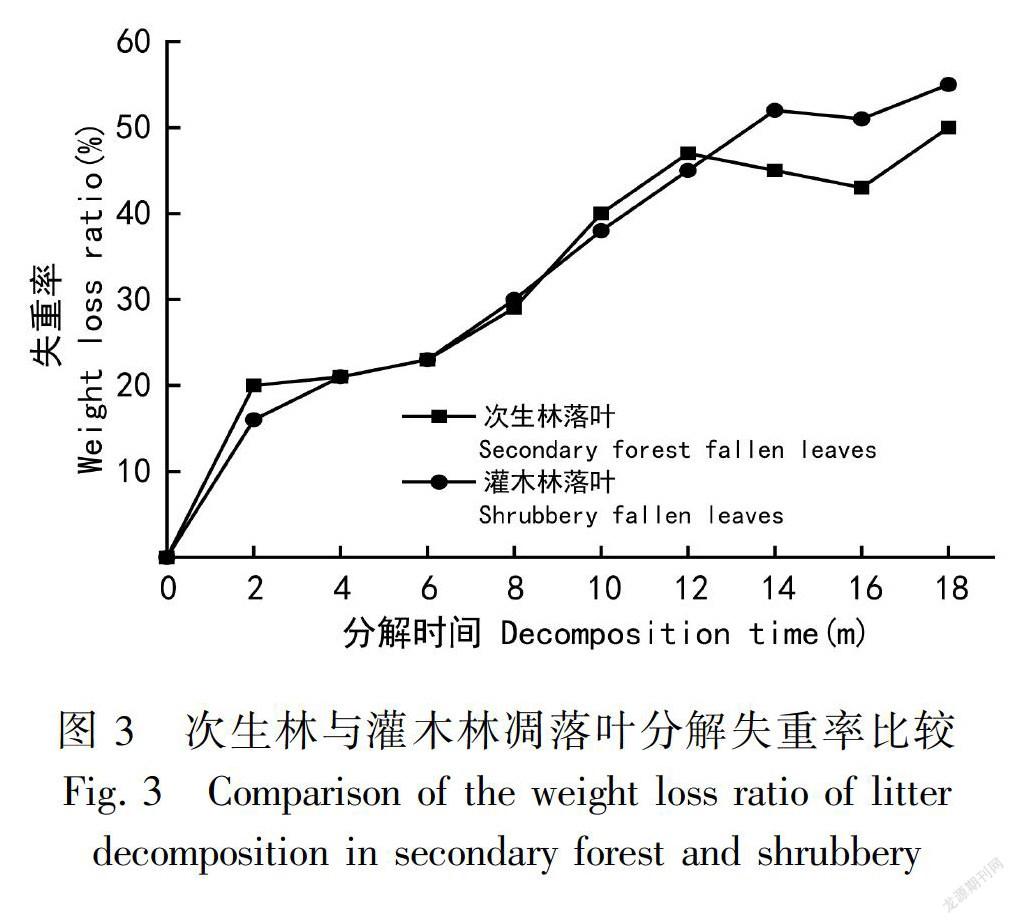
落叶凋落物失重率在次生林中分解前4个月稍快于灌木林中的失重率,而在分解4~11个月时失重率基本同步,分解11~12个月时次生林失重率稍快于灌木林失重率,在分解的第12~18个月时灌木林的失重率就开始大于次生林的失重率,在第16个月时灌木林失重率大于次生林8%;而从次生林和灌木林的常绿叶来看,它们在分解的前10个月中,基本上是同步的,在10~14个月时灌木林失重率大于次生林失重率,第14~16个月又基本同步,在第18个月的时候,灌木林常绿叶的分解速度开始快于次生林(图3)。
2.4 凋落叶在不同林分类型中的分解速率比较
为比较落叶凋落物和常绿叶凋落物在不同林分类型中的分解速率差异,利用Olson指数衰减模型对凋落叶分解50%及95%所需时间进行估算。落叶在不同林分类型中的分解速率不相同,灌木林落叶分解系数为0.471,次生林落叶分解系数为0.376。灌木林落叶凋落物分解50%所需时间1.48 a,分解90%所需时间6.38 a,而次生林落叶凋落物分解50%所需时间1.85 a,分解90%所需时间7.99 a,从而可知落叶凋落物在灌木林中的分解速率大于在次生林中的分解速率。常绿叶凋落物在灌木林中的分解系数为0.734,在次生林中的分解系数为0.371,灌木林常绿叶凋落物分解50%所需时间1.87 a,分解90%所需时间8.10 a,而次生林常绿叶凋落物分解50%所需时间1.95 a,分解90%所需时间8.45 a,表明常绿叶凋落物在灌木林和次生林中的分解速率相差不大,但从总体来看常绿叶凋落物在灌木林中的分解速率大于次生林中的分解速率。落叶凋落物与常绿叶凋落物的分解模型判定系数均很高,拟合效果比较好(R2>0.9,P<0.01)(表3)。
2.5 不同坡位凋落物失重率的变化
次生林和灌木林中三种凋落物在不同坡位的失重率不相同,次生林落叶凋落物失重率在分解前2个月基本同步,后期均为中坡>上坡;次生林常绿叶凋落物的失重率在分解的前14个月为上坡>中坡,在后4个月中,中坡开始大于上坡;而次生林枯枝凋落物的失重率在分解前2个月为中坡>上坡,分解2~6个月时上坡>中坡,分解6个月后均为中坡>上坡。落叶凋落物和枯枝在灌木林中的失重率全程为中坡>上坡;灌木林常绿叶失重率在分解前6個月为上坡>中坡,分解8个月后中坡>上坡。但在整个分解的过程中,失重率对比变化的总体趋势是中坡>上坡(图4)。
2.6 不同类型凋落物养分的动态变化
初始C含量落叶41.53%,常绿叶43.94%,枯枝46.15%,C含量在分解全程为枯枝>常绿叶>落叶,三种凋落物的C含量波动性较大,总体变化趋势随分解时间增加而减少;初始N含量落叶1.29%,常绿叶1.14%,枯枝0.79%,N含量全程表现为落叶>常绿叶>枯枝,三种凋落物的N含量随分解时间增加而增加;初始C/N比值落叶32.10,常绿叶38.48,枯枝58.20,整个分解过程中的C/N比值为枯枝>常绿叶>落叶,随着分解时间增加,三种凋落物的C/N比值均降低(图5)。
3讨论
3.1 不同类型凋落物的分解失重率差异
通过分解实验可以看出,分解过程中凋落物的分解速率是落叶>常绿叶>枯枝。凋落物分解速率在分解初期达最大,随后变缓,分解12个月后分解速率又开始增快,各种凋落物的分解趋势都呈现“快-慢-快”的变化,这种分解进程主要归结于两个方面:一方面,凋落物的分解速率与凋落物中容易分解的成分含量有关,分解样品刚放于林地,降水对溶性养分的强烈淋溶,与此同时,微生物、腐食小动物和其他分解者发现了新的食物来源, 从而加快了凋落物的分解; 另一方面, 与当地表 3凋落物的分解残留率随时间的指数回归方程的气候条件相关,温度、湿度等气候因子影响着土壤生物的活动和土壤生化过程。在分解初期,正值秋冬季节,温度偏低,分解者活动比较弱,夏绿叶和枯枝的分解速度较慢,而在分解8~12个月的时候,正处于夏秋季节,这时是高温高湿的季节,分解者活动旺盛,此时的分解作用十分强烈。Moore et al.(1999)的研究结果表明,随着气温升高,凋落物分解率会增加4%~7%,而本研究发现,夏秋季凋落叶分解率比冬春季凋落叶分解率高6%~8%,两者研究结果相近。总的来说,这是由于分解开始时,可溶性有机物的淋洗和易分解碳水化合物的分解,但到一定时期,难分解的纤维素和单宁等物质的积累,导致分解速度变慢(Mc Claugherty et al., 1985),随着时间推移,组织结构崩溃,分解速度又加快(游巍斌等,2010)。在分解1 a以后,三种凋落物的剩余重量率分别为落叶53.0%,常绿叶70.0%,枯枝75.0%。由此看出,落叶分解最快,另外两类凋落物分解较慢。王希华等(2004)的研究表明,叶凋落物分解受SLA(比叶面积)的影响,SLA越大,代表凋落物和地面接触的面积就越大,使更多的土壤微生物在其上活动的范围就越广泛,从而加速了凋落物的分解,与此同时,小的SLA通常是叶片较厚,角质类物质较多,这对降雨淋溶起到了抑制作用,从而使凋落物的分解变慢。因为落叶叶片较大,为纸质叶片,所以在水热条件适宜时,它们会强烈腐烂。在2018年1月底的时候,落叶凋落物干重流失率高达72.0%,分解袋中的枯叶几乎腐烂,只留下少量的叶状和叶柄,一些活性植物的根部侵入一些分解袋中,同时还有个别的地衣类植物开始在里面生长。常绿叶类凋落物叶片相对较小,为厚革质,即使在相同条件下,分解也显得慢些,最高失重率为49.0%。在机械分解过程中,常绿叶凋落物分解的作用没有落叶凋落物表现得那么强烈。而枯枝由于含纤维素较多,且质厚,分解则显得更为缓慢。
三种类型的凋落物达到分解高峰值的时间不一样,森林凋落物的分解一般是先快后慢,包括粉碎、物理淋溶和有机物代谢三个分解过程(Wood, 1991),落叶面积比较大,从自身构造上看韧度较低,表面没有蜡质保护膜, 在微生物的作用下很容图 5不同类型凋落物的C、N、C\\N含量比较易分解,同时也易受到物理粉碎作用,因此在很短的时间就达到了分解高峰。常绿叶的叶片是革质,且表面通常具有蜡质的保护膜,叶内的养分很少流失,但如果粉碎到一定程度后,就会开始进行化学分解,微生物利用其中的养分合成分解酶,迅速对其进行分解(严海元等,2010),所以由于常绿叶的叶片结构,它的分解速率高峰会到达晚一些。枯枝分速率最慢,分解速率高峰到达最迟,枯枝主要是由纤维素和木质素组成,这两种物质在分解过程中很难被物理分解和化学分解,要经过很长时间才能实现其成熟破碎,养分释放也比较慢,微生物对枯枝的作用不能很快实现,因此枯枝的分解速率慢(宋福强等,2009)。
3.2 次生林和灌木林对分解差异性的影响
在相同的气候条件下,森林类型是影响凋落物分解速率的重要因素,凋落物分解速率随林分组成、林分密度不同而变化(Facelli & Pickett, 1991)。本研究中,通过对次生林与灌木林中凋落物的分解速率比较,结果表明两种林分的落叶凋落物的分解速率在前期基本上是次生林的失重率稍大于灌木林的,而在进行分解的后期,却出现了落叶凋落物在灌木林中的分解速率大于次生林的。首先常绿叶凋落物前10个月在两种林分中的分解速率基本上是同步的,随着分解时间的增加,在灌木林中的失重率稍大于在次生林中的,然后两者又几乎同步,最后常绿叶在灌木林中的分解速度大于在次生林中的。首先,是因为群落的物种组成、群落林分密度、群落郁闭度不同而导致的(刘玉国等,2011),喀斯特次生林的林分层次结构分化明显,乔木层、灌木层比较发达,地表有少量藤刺、蕨类、地衣苔藓等分布,而喀斯特灌木林的林分垂直结构单一,无或有少量乔木,主要以灌木层为主。其次,次生林和灌木林分别处于阳坡和阴坡,因为坡向不同会造成林分水热状况的差异,必然会影响到林中凋落物分解速度的快慢(程煜等,2011)。灌木林处于阴坡,植被生活型都以灌木为主,地面接受太阳光照不多,湿度比较大,温暖潮湿的环境使土壤微生物活动旺盛,有利于有机物的迅速分解,从整体而言,不论是落叶凋落物还是常绿叶凋落物在灌木林中的分解速率都大于次生林的。
3.3 坡位对凋落物分解差异性影响
Vitousek et al.(1994)学者对太平洋熱带岛屿Mauna Loa的研究表明,随着海拔升高,气温下降,凋落物分解速率呈指数下降。山体不同坡位的水分条件存在着一定的差异,由于受到环境气候及光照的影响,从而导致不同坡位林分内的温度和湿度不同(Hornsby et al., 1995)。当坡位不同时,表现出各种凋落物分解速率的总体趋势为中坡>上坡,因为上坡位接受到的太阳辐射较多、温度较高、土层很薄、土壤养分含量和土壤含水量较低的原因,不利于凋落物的分解,随着富含养分的表层土壤随坡面流向中下坡汇集,导致中、下坡位的土壤氮含量和有机质含量相对较高,这对凋落物的分解起到了很大的促进作用,在养分含量较高的地方,微生物的活动比较强烈,使凋落物养分归还地表的过程大大缩短,养分循环加快,但是由于上坡和中坡的高度相差不是太大,所以这种差异不是太明显。但在次生林常绿叶分解的前期,出现了上坡>中坡,对于这个结果,本研究目前还无法验证,有待下一步的实验分析来阐述。
3.4 不同类型凋落物养分动态
不同类型凋落物的化学组成不同,从而影响它们的分解速率。落叶凋落物的初始C含量为41.53%,常绿叶凋落物的初始C含量为43.94%,枯枝凋落物的初始C含量為46.15%,枯枝凋落物的C含量大于相同时期的叶凋落物。较高的C含量是由于凋落物需合成有机物质,从而维持其生长和代谢。在凋落物分解过程中,N元素是制约凋落物分解的一个很重要的因素(曾锋等,2010)。在本研究中,落叶凋落物的初始N含量为1.29%,常绿叶凋落物的初始N含量为1.14%,枯枝凋落物的初始N含量为0.79%,分解的整个过程中,都是落叶凋落物的N含量>常绿叶凋落物的N含量>枯枝凋落物的N含量,很多学者研究表明,N含量高的凋落物其分解速率比较快(王相娥等,2009),分解时的N元素的释放模式为富集—释放模式,氮元素的富集在凋落物分解过程中普遍存在,因为在凋落叶分解过程中,微生物活动对氮元素有明显的需要,当凋落叶中初始氮含量较低时,微生物会从外界环境中吸收氮元素来满足自身活动的需要,这样便引起了氮元素含量上升(窦荣鹏等,2010)。因此,在分解的初始阶段N的绝对含量趋于增加(许晓静等,2007),本文的研究与之相一致。Melin(1930)在发表《北美几种森林凋落物的生物分解特征》一文中,使用了C/N比来分析凋落物的分解特征,C/N比后来成了评价凋落物分解的经典指标。本研究中,C/N比值随着三种不同类型凋落物的分解呈现不断下降趋势,这是由于氮沉降会导致N元素增多,凋落物分解到后期,难以分解的物质相对增加,分解速率变小,从而C/N比值逐渐降低(曾昭霞等,2011)。本研究结果表明,枯枝凋落物C含量>常绿叶凋落物C含量>落叶凋落物C含量,落叶凋落物的初始N含量>常绿叶凋落物的初始N含量>枯枝凋落物的初始N含量,C/N比值基本上表现为枯枝凋落物>常绿叶凋落物>落叶凋落物,分解速率为落叶凋落物>常绿叶凋落物>枯枝凋落物。
不同类型凋落物在不同林分中存在着不同分解速率即是各种因素综合作用的结果。喀斯特地区的森林生态系统比较脆弱,一旦遭到破坏,就很难恢复成原来的生态系统结构。森林凋落物是覆盖在地表上的腐殖质,其结构疏松,不仅具有吸水能力,而且透水性很强,能够降低径流速度,丰富地下水源,减少土壤溅蚀,在水土保持、水源涵养方面发挥着巨大的作用。因此,研究喀斯特森林退化区凋落物的分解速率变化,有利于相关部门更好地了解植被恢复与生态系统重建的优势条件,为喀斯特森林退化区的综合治理和生态环境的保护提供科学依据。
参考文献:
BERG B, BERG MP, BOTTNER P, 1993. Litter mass loss rates in pine forests of Europe and Eastern United States: Some relationships with climate and litter quality [J]. Biogeochemistry, 20: 127-53.
BERG B, MC CLAUGHERTY C, 1989. Nitrogen and phosphorus release from decomposing litter in relation to the disappearance of lignin [J]. Can J Bot, 67, 1148-1156.
CHEN Y, FENG T, YU YL, et al., 2017. Study on the dynamics of litters in three plant communities in the karst mountainous area of Northwest Guizhou [J]. J Guizhou Inst Eng Appl Technol, 35(3): 73-78. [陈元, 冯图, 喻元良, 等, 2017. 黔西北喀斯特山区三种植物群落凋落物动态研究 [J]. 贵州工程应用技术学院学报, 35(3):73-78.]
CHENG Y, CHEN C, FAN HL, et al., 2011. Effects of different slope directions on litter decomposition and nutrient release rate of schima superba masson pine [J]. Chin Agric Bull, 27(31): 6-17. [程煜, 陈灿, 范海兰, 等, 2011. 不同坡向对木荷马尾松凋落物分解及养分释放速率的影响 [J]. 中国农学通报, 27(31):6-17.]
DOU RP, JIANG H, YU SQ, et al., 2010. Decomposition of cryptomeria fortunei litter in subtropical and tropical China [J]. Acta Ecol Sin, 30(7): 1758-1763. [窦荣鹏, 江洪, 余树全, 等, 2010. 柳杉凋落物在中国亚热带和热带的分解 [J]. 生态学报, 30(7):1758-1763.]
EBERMAYER E, 1876. Die qesamte Lehre der woldstreu mit Rucksicht auf die chemische statik woldbaues [M]. Berlin: Julius Springer: 116.
EDMONDS RL, THOMAS TB, 1995. Decomposition and nutrient release from green needles of western hemlock and Pacific silver fir in an old-growth temperate rain forests, Olympic National Park, Washington [J]. Can J For Res, 25: 1049-1057.
FACELLI JM, PICKETT STA, 1991. Plant litter: Its dynamics and effects on plant community structure [J]. Bot Rev, 57(1): 1-32.
HORNSBY DC, LOCKABY BG, CHAPPELKA AH, 1995. Influence of microclimate on decomposition in Loblolly pine stands: A filed microcosm approach [J]. Can J For Res, 25: 1570-1577.
LIU YG, LIU ZC, WEI YF, et al., 2011. Species composition and community structure characteristics of different vegetation succession stages in Puding County of Guizhou Province [J]. J Plant Ecol, 35(10): 1009-1018. [刘玉国, 刘长成, 魏雅芬, 等, 2011. 贵州省普定县不同植被演替阶段的物种组成与群落结构特征 [J]. 植物生态学报, 35(10):1009-1018.]
LU XQ, YANG WX, DING FJ, et al., 2014. Dynamics of litters and nutrient restoration in primary forest in karst Maolan area [J]. J Ecol Rural Environ, 30(5): 614-619. [卢晓强, 杨万霞, 丁访军, 等, 2014. 茂兰喀斯特地区原始林凋落物量动态与养分归还 [J]. 生态与农村环境学报, 30(5):614-619.]
MC CLAUGHERTY CA, PASTOR J, ABER JD, et al., 1985. Forest litter decomposition in relation to soil nitrogen dynamics and litter quality [J]. Ecol, 66: 266-275.
MELIN E, 1930. Biological decomposition of some types of litter from North American forests [J]. Ecol, 11: 72-101.
MOORE TR, TAYLOR B, PRESCOTT C, 1999. Litter decomposition rates in Canadian forests [J]. Glob Change Biol, 5(1): 75-82.
OLSON JS, 1963. Energy storage and the balance of producers and decomposers in ecological systems [J]. Ecol, 44(2): 322-331.
QIAN ZM, RAN JC, RONG L, et al., 2009. Dynamic study on litters of karst forest in Maolan [J]. Anhui Agric Sci, 37(8): 3485-3487+3523. [錢正敏, 冉景丞, 容丽, 等, 2009. 茂兰喀斯特森林凋落物量动态研究 [J]. 安徽农业科学, 37(8):3485-3487+3523.]
SINGH KP, SINGH PK, TRIPATHI SK, 1999. Litterfall, litter decomposition and nutrient release patterns in four native tree species raised on coalmine spoil at Singrauli, India [J]. Biol Fert Soil, 29: 371-378.
SONG FQ, FAN XX, ZHANG XX, et al., 2009. Enzyme activity of three filamentous fungi in decomposing litter of pinus camphor [J]. Microbiol Bull, 36(5):635-639. [宋福强, 范晓旭, 张星星, 等, 2009. 三株丝状真菌分解樟子松凋落物酶活性 [J]. 微生物学通报, 36(5):635-639.]
SUN OJ, CAMPBELL J, LAW BE, et al., 2004. Dynamics of carbon stocks in soils and detritus across chronosequences of different forest types in the Pacific Northwest USA [J]. Glob Chang Biol, 10: 1470-1481.
SWIFT MJ, HEAL OW, ANDERSON JM, 1979. Decomposition in terrestrial ecosystems [J]. Berkeley: University Californ Press: 1-372.
TANG SS, YANG WQ, YIN R, et al., 2014. Distribution characteristics and controlling factors of decomposition rate of litters in Chinese forest ecosystem [J]. J Plant Ecol, 38(6): 529-539. [唐仕姗, 杨万勤, 殷睿, 等, 2014. 中国森林生态系统凋落叶分解速率的分布特征及其控制因子 [J]. 植物生态学报, 38(6):529-539.]
VITOUSEK PM, TURNER DR, PARTON WJ, et al., 1994. Litter decomposition on the Mauna Loa environment matrix, Hawaii I: Patterns, mechanisms, and models [J]. Ecology, 75(2): 418-429.
WANG XH, HUANG JJ, YAN ER, et al., 2004. Study on the decomposition of litters of common plants in Tiantong National Forest Park [J]. J Plant Ecol, 28(4): 457-467. [王希华, 黄建军, 闫恩荣, 2004. 天童国家森林公园常见植物凋落叶分解的研究 [J]. 植物生态学报, 28(4):457-467.]
WANG QL, 2009. Ecological function of litters in several karst secondary forests in central Guizhou [D]. Guiyang: Guizhou Norm University. [王庆玲, 2009. 黔中地区几种喀斯特次生林凋落物生态功能研究 [D]. 贵阳: 贵州师范大学.]
WANG FY, 1989. A review of studies on forest litter [J]. Ecol Prog, 6(2): 82-98. [王凤友, 1989. 森林凋落量研究综述 [J]. 生态学进展, 6(2):82-98.]
WANG XE, XUE L, XIE TF, et al., 2009. A review of studies on litter decomposition [J]. Soils, 40(6): 1473-1478. [王相娥, 薛立, 谢腾芳, 等, 2009. 落物分解研究综述 [J]. 土壤通报, 40(6):1473-1478.]
WOOD TG, 1991. Field investigation on the decomposition of leave of Eucalyptus delegatensis in relation to environmental factors [J]. Pedobiol, 14: 343-371.
XIONG HF, WANG SJ, RONG L, et al., 2013. Dynamics of plant community litters in different succession stages in Puding karst area [J]. Chin J Ecol, 32(4): 802-806. [熊紅福, 王世杰, 容丽, 等, 2013. 普定喀斯特地区不同演替阶段植物群落凋落物动态 [J]. 生态学杂志, 32(4):802-806.]
XU XJ, ZHANG K, LIU B, et al., 2007. Research progress on forest litter decomposition [J]. Sci Water Soil Conserv Chin, (4): 108-114. [许晓静, 张凯, 刘波, 等, 2007. 森林凋落物分解研究进展 [J]. 中国水土保持科学, (4):108-114.]
YAN HY, GU XR, SHEN H, et al., 2010. Microbial decomposition of forest litter [J]. J Ecol, 29(9): 1827-1835. [严海元, 辜夕容, 申鸿, 2010. 森林凋落物的微生物分解 [J]. 生态学杂志, 29(9):1827-1835.]
YANG AX, PENG Y, 2007. Study on hydrological and ecological functions of karst forest ecosystem in Guizhou [J]. Anhui Agric Sci, 35(36): 11995-11997+12037. [杨安学, 彭云, 2007. 贵州喀斯特森林生态系统水文生态功能的研究 [J]. 安徽农业科学, 35(36):11995-11997+12037.]
YOU WW, LIU YS, HE DJ, et al., 2010. Decomposition characteristics of natural forest litter in Wuyi Mountain scenic and historic interest area [J]. J Sichuan Agric Univ, 28(2):141-147. [游巍斌, 刘勇生, 何东进, 等, 2010. 武夷山风景名胜区不同天然林凋落物分解特征 [J]. 四川农业大学学报, 28(2):141-147.]
YU GS, WANG SJ, RONG L, et al., 2011. Litter dynamics of major succession communities in Maolan karst forest [J]. J Plant Ecol, 35(10): 1019-1028. [俞国松, 王世杰, 容丽, 等, 2011. 茂兰喀斯特森林主要演替群落的凋落物动态 [J]. 植物生态学报, 35(10):1019-1028.]
ZENG F, QIU ZJ, XU XY, et al., 2010. Research progress on forest litter decomposition [J]. J Ecol Environ, 19(1): 239-243. [曾锋, 邱治军, 许秀玉, 等, 2010. 森林凋落物分解研究进展 [J]. 生态环境学报, 19(1):239-243.]
ZENG SX, LIU XL, SONG XJ, et al., 2011. Litter amount and degradation of primary forest and secondary forest in karst area of Northwest Guangxi [J]. J Ecol, 30(2): 201-207. [曾昭霞, 刘孝利, 宋希娟, 等, 2011. 桂西北喀斯特区原生林与次生林凋落物量及其降解 [J]. 生态学杂志, 30(2):201-207. ]
ZHAO C, LONG J, LI J, et al., 2018. Litter stock and nutrient characteristics in different slope directions and decomposition layers of karst forest in Maolan [J]. J Ecol, 37(2): 295-303. [赵畅, 龙健, 李娟, 等, 2018. 茂兰喀斯特原生林不同坡向及分解层的凋落物现存量和养分特征 [J]. 生态学杂志, 37(2):295-303.]
ZHANG H, ZHUANG XY, et al., 2008. Decomposing ability of dead leaves of 4 native broad-leaved trees in South China [J]. Acta Ecol Sin, 28(5): 2395-2403. [张浩, 庄雪影, 2008. 华南4种乡土阔叶树种枯落叶分解能力 [J]. 生态学报, 28(5):2395-2403.]
ZHANG DQ, YE WH, YU QF, et al., 2000. Study on representative forest litter in Dinghu Mountain succession series [J]. Acta Ecol Sin, 20(6):938-944. [张德强, 叶万辉, 余清发, 等, 2000. 鼎湖山演替系列中代表性森林凋落物研究 [J]. 生态学报, 20(6):938-944.]
ZHU SQ, WEI LM, ZHANG CG, et al., 1995. A preliminary study on the growth characteristics of tree species in karst forest in Maolan [J]. J Guizhou Agric Univ, (1): 8-16. [朱守謙, 魏鲁明, 张从贵, 等, 1995. 茂兰喀斯特森林树种生长特点初步研究 [J]. 贵州农学院学报, (1):8-16.]
ZHU SY, WANG KL, ZENG FP, et al., 2009. Characteristics of nutrient restoration of litter in karst secondary forest in Northwest Guangxi [J]. J Ecol Environ, 18(1): 274-279. [朱双燕, 王克林, 曾馥平, 等, 2009. 桂西北喀斯特次生林凋落物养分归还特征 [J]. 生态环境学报, 18(1):274-279.]
猜你喜欢
中国-东盟博览(旅游版)(2021年6期)2021-08-09
阅读(低年级)(2020年8期)2020-11-06
军事文摘·科学少年(2019年3期)2019-03-29
老区建设(2017年2期)2017-04-13
中国周刊(2017年2期)2017-02-23
湖北农业科学(2016年20期)2017-02-15
未来英才(2016年20期)2017-01-03
当代贵州(2014年36期)2015-05-28
江苏农业科学(2015年1期)2015-04-17
文化月刊·下旬刊(2014年6期)2014-08-28
