
植物类病变突变体的研究进展
2020-02-29 02:40刘思辰曹晓宁王海岗王君杰陈凌田翔秦慧彬乔治军
山西农业科学 2020年2期
刘思辰,曹晓宁,王海岗,王君杰,陈凌,田翔,秦慧彬,乔治军
(山西省农业科学院农作物品种资源研究所,农业部黄土高原作物基因资源与种质创制重点实验室,杂粮种质资源发掘与遗传改良山西省重点实验室,山西太原030031)
植物类病变(lesion mimic)是指在无外界致病因素和逆境因素的影响下,植株叶片、叶鞘等部位自发产生坏死斑的现象。多种植物中均发现类病变突变体[1]。类病变突变与植物过敏性反应(hypersensitive response,HR)症状类似[2],可在类病变产生部位发生细胞坏死,属于程序化细胞死亡(programmed cell death,PCD),而且许多斑点叶突变体对某些植物病原物表现出了一定的抗性[3]。因此,研究类病变突变体对了解细胞的程序性死亡以及植株抗病反应机制具有重要意义[4]。类病变突变体也是抗病种质资源的重要来源。在大麦中发现的类病变突变体mlo对白粉病生理小种都具有持久抗性[5],类病变突变体mlo在欧洲许多国家已经大面积种植。
本研究通过综述植物类病变突变体的来源、类病变表型的形成机制及类病斑基因的研究进展,对深入研究植物类病变突变体对作物抗病的分子机理研究和栽培品种的遗传改良都具有重要的意义。因此,类病变突变体的研究也能够为具有较宽的抗谱、抗性持久的抗病基因提供丰富的种质资源。
1 类病变变异的来源
类病变突变体主要来源有2种,即自然变异和人工诱变。
1.1 自然变异
自然变异频率很低,仅为10-8~10-5,此类突变体占比较小。1965年,SEKIGUCHI等发现了水稻第一个自然突变的类病变突变体sl(sekiguchi lesion),该突变性状受1对隐性基因控制。随后,bl1、bl2和spl1等水稻自然类病变突变体被相继发现[6]。
1.2 人工诱变
其主要包括物理、化学和生物诱变等。物理诱变是指利用各种高能射线进行照射处理植物种子进行人工诱发变异,如水稻突变体lmi、spl3、spl5、rls1等[7]。化学诱变是利用化学诱变剂(如烷化剂、碱基类似物、抗生素等)处理植物组织,最常用的诱变剂是EMS。如lsd1、acd1、lmm1、spl11、spl29和les23等都是通过EMS诱导获得的类病变突变体[7-8]。生物诱变主要包括T-DNA插入[9]、转座子插入[6]等发生的变异,导致植株出现病斑表型。此外,在植物组织培养过程中体细胞再生也会导致类病变的发生。
2 类病变突变体的特征
类病变主要发生部位是叶片或叶鞘,其病斑的颜色和性状不同,病斑出现的时期也有差异,多数出现在苗期也就是播种后2~3周(如水稻类病变突变体spl1~spl4)。但也有些突变体的斑点直到孕穗期才出现,如水稻类病变突变体sp16。根据病斑的扩展方式,类病变突变体也可分为起始型和扩散型2类[10]。起始型突变体斑点是一种局部自发性的细胞坏死现象,不向周边蔓延扩散,如blm和M1009。扩散型突变体的病斑会扩散到整个叶片及植株,如spl1和lmm1[7]。
3 类病变突变体的遗传模式
大多数突变体为单个基因控制,主要受1~2个基因控制,在玉米中多数为显性突变,定位玉米斑点突变的基因分布在各个染色体上。小麦中已报道的类病变突变体有7个,除了突变体C591(M8)受显性基因控制外,其余均为隐性遗传。迄今报道的200多例水稻斑点突变,已经对80多份类病变突变体材料进行了遗传分析和鉴定,类病变性状主要受1~2个核基因控制,在这些突变体中有73个受单隐性基因控制。
4 类病变基因的克隆与功能分析
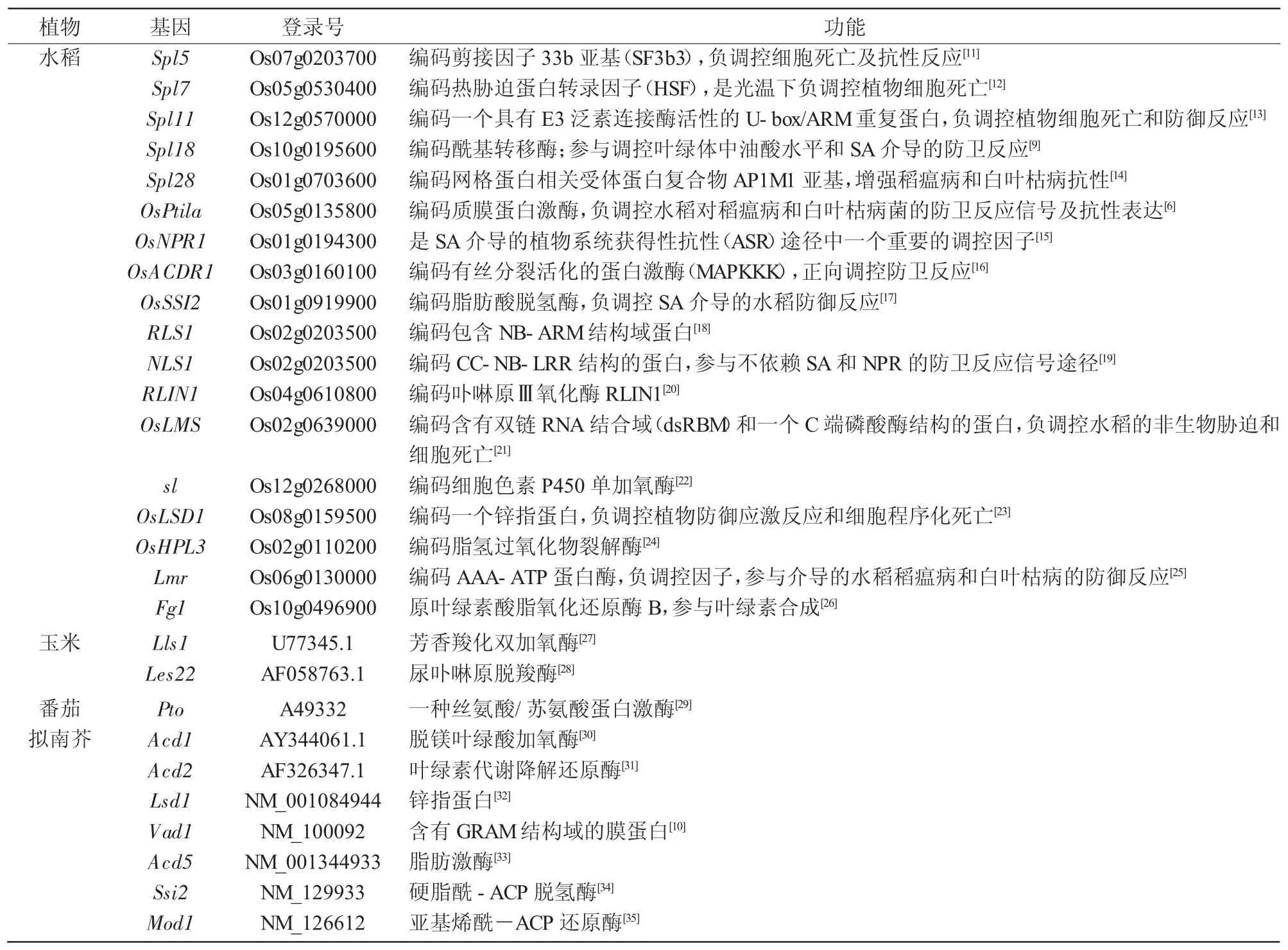
表1 植物类病变基因
自第1例类病变突变体报道以来,研究人员利用各种分子遗传学技术,在拟南芥、玉米、大麦、水稻、番茄等的类病变突变体中克隆到多个控制类病变性状的基因(表1)。
5 类病变突变体表型的形成机制
5.1 抗病基因的改变或功能缺失
植物抗病基因的改变启动了过敏反应,导致植物细胞程序性死亡提前发生,诱发植物发生类病斑的症状。大麦抗白粉病基因Mlo的碱基突变致使其突变体自发形成坏死斑[20]。水稻NLS1基因、玉米RP1基因及番茄Pto基因的突变,会引发过敏性反应和程序性细胞死亡,形成坏死病斑。
5.2 正常代谢途径紊乱
植物正常代谢紊乱会使植物PCD途径受到干扰,有可能引起类病变表型的发生。主要涉及脂肪酸、氨基酸以及卟啉代谢途径等。玉米类病变les22基因突变显著降低尿卟啉原脱羧酶活性,诱导植株产生病变表型[5]。水稻中nls1基因突变伴随防御反应组成性表达,其突变体中活性氧大量积累,导致类病变表型的发生[28]。水稻rlin1突变体由于编码粪卟啉原III氧化酶的基因发生单碱基突变致使该酶活性降低,影响粪卟啉原Ⅲ氧化形成原卟啉Ⅸ的生化反应过程,导致植株出现类病变表型[21]。拟南芥SSCD1基因突变后络氨酸降解异常,植株出现病斑表型[31]。MOU等[35]研究发现,编码拟南芥脂肪酸合成的关键酶(烯酰-ACP还原酶enoyl-ACP reductase,ENR)的mod1基因发生突变后,烯酰基酰基载体蛋白还原酶的活性显著降低,阻碍脂肪酸的合成,植株表现出类病变表型。水稻中多不饱和脂肪酸合成途径中的关键酶OsSSI2基因突变导致类病变的产生[17]。
5.3 活性氧在植物体内的积累
类病变突变体坏死病斑的产生往往伴随着活性氧物质的积累,这些富集的ROS可以直接破坏正常细胞的结构,从而导致类病变的产生。玉米les22突变体的叶片细胞坏死就是由活性氧积累直接造成的[28]。拟南芥突变体rcd1中过氧化物(O2-)和臭氧分子(O3)的过量积累导致产生坏死病斑[36-37]。水稻noe1突变体中编码基因改变后,叶片中H2O2过量积累,最终诱导突变体植株产生类病变的表型[38]。
5.4 植株体内激素失调
激素信号在类病变表型发生的过程中起着重要的作用,主要是乙烯、水杨酸和茉莉酸,这几种激素代谢失调均有可能导致类病变的发生。水稻OsSSI2基因的突变导致内源水杨酸含量和防御因子WRKY45表达量的上调[17]。水稻中敲除OsEDR1基因影响乙烯的合成,破坏体内的激素平衡,从而诱发产生类病变坏死病斑[16]。拟南芥突变体hrl1中乙烯、水杨酸过量也会导致病变表型的产生。上述结果暗示,植物激素可能参与植物类病变表型的发生。
5.5 生长环境因素
生长条件同样可以诱导植物类病变表型的产生。玉米类病变突变体les22的类病变表型受光照的诱导[28]。刘道峰等[39]用锡纸遮光处理lmi突变体叶片,发现遮光部位无坏死斑点形成,表明lmi突变体细胞受光照诱导。MATEO等[40]构建了lsd1cao双突变体探讨病斑形成与光系统II的关系,推测lsd1类病变的表达与光合系统II有关。王建军等[41]对6个水稻lrd类病变突变体进行不同温度梯度处理,发现低温明显促进突变体lrd35和lrd40类病变发生,突变体lrd31在中温下出现坏死斑。JAMBUNATHAN等[42]研究发现,拟南芥cpnl突变体属于湿度敏感型突变体,通过调控Ca2+流向负向调控细胞死亡,产生类病变表型。
6 类病变突变体的抗病性研究
目前,已鉴定的很多类病变突变体或多或少都表现出增强的抗病性,不同的突变体抗病性存在较大的差异[42]。水稻突变体spl1、spl5、spl9和spl11表现出稻瘟病抗性增强,spl21、lmes1、hm197等12个突变体为白叶枯病抗性增强,突变体lmm1则是纹枯病抗性增强。这些突变体的抗病谱狭窄,其在育种上的应用并不多。一些具有广谱抗性的突变体有非常高的实际应用价值,例如,在大麦中发现的类病变突变体mlo,对白粉病具有广谱性抗性,在欧洲许多国家已经大面积种植。水稻中也有具有广谱抗性的类病变突变体如spl11、lrd40、spl5等,为水稻抗病育种提供了非常重要的材料来源。
7 展望
类病变突变体是研究植物对病原菌的防御机制和程序性细胞死亡的理想材料。通过对类病变突变体研究,发现类病变的研究不仅涉及程序性细胞死亡、抗病抗逆生理以及HR反应产生的活性氧与自由基等方面,还与光信号及温度诱导信号等有着密切的联系。植物对病原菌的广谱抗性通常在其类病变突变体中表现出来,因此,类病斑突变体是植物育种中重要的抗病种质资源。然而,部分类病变突变体还伴随着株高降低、产量下降、分蘖能力减弱、早衰、雄性不育等农艺性状的改变[42-43],使得类病变突变体中表现出来的广谱抗性直接应用到农业生产中存在困难。因此,发掘对农艺性状影响小、并具有广谱抗性的类病变突变体材料在抗病育种上具有重大意义。
猜你喜欢
作物学报(2022年2期)2022-11-06
亚热带农业研究(2022年1期)2022-08-08
今日农业(2022年4期)2022-06-01
作物学报(2022年8期)2022-05-29
蔬菜(2021年7期)2021-11-27
发明与创新·大科技(2019年5期)2019-07-31
飞碟探索(2015年9期)2015-11-05
红领巾·探索(2015年9期)2015-09-10
恋爱婚姻家庭·养生版(2011年8期)2011-05-14
中学生物学(2008年3期)2008-06-03
