
2种海水臂尾轮虫品系生活史特征
2020-03-03 14:37张凯雷马慧娟李茹梦杨家新
江苏农业科学 2020年22期
张凯雷 马慧娟 李茹梦 杨家新

摘要:应用单个体和种群培养方法,研究在一定温度(20℃)下,以浓度为0.25×106、0.5×106、1×106、2×106、4×106、8×106cells/mL的海水小球藻(Chlorellasp.)为食,比较射阳褶皱臂尾轮虫(Brachionusplicatilis,SY)和玛雅臂尾轮虫(Brachionusmanjavacas,BM)的生活史特征。研究表明,SY品系和BM品系的种群增长率均保持先增长后缓慢下降的趋势。BM品系和SY品系在初期食物浓度为4×106cells/mL时的种群增长率始终高于其他浓度处理;在后期,食物浓度为8×106cells/mL的种群增长率才最高。BM品系的种群数量与种群增长率变化趋势一样;而SY品系在食物浓度为8×106cells/mL时的种群数量一直高于其他浓度。2个品系轮虫的生殖前期都随食物浓度升高而降低。在(0.25×106、0.5×106cells/mL)时,BM品系的生殖前期比SY品系的生殖前期要长;在(4×106、8×106cells/mL)时,SY品系的生殖前期比BM品系长;在(1×106、2×106cells/mL)时,2个品系轮虫生殖前期无显著差异。2个品系褶皱臂尾轮虫的子代数均随食物浓度增大而变多,在低浓度时,SY品系子代数高于BM品系;在中高浓度时,BM品系子代数高于SY品系。2个品系轮虫的寿命随着食物浓度升高先增长后缩短,都在食物浓度为4×106cells/mL或8×106cells/mL时的寿命最长。
关键词:褶皱臂尾轮虫;品系;食物浓度;生殖前期;子代数;寿命
中图分类号:S963.21+4文献标志码:A
文章编号:1002-1302(2020)22-0169-05
通信作者:杨家新,男,教授,博士生导师,从事轮虫繁殖生物学研究。E-mail:1063859395@qq.com。
褶皱臂尾轮虫(Brachionusplicatilis)作为海水人工育苗的一种轮虫,其繁殖状况的不同都会在生产上带来不同的效益。当然,生殖前期越短,轮虫在前期就会繁殖越快;寿命越长,维持种群高密度的时间就越长;子代数越高,密度繁殖越快。在不同食物浓度条件下,种群增长率、种群数量的变化以及生殖前期、寿命、子代数也会不同。已有研究表明,温度和食物浓度的不同会对轮虫生殖期产生影响[1-2],品系的不同也会对萼花臂尾轮虫的生活史产生影响[3]。也有研究发现。萼花臂尾轮虫的姐妹种和它们的地理位置并没有显著差异[4]。Rehberg-Haas等也研究了不同巴芙藻对褶皱臂尾轮虫的影响[5]。Cheng等研究了盐度和食物浓度对圆形臂尾轮虫种群动态的影响[6]。除了藻种,还有轮虫品系、温度、盐度、光照和食物浓度等都对褶皱臂尾轮虫生长都具有影响,由于环境因子的多样性,褶皱臂尾轮虫的生殖发育也会发生不同改变。
本试验以褶皱臂尾轮虫(SY)和玛雅臂尾轮虫(Brachionusplicatilis,BM)这2个品系轮虫为研究对象,探讨在不同食物浓度下2个品系轮虫的生长状况及生态学参数(生殖前期、寿命、子代数)的差异。为寻找2个品系轮虫在不同生长阶段最适的食物浓度范围,确认不同的食物浓度对轮虫的影响及是否与品系有关,本研究对中国射阳品系和美国品系的轮虫进行研究比较。美国BM品系之所以能够成为广泛的研究对象,是因为其具有稳定的特点,本研究想通过试验来确认中国SY品系轮虫是否也像BM品系一样具有稳定的特点,能否成为中国本土特有的品系,并能够成为一种广泛的试验对象。
1材料与方法
1.1轮虫的来源和预培养
1.1.1轮虫来源SY和BM品系都是从冷冻休眠卵中孵化而来的,SY品系休眠卵底泥是从射阳池塘中获取,BM品系休眠卵是美国Snell教授所赠。轮虫休眠卵采用冯大庆的人工海水进行孵化,孵化温度为(20±1)℃,光照度为4000lx,光∶暗(L∶D)=1∶1[7]。在实验室中对这2个品系轮虫进行单个体克隆培养。轮虫培养所需饵料由F2培养基培养:将处于指数增长期的海水小球藻离心浓缩,再加海水培养基配制成悬液喂养轮虫。F2培养基(1L海水中所加的量):1mL7.5g/100mLNaNO3、[JP3]1mL0.5g/100mLNaH2PO4·H2O、1mL3g/100mLNaSiO3·9H2O、1mL微量元素溶液、1mL維生素溶液。[JP3]其中,100mL微量元素溶液含有0.315gFeCl3·6H2O、0.436gNa2EDTA·2H2O、0.98mgCuSO4·5H2O、0.63mgNa2MoO4·2H2O、2.2mgZnSO4·7H2O、1.0mgCoCl2·6H2O、18.0mgMnCl2·4H2O。50mL维生素溶液含有0.05mg维生素B12、0.05mg维生素H、0.01g维生素B1。
1.1.2预培养将SY和BM品系轮虫分别放置于6种食物浓度(0.25×106、0.5×106、1×106、2×106、4×106、8×106cells/mL)中进行预培养,所用的容器为50mL烧杯,环境温度为(20±1)℃,光照度为4000lx,L∶D=1∶1。预培养期间,使轮虫始终处于指数增长期,培养时间为1个星期以上。挑龄期小于4h且活力充足的幼体进行试验。
1.2试验方法
1.2.1种群数量的测定采用群体累积培养方法:通过预培养获得处于指数增长期的轮虫,按试验浓度将轮虫接种到6个不同浓度的六孔板中。试验变量为小球藻的浓度,分别为0.25×106、0.5×106、1×106、2×106、4×106、8×106cells/mL,依次记为处理1、处理2、处理3、处理4、处理5、处理6。每组设6个平行,接种密度为1ind/mL,每孔容积为5mL,各试验条件与预培养条件一样。试验期间,每24h换1次轮虫培养液,直至轮虫种群密度开始下降,试验结束。记录每天的种群数量。
1.2.2生殖前期、子代数及寿命的测定采用单个体培养方法:通过预培养获得处于指数增长期的轮虫,按试验浓度将轮虫接种到6个不同浓度的24孔板中,小球藻浓度分别为0.25×106、0.5×106、1×106、2×106、4×106、8×106cells/mL,依次记为处理1、处理2、处理3、处理4、处理5、处理6。每组设12个平行,接种密度为1ind/mL,每孔容积为1mL,其他试验条件均与预培养条件一样。试验期间,每24h换1次轮虫培养液,记录第1次产卵的时间,每天把新生幼體挑除,直至母体死亡,试验结束。记录总共所产的后代数及一共所经历的时间。
1.3种群增长指标及测定方法
培养结束后,通过所得数据,可以计算出种群增长率(populationgrowthrate,r)。
[JZ]种群增长率(r)=(lnNt-lnN1)/t。
式中:t代表试验时间,d;N1和Nt分别表示1d和td雌体的数目。
生殖前期指幼体到产生第1枚卵所经历的时间。后代数指一个轮虫一生所产的后代数。寿命表示从出生到死亡所经历的时间,h。
1.4数据处理
应用软件sigmaplot12.5作图,SPSS22对试验数据进行t检验。
2结果与分析
2.12个品系褶皱臂尾轮虫种群动态变化
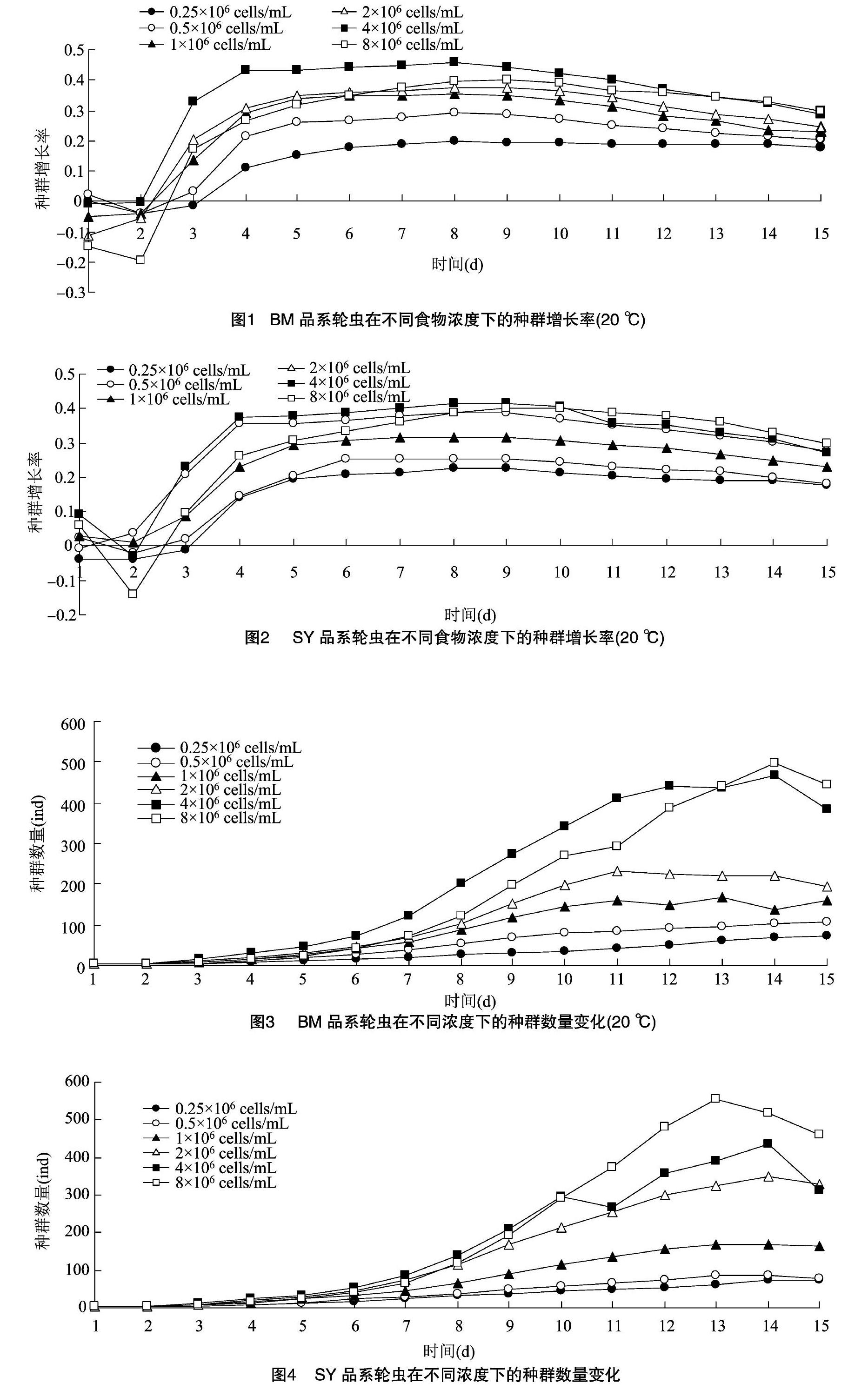
由图1和图2可知,2个品系褶皱臂尾轮虫在不同浓度下,种群增长率均随时间延长呈先快速增长后缓慢下降的趋势。单因素方差分析表明,6d之前,BM品系处理6的种群增长率低于处理4,且显著低于处理5的种群增长率(P<0.05,差异显著性分析未列出,下同);13d后,处理6的种群增长率才超过处理5。8d以前,SY品系处理6的种群增长率低于处理5,10d后,处理6的种群增长率才超过处理5。除了处理6的食物浓度下,BM和SY品系的种群增长率差异不显著外(P>0.05),其他食物浓度下SY品系的种群增长率显著高于SM品系。
2.22个品系褶皱臂尾轮虫种群数量变化
图3和图4为2个品系轮虫在不同浓度下的种群动态变化。在5mL容量的孔板中,2个品系轮虫的种群变化在不同浓度下均先增长后下降。在不同食物浓度下,褶皱臂尾轮虫所到达种群高峰所需要的时间不一样,BM品系到达峰值所需时间随食
物浓度的升高先减少后增加,处理4所需时间最短,只需11d。各食物浓度下,SY品系到达峰值所需时间差不多。2个品系种群增长率的最高值均出现在处理5。单因素方差分析表明,6~13dBM品系处理5的种群数量要显著高于其他浓度,到14d后处理6的种群数量才超过处理5,与处理5的种群数量差异不显著。SY品系在3~10d时,处理5的种群数量要高于其他浓度的种群数量,与处理6不显著;11d后,处理6要显著高于其他浓度。
2.32个品系褶皱臂尾轮虫生活史特征的变化
由表1可知,在20℃时,BM品系的生殖前期随食物浓度的升高呈缩短的趋势,在5mL容量的孔板中,处理1、处理2、处理3之间没有显著差异,处理5和处理6之间差异显著。SY品系的生殖前期随食物浓度的升高呈波浪变化,总体呈缩短趋势,2个品系都是处理1所需的生殖前期最长,BM品系在处理6所需生殖前期最短,而SY品系生殖前期最短所需浓度为处理5。BM品系的总子代数随食物浓度的升高呈上升的趋势,SY品系的总子代数呈先下降后上升的趋势。2个品系分别在处理5和处理6时的总子代数最多,BM品系在0.25×106个/mL时总子代数最少,而SY品系子代数最少时所需浓度为1×106cells/mL。BM品系轮虫的寿命随食物浓度的升高先延长后缩短,在处理5处寿命最长;SY品系寿命不断延长,在处理5和处理6处寿命最长。在中低浓度时(处理1、处理2、处理3和处理4),SY品系的寿命要低于BM品系,在高浓度时(处理5、处理6)SY品系的寿命要高于BM品系。
3讨论与结论
3.1BM品系和SY品系种群增长率和种群数量的变化
已有多数研究显示,食物浓度、品系和温度等因素都对萼花臂尾轮虫、壶状臂尾轮虫有显著影响。席贻龙等对壶状臂尾轮虫的研究说明了高温会对壶状臂尾轮虫的发育产生负面影响,缩短轮虫寿命[8]。本研究发现,在不同食物浓度下,2种轮虫的种群增长率均呈先快速增长后缓慢下降的趋势,种群变化也是先增长后下降,这与耿红等在红臂尾轮虫的研究所得的结果[9]一致。胡好远等对不同品系的角突臂尾轮虫进行了研究,发现不同品系角突臂尾轮虫之间的生活史特征存在着显著差异[10]。本试验通过研究2种国内外的轮虫,发现2种轮虫的种群增长率并没有显著差异,这与之前的研究有所不同。BM品系在处理6浓度处,13d后的种群增长率要大于处理5,这有可能是因为BM品系在13d前,处理5的浓度最适合BM品系生长;到了13d后,由于种群密度变大,处理6更能适合高密度BM品系的生长。而在SY品系中,10d前处理5的种群增长率要高于处理6,说明在初期高食物浓度对SY品系轮虫产生了抑制作用;10d后,处理6处的种群增长率才开始慢慢超过处理5。2种品系在前期都受到高食物浓度的抑制,从2个品系种群变化可以看出,SY品系在前期高浓度处理的种群数量要高于其他浓度,BM品系在前期其他浓度处理的种群数量要低于处理5,说明了高食物浓度对BM品系的抑制要大于SY品系。姚久祥等认为,在前期低浓度会促进轮虫的生长,食物浓度过高,会产生毒素,抑制轮虫生长[11],本研究所得到的结论与之基本一致。
3.2BM品系和SY品系生殖前期、总子代数和寿命的比较
BM品系的生殖前期随食物浓度的升高而缩短,食物浓度低于处理4时,生殖前期没有显著差异,这与席贻龙等在壶状臂尾轮虫所得到的食物浓度低于1.5×106cells/mL对轮虫的生殖前期无显著影响的结论[8]相一致;SY品系虽然也是呈缩短的趋势,但是变化并没有BM品系那么大,只有处理1与处理5的生殖前期存在显著差异,其他浓度并无显著差异。
SY的子代数随浓度的升高呈上升的趋势;BM的总子代数随浓度的升高先上升后下降,在处理5处达到了最多,这说明BM的最适生长浓度有可能在处理5处,SY品系要比BM品系更能适应高浓度。
2个品系在处理5处的寿命要显著长于低浓度,SY在处理6处与处理5处寿命相比基本保持不变,BM品系相比明显下降,表明BM品系在处理5处的生长发育要更好,更适合BM生长。董丽丽等对3个品系的萼花臂尾轮虫进行了研究,发现平均寿命随食物浓度升高先增加后减小,在3×106cells/mL处,平均寿命最大[12],与本研究BM品系褶皺臂尾轮虫所得到的结论相一致。但与SY品系的结论有所差别,有可能由于不同地理差异所产生不同的影响。BM品系和SY品系在生活史特征上有诸多异同,说明地理位置对褶皱臂尾轮虫的生活史有一定影响,这与吕林兰等所得到的不同品系萼花臂尾轮虫的发育和种群增长存在显著差异[13]相一致。
BM品系在处理6处的生殖前期和寿命要比处理5处短,子代数要少于处理5;SY品系在处理6处的生殖前期要比处理5长,BM品系种群数量最高值出现在处理5。综合本研究可知,在20℃下,国内SY品系要比美国BM品系对高浓度海水小球藻的耐受性强,在低于处理5的食物浓度下,BM品系适合大规模培养,高于处理5浓度时,SY品系更适合大规模培养。本研究只对海水小球藻不同浓度进行了试验,李磊等研究了不同的微藻对褶皱臂尾轮虫的影响,认为轮虫的摄食率与饵料类型有关[14]。有人认为,轮虫依靠纤毛滤食,所以食物细胞的大小和密度影响轮虫的摄食情况[15]。也有研究说明,由于藻类细胞大小不一,通过几种藻类的换用能够更好地促进轮虫的生长[16]。轮虫食物浓度低,繁殖率降低,寿命缩短,这与Yoshinaga等得到的结论[17]相一致。
参考文献:
[1]葛雅丽,席贻龙,陈芳,等.食物浓度和品系对萼花臂尾轮虫生殖期历时和混交雌体形成的影响[J].生态学报,2005,25(8):3-9.
[2]Pérez-legaspiIA,Rico-martínezR.Effectoftemperatureandfoodconcentrationintwospeciesoflittoralrotifers[J].Hydrobiologia,1998,387/388:341-348.
[3]冯粒克,席贻龙.三个地理品系萼花臂尾轮虫生活史特征的比较[J].动物学杂志,2004,39(1):12-15.
[4]XiangX,XiY,WenX,etal.PatternsandprocessesinthegeneticdifferentiationoftheBrachionuscalycifloruscomplex,apassivelydispersingfreshwaterzooplankton[J].MolecularPhylogeneticsandEvolution,2011,59(2):386-398.
[5]Rehberg-HaasS,MeyerS,LippemeierS,etal.AcomparisonamongdifferentPavlovasp.productsforcultivationofBrachionusplicatilis[J].Aquaculture,2015,435:424-430.
[6]ChengSH,KaS,KumarR,etal.Effectsofsalinity,foodlevel,andthepresenceofmicrocrustaceanzooplanktersonthepopulationdynamicsofrotiferBrachionusrotundiformis[J].Hydrobiologia,2011,666(1):289-299.
[7]冯大庆.人工海水的简易制法[J].生物学通报,1986(10):37-39.[HJ2.14mm]
[8]席贻龙,黄祥飞.温度对壶状臂尾轮虫实验种群动态的影响[J].海洋与湖沼,2000,31(1):23-28.
[9]耿红,席贻龙,胡好远.食物种类和浓度对红臂尾轮虫种群增长、个体大小及卵大小的影响[J].应用生态学报,2003,14(5):753-756.
[10]胡好远,席贻龙,耿红.三个品系角突臂尾轮虫生长和发育的比较研究[J].应用生态学报,2003,14(4):565-568.
[11]姚久祥,彭敏.藻类食物对轮虫群体增殖的影响[J].广西水产科技,2001(4):21-25.
[12]董丽丽,席贻龙,刘桂云,等.温度和食物浓度对三品系萼花臂尾轮虫实验种群动态的影响[J].应用生态学报,2004,15(11):2165-2169.
[13]吕林兰,董学兴,董斯明,等.两品系萼花臂尾轮虫的生活史特征和受精率研究[J].大连海洋大学学报,2012,27(5):401-410.
[14]李磊,朱小明,吴洪喜.营养强化时褶皱臂尾轮虫对饵料微藻的摄食[J].生态学杂志,2011(9):2019-2025.
[15]RothhauptKO.Differencesinparticlesize-dependentfeedingefficienciesofcloselyrelatedrotiferspecies[J].LimnologyandOceanography,1990,35(1):16-23.
[16]ViayehRM,MohammadiH,ShafieiAB.PopulationgrowthofsixIranianBrachionusrotiferstrainsinresponsetosalinityandfoodtype[J].InternationaleRevuederGesamtenHydrobiologieundHydrographie,2010,95(6):461-470.
[17]YoshinagaT,HagiwaraA,TsukamotoK.Lifehistoryresponseandage-specifictolerancetostarvationinBrachionusplicatilisofMuller(Rotifera)[J].JExpMarBiolEcol,2003,287(2):261-271.
猜你喜欢
贵州畜牧兽医(2023年3期)2023-06-29
农技服务(2023年2期)2023-03-15
中老年保健(2021年8期)2021-12-02
作文评点报·低幼版(2020年3期)2020-02-12
园林科技(2020年2期)2020-01-18
华人时刊(2018年17期)2018-12-07
数学年刊A辑(中文版)(2015年3期)2015-10-30
数学年刊A辑(中文版)(2015年3期)2015-10-30
海南热带海洋学院学报(2014年2期)2014-08-08
四川轻化工大学学报(自然科学版)(2014年3期)2014-04-16
