
GlgC基因种子特异过表达载体的构建及对甘蓝型油菜的转化
2020-04-23 10:09赵凯琴罗延青王敬乔李劲峰张云云杨清辉
西南农业学报 2020年2期
赵凯琴,罗延青,俎 峰,王敬乔*,李劲峰,陈 苇,张云云,杨清辉
(1.云南农业大学农学与生物技术学院,云南 昆明 650201;2.云南省农业科学院经济作物研究所,云南 昆明 650205)
【研究意义】油菜(BrassicanapusL.)是世界上食用植物油的主要来源之一。提高油菜单位面积产量或产油量一直是中国油菜育种重点研究课题。提高油菜品种含油量是提高油菜产油量的最有效方式[1],是提高油菜生产效益的关键[2]和油菜育种的主要目标[3]。通过GlgC基因在油菜胚发育前期、中后期表达,以期增加淀粉积累及油脂合成前体物质(果糖)的含量,提高前期胚的生长速度;同时为中、后期胚发育提供最直接的碳源,达到增加种子含油量的目的。【前人研究进展】 AGPase(腺苷二磷酸葡萄糖焦磷酸化酶)是淀粉合成途径的关键限速酶,其催化淀粉合成前体腺苷二磷酸葡萄糖的产生。GlgC是同源四聚体,单基因编码AGPase。AGPase在细菌和植物及糖原合成中起相似作用[4]。Stark等[5]的研究证明,来源于细菌的AGPase基因GlgC在马铃薯球茎中过表达可增加淀粉含量30 %。Sweetlove等[6]通过抑制马铃薯AGPase 的活性导致淀粉合成的部分或全部终止。Helence[7]等利用胚特异启动子降低了AGPase在转基因油菜中的表达;通过同位素示踪分析显示,AGPase活性下降导致淀粉合成效率下降50 %~60 %,淀粉含量下降接近50 %~70 %;同时,通过对种子发育各个时期油含量的分析,AGPase活性下降导致了含油量的下降,特别是在胚发育中期以前,转基因株系胚含油量比野生型下降50 %以上。目的基因的组织特异性及时空表达对最终产物的作用效果影响极大,如利用组成型的35S启动子和维管特异启动子4CL分别过表达蔗糖合酶基因SuSy,前者使杨树茎干重下降,后者使茎干重上升[8]。因此,启动子的选择能直接影响预期结果。【本研究切入点】构建了两个含特异表达启动子的GlgC基因种子特异过表达载体,以农杆菌介导法导入甘蓝型油菜。【拟解决的关键问题】获得甘蓝型油菜转GlgC基因阳性株,为利用遗传改良技术实现上述途径在油菜基因工程育种中的应用提供材料基础。
1 材料与方法
1.1 材料
1.1.1 受体 甘蓝型油菜BrassicanapusL. ‘ westar’(加拿大引进)。
1.1.2 菌种 大肠杆菌(Escherichiacoli)DH5α感受态及根癌农杆菌(Agrobacteriumtumefaciens) LB4404感受态均购于Takara 大连宝生物工程有限公司。
1.1.3 载体 克隆载体pMD18-T Vector 购于Takara 大连宝生物工程有限公司;载体pMB-TG、pMB3-14、pMB-Fad2为本实验室保存及构建。
1.1.4 分子试剂及化学试剂 所有酶、核酸分子量标准DNA Marker 购于Takara 大连宝生物工程有限公司;化学试剂均为国产分析纯。
1.2 方法
1.2.1 拟南芥胚特异启动子DOF的克隆(At4g21080) 根据Genbank 中已公布的拟南芥早期胚特异启动子At4g21080(DOF4.5)的核苷酸序列设计引物(表1),以野生型拟南芥基因组DNA为模板,扩增985 bp启动子片段。PCR程序: 94 ℃,5 min;94 ℃,30 s,55 ℃,1 min,72 ℃,1 min 30 s,35个循环;72 ℃,10 min。将PCR产物连接到PMD18T上,载体命名为pMD18T-DOF。
1.2.2GlgC特异启动子过表达载体的构建 拟南芥胚特异启动子DOF+ GlgC突变基因表达载体的构建:①载体pMB-TG上有2个XbaI位点,为下一步的重组克隆要先消除3′端的XbaI位点。用XbaI部分酶切pMB-TG,klenow补平,自连,转化大肠杆菌DH5自连挑菌落提取质粒进行XbaI单酶切和XbaI+KpnI双酶切鉴定,酶切结果应为线性和-330 bp片段。将消除3′端的XbaI位点的pMB-TG命名为pMB-TG-XbaI。②连接头:XbaI单酶切pMB-TG-XbaI,klenow补平,胶回收,去磷酸化,连接PXhoI,转化大肠杆菌DH5肠杆挑菌落提取质粒进行XhoI+KpnI双酶切鉴定(-330 bp)。将消除5′端XbaI并连上PXhoI的pMB-TG-XbaI命名为pMB-TG-XhoI。③用SmaI+SalI双酶切pMD18T-DOF,电泳回收~1.0 kB的拟南芥早期胚特异启动子At4g21080(DOF4.5)片段插入到经PmeI+XhoI双酶切的pMB-TG-XhoI并去磷酸化的开环载体上(SmaI和PmeI为平末端;SalI和XhoI为同链末端),转化后鉴定的阳性正向克隆送测序。
种子特异蛋白启动子Cruc+GlgC突变基因表达载体的构建:将实验室已构建的含Cruc启动子的pMB3-14进行SalI酶切,切下Cruc启动子片段,连接到经过相同酶切,回收载体大片段的pMB-Fad2载体上,命名为pMB-Cruc。XbaI单酶切含~1.6 kBGlgC片段的pMD18T-TG,将目的片段连接到经XbaI 酶切的pMB-Cruc上,转化大肠杆菌DH5α,鉴定后的阳性正向克隆送测序。
1.2.3GlgC基因表达载体的甘蓝型油菜遗传转化GlgC基因表达载体的农杆菌转化:将重组质粒pMB-DOF-TG、pMB-Cruc-TG导入根癌农杆菌LBA4404(采用冻融法)。
农杆菌介导法转化油菜及抗Kan再生植株筛选:①将转化受体‘Wester’种子用25 %次氯酸钠灭菌,在MS培养基中25 ℃暗培养7 d;②挑取农杆菌单克隆,置于相应抗性的10 mL YEP液体培养基,27 ℃震荡培养(200 r/min)至对数生长期,OD 值0.3备用;③取上述已制备好的农杆菌吸取0.5 mL加入20 mL MS培养基,将幼苗下胚轴切成0.5~1.0 cm 的小段浸入60 min,将下胚轴用滤纸吸干后转入共培养基培养3 d,25 ℃;④转入诱导培养基中光照培养14 d;其后在分化培养基中15 d继代1次,至分化幼苗;再转入筛选培养5~7 d继代1次;待幼苗2~3 cm时转入生根培养(培养基配方参考文献[9])。
1.2.4 转化植株的分子生物学鉴定 PCR检测:PCR直接扩增,以转基因kan阳性植株叶片为模板。引物为GlgC及NP TⅡ(表1)。PCR程序:98 ℃2 min;98 ℃10 s,60 ℃15 s,68 ℃1 min,40 个循环;68 ℃10 min。PCR产物进行琼脂糖凝胶水平电泳。
半定量RT-PCR检测:取经PCR检测为阳性的转基因植株叶片100 mg,提取总mRNA。按PrimeScript RT reagent Kit with gDNA Eraser说明书合成 cDNA。内参基因为Brasicanapusactin,扩增引物为BnAc;目的片段AlgC扩增引物为GlgCsq(表1)。PCR程序:94 ℃ 5 min;94 ℃ 40 s,58 ℃ 1 min,72 ℃ 1 min,34个循环;72 ℃ 延伸10 min。PCR产物进行琼脂糖凝胶水平电泳。

表1 本文所涉及的引物序列

图1 DOF克隆序列比对

1: M. DL5000 bp;2~5: pMD18T-DOF用EcoRI+SalI酶切1: M DL5000; 2-5: pMD18T-DOF digested by EcoRI+SalI
2 结果与分析
2.1 DOF启动子的克隆及序列分析
对pMD18T-DOF阳性克隆菌液测序,结果显示成功获得DOF基因的克隆(图1);对pMD18T-DOF质粒进行EcoRI+SalI双酶切,琼脂糖电泳得到1025 bp条带,成功获得连有DOF启动子的pMD18T-DOF载体(图2)。
2.2 特异启动子GlgC表达载体构建
2.2.1 拟南芥胚特异启动子DOF(At4g21080)+GlgC突变基因表达载体的构建 对转化后鉴定的阳性正向克隆进行测序,测序验证目标序列与pMB-TG-XhoI(去磷酸化)载体融合表达的读码框正确。结果显示成功得到重组质粒pMB-DOF-TG(构建流程见图3)。
2.2.2 种子特异蛋白启动子Cruc+GlgC突变基因表达载体的构建 重组载体经SacI+XbaI双酶切验证及测序比对,结果显示成功构建pMB-Cruc-TG载体(图4)。
2.3 GlgC特异过表达载体在甘蓝型油菜中的转化
通过农杆菌介导将GlgC基因转入甘蓝型油菜中,受体材料为油菜下胚轴,经再生、分化及筛选,共获得甘蓝型油菜kan阳性植株228 株。
2.4 转化植株的分子生物学鉴定
2.4.1 转化植株的PCR鉴定 以kan阳性株DNA为模板,PCR产物经琼脂糖凝胶水平电泳显示获得750 bp NPTII及1300 bpGlgC目标条带。228 株kan抗性转化株中有202 株为PCR阳性株(图5~6)。
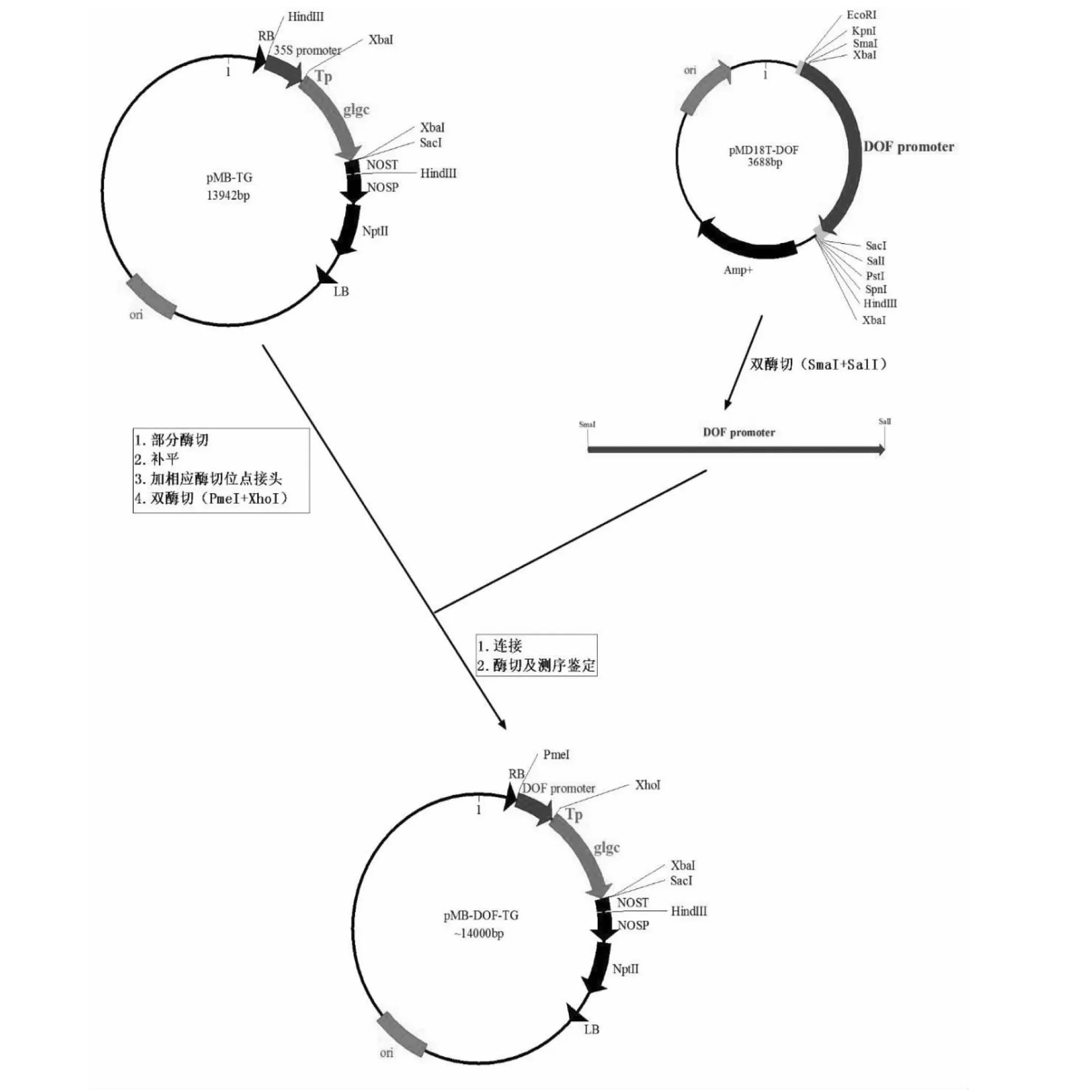
图3 种子特异过表达载体pMB-DOF-TG构建流程
2.4.2GlgC基因的RT-PCR表达与分析 结果显示,外源导入的GlgC基因在PCR阳性化株内能够表达。图7为部分RT-PCR产物琼脂糖凝胶电泳结果。
3 讨 论
在油菜种子中淀粉与油脂都在质体中进行合成,淀粉的降解可能为油脂的快速合成提供最近的碳源[10]。淀粉合成与环境条件、营养供应密切相关,淀粉在植物生长速度较慢时(如营养物质缺乏,低温等条件下)积累[11];低温和长日照均有利于淀粉的合成与积累[12]。同样,对油菜和拟南芥的研究显示环境条件对种子油含量也存在相当大的影响[13]。栾运芳等[14]的研究表明,在全国范围内,西藏种植的油菜含油量最高,与西藏地区昼夜温差大、日照长的环境条件密切相关。付三雄[15]在对南京(海拔8.9 m)和拉萨(海拔3652 m)2个不同海拔地区种植的甘蓝型油菜高油品系H105的研究中发现,该材料含油量在两地分别为46.04 %±1.42 %和53.09 %±1.35 %,显示同一品种在西藏种植时含油量更高。这一结果间接表明了淀粉与油脂合成间存在正相关关系。
另外,目的基因的组织特异性及时空表达对最终产物的作用效果影响极大,在过表达研究中,启动子的选择能直接影响预期结果。
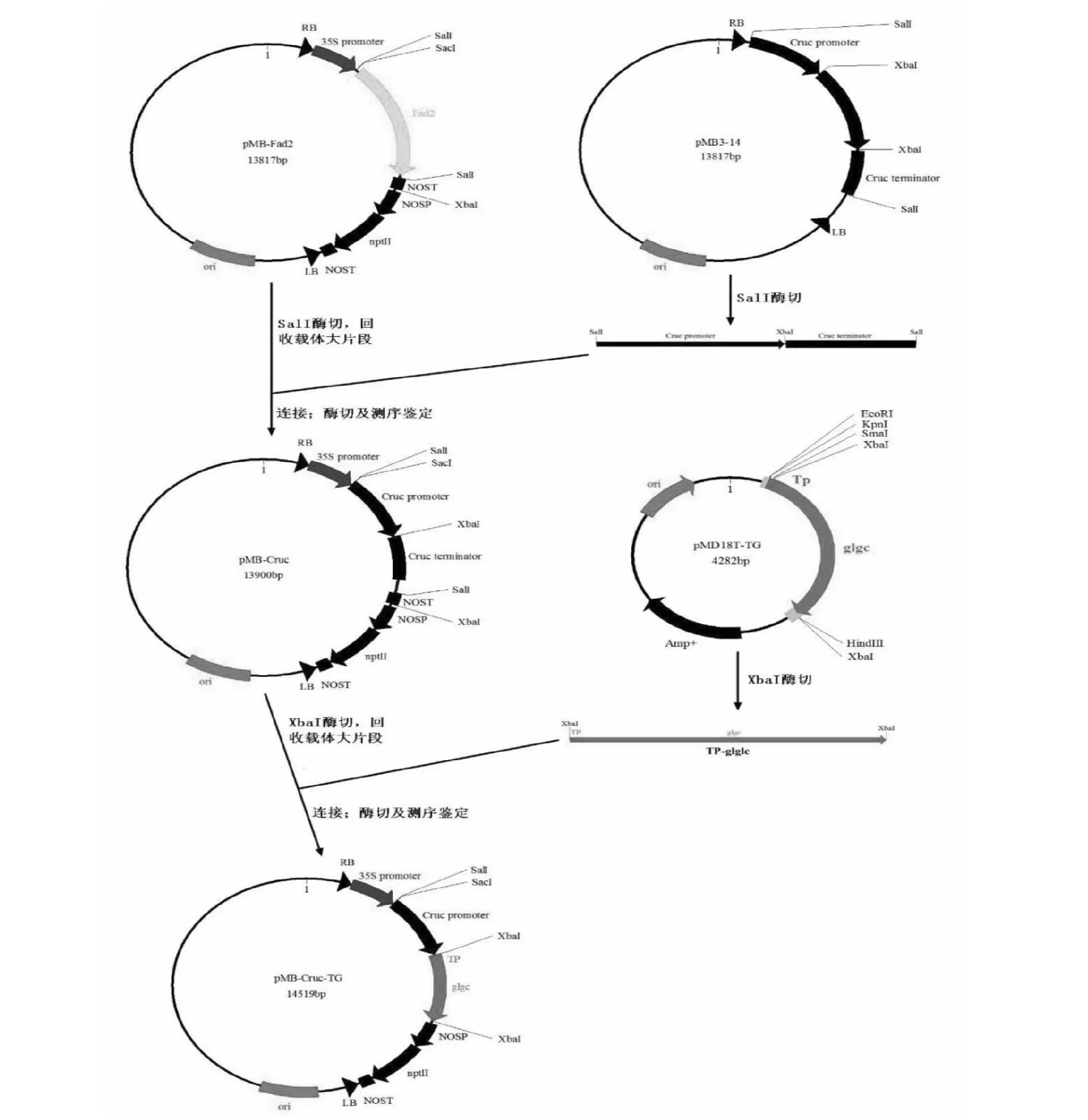
图4 种子特异过表达载体pMB-Cruc-TG构建流程

1: DL15000(M);2: 阴性对照(非转基因植株);3: 阳性对照(pMB-DOF-TG);4~25: 转基因植株1: DL15000(M); 2: Negative control(non-transgenic plants); 3: Positive control(pMB-DOF-TG); 4-25: Transgenic plants

1:DL5000(M);2:阳性对照(pMB-Cruc-TG);3:阴性对照(非转基因植株);4~25:转基因植株1: DL5000(M);2: positive control(pMB-Cruc-TG); 3: Negative control(non-transgenic plants); 4-25: Transgenic plants

1:阴性对照;2:阳性对照;3~12:转基因植株1: Transgenic lines; 2: Positive control; 3-12: Negative control
4 结 论
笔者在前期研究中克隆了淀粉合成途径的关键酶AGPase的大肠杆菌同源基因GlgC并进行了突变,获得突变体GlgC16基因,降低对无机磷酸盐敏感性,并进行了质体定位[8]。本研究在油菜AGPase小亚基导肽基因片段与上述GlgC突变基因构建融合基因的基础上,将GlgC突变基因分别和两个特异表达启动子DOF(拟南芥早期胚(球形胚阶段)启动子)、Cruc(油菜种子蛋白启动子)重组,构建了2个GlgC过表达载体pMB-DOF-TG和pMB-Cruc-TG,转化农杆菌LBA4404。通过油菜农杆菌转化体系进行转化,并验证了外源导入的GlgC基因在阳性转基因油菜植株内获得表达,获得了甘蓝型油菜转GlgC基因阳性株,为利用遗传改良技术实现上述途径在油菜育种中的应用奠定了前期基础。期望通过GlgC基因的过表达,增加淀粉含量,为胚中油脂的快速合成提供最直接的碳源,达到增加含油量的目的。
猜你喜欢
学与玩(2022年10期)2022-11-23
作物学报(2022年12期)2022-10-14
今日农业(2022年3期)2022-06-05
作物学报(2022年4期)2022-02-10
作物学报(2022年4期)2022-02-10
作物学报(2022年3期)2022-01-22
园艺与种苗(2020年12期)2021-01-08
创新科技(2015年1期)2015-12-24
飞碟探索(2015年9期)2015-11-05
红领巾·探索(2015年9期)2015-09-10
