
天童山阔叶林不同树种各器官非结构性碳水化合物储量及分配
2020-04-23 10:10鞠鹏杰黄长兵
西南农业学报 2020年2期
鞠鹏杰,黄长兵
(苏州农业职业技术学院,江苏 苏州 215000)
【研究意义】对于植物碳水化合物而言,其能够根据结构特性分为结构性、非结构性两种,后者作为短期的代谢活动库在植物生长过程中具有重要作用[1-2],也就是常说的NSC,其不仅能够以淀粉的形式存在,同时能够以可溶性糖分的形式进行存储,这是光合产物的一种暂时储存形式,对于植被生长发育至关重要,参与其生命过程[3],此外,这种物质能够提升植被的抗旱性能,当光合作用产生的能量供应不足时,其能够作为一种缓冲物质提供相应的碳源,从而保障植被的正常新陈代谢,进而保障植被的生长发育;此外,其具有明显的渗透调节作用,能够有效调节相应的膨压,在细胞吸水方面作用显著;与此同时,其在促进水分传导方面的效果依然较为突出[4-5]。对于林木而言,其树干中形成了一定的NSC储量,这些物质存储能够有效提供能量缓冲,成为林木正常生长发育的保障物质之一,同时这也是碳收支的反映,体现着碳循环的动态变化。因此,如何评估树干的NSC储量成为分析树木碳循环的一个重要方面,也能够对树木生长过程中的环境胁迫开展相关研究,对于深化树木新陈代谢活动研究起着有益作用,更有利于碳循环模型构建[6-7]。对于整个树木而言,其生物量最主要的部分在于树干,这也是NSC主要所在,因此为了进一步开展NSC方面的研究,必须深入探讨树干的NSC分布,尤其是纵向及横向的对比,这样不仅能够对林木的生长特性开展相关研究,同时对比碳循环的变化,提升存储量估算准确性[8]。对于成年树干而言,从横断面的角度来看,其木质部分不仅有诸多边材构成,更有心材这一核心部位[9-10];边材的主要作用在于进行液流的输送,同时对养分进行相应的存储,对水分进行相应的分配及传输[11],同时将碳水化合物进行储存以供林木生长所需;对于新材来说,其体内能够将树脂及胶质等物质进行有效的聚集;心材和边材的作用具有较大差异,这是树木生理特征的重要表现之一,是树木的核心部位。【前人研究进展】学者张海燕[12]通过对白桦树进行研究发现,其与紫椴树干的NSC存储能量具有较大差异,差异超过10 %,尤其是树干1.3 m处木质外侧。学者成方妍[13]通过实地对比分析得知,无论是林分还是个体,树皮有利于强化树干的NSC存储,其贡献率接近30%,但是多数研究并未对树皮在NSC存储方面加以研究分析。【本研究切入点】不同的树种具有明显差异的NSC存储能力,而且其空间分布也会产生较大的影响,对相应的存储估算产生明显的不确定性[9]。【拟解决的关键问题】基于此,本研究将不同阔叶林树种作为研究对象,从树皮、边材及心材等角度来探讨其NSC存储方面的差异,为了提升实验准确性,本研究进行了连续3年的实地研究,同时对存储能量与树干高度方面的关系进行了探究,并对各树种间的NSC存储误差进行了对比,从而深入了解林木物质能量循环,进一步了解其碳循环和能量交换,为林木保护提供有益借鉴和参考。
1 材料与方法
1.1 研究区概况
本实验选择在浙江宁波的天童山进行,该区域位于宁波东南部,受所处经纬度的影响,该区域的亚热带季风特点较为明显,总体来说具有较丰沛的降雨,根据近年来气象资料统计得知,其年均降雨量接近1500 mm,冬季气温较低且较为干旱;该区域全年平均气温为17 ℃,夏季气温较高且多雨。该研究区域具有较为复杂的地形,北高东低的特点使得该区域存在2条较大的山脊、沟谷,其土壤类型以黄红壤为主,但是其酸性较为明显,土层厚度分布差异明显。受地理位置制约,该区域分布了大量的常绿阔叶林,尤其是樟科、壳斗科和山茶科,人工林主要是杉木林、马尾松林和竹林,其间分布着较多灌丛。

表1 样地基本特征
1.2 样地布设
本实验开始于2015年,并进行连续3年的实地观测,具体地点位于天童森林观测站,选取的植被为木荷、青冈、烤树和石栎4种,各选取9株加以研究,首先记录其生长特征等,具体如表1所示。待其叶片完全凋落,采用生长锥进行树芯的采样,要求采样区域位于树干南向:首先在距离地面10 cm处的根颈进行一次采样,然后在距离地表1.3 m处采样,接下来在冠基及冠中的部位分别进行采样,每次进行3次采样,并把各个采样编号后于冷箱保存;在进行指标测定之前,需要在微波炉中进行长达1.5 min的杀青处理,以促进酶的变化,之后烘干称重,并进行粉碎。
对于NSC存储而言,其具有较为明显的时间分布差异,因此每次取样在14:00-17:00进行,地上部分分为树干、树枝、叶片,而根系以2 mm为界分为粗根及细根;将获取的样品进行冲洗后晾干,然后进行1.5 min杀青处理,避免酶反应对实验带来的误差,再在80 ℃下烘干,称重后进行粉碎,测定NSC含量。
1.3 室内分析
本研究中NSC包括可溶性糖和淀粉,其含量通过苯酚硫酸改进法进行测定,对于NSC而言,其超过90 %的成分为可溶性糖及淀粉,本研究将其称为总非结构性碳水化合物[9-11]。
利用光合仪开展光合速率的测定,同时对气孔导度进行测量,测定时间选在8:00-18:00,所用光源为红蓝光,光合辐射、CO2浓度分别为1000、385 μmol m-2·s-1。
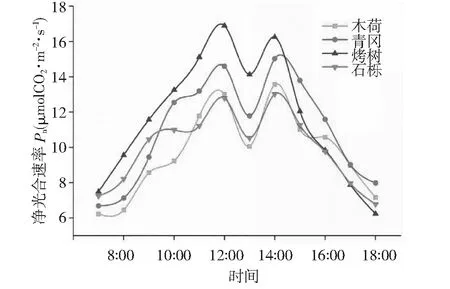
图1 天童山阔叶林不同树种叶片净光合速率日变化趋势
1.4 数据处理
所有数据均以平均值±标准误差显示;采用双因素方差分析(Two-way ANOVA)差异显著性,Origin 9.0作图。
2 结果与分析
2.1 天童山阔叶林不同树种叶片净光合速率
由图1可知,天童山阔叶林不同树种叶片光合速率在一天之内呈“M”型变化趋势,在13:00出现降低,此时由于光照过强,导致叶片气孔关闭,因此13:00出现降低。对于木荷,光合速率变化范围在6.23~13.56 μmol CO2m-2·s-1,青冈光合速率变化范围在6.69~15.02 μmol CO2m-2·s-1,烤树光合速率变化范围在7.49~16.89 μmol CO2m-2·s-1,石栎光合速率变化范围在7.25~13.026 μmol CO2m-2·s-1,相同时刻光合速率大小基本表现为烤树>青冈>石栎>木荷。由图2可知,天童山阔叶林不同树种叶片光合速率在一年之内呈先增加后降低趋势,相同季节光合速率大小基本表现为青冈>烤树>石栎>木荷。
2.2 天童山阔叶林不同树种地上NSC储量
由表2可知,叶片NSC储量、可溶性糖和可溶性淀粉变化范围在5.01 %~6.59 %、3.12 %~6.59 %和1.58 %~2.26 %,其大小依次表现为烤树>青冈>石栎>木荷;枝条NSC储量、可溶性糖和可溶性淀粉变化范围在9.28 %~11.24 %、6.03 %~7.56 %和2.15 %~3.45 %,其大小依次表现为烤树>青冈>石栎>木荷;皮NSC储量、可溶性糖和可溶性淀粉变化范围在11.36 %~13.02 %、8.16 %~10.16 %和3.25 %~5.13 %,其大小依次表现为烤树>青冈>石栎>木荷;边材NSC储量、可溶性糖和可溶性淀粉变化范围在3.16 %~5.02 %、1.13 %~2.15 %和0.72 %~1.36 %之间,其大小依次表现为烤树>青冈>石栎>木荷;心材NSC储量、可溶性糖和可溶性淀粉变化范围在2.04 %~3.14 %、0.61 %~1.14 %和0.23 %~0.81 %,其大小依次表现为烤树>青冈>石栎>木荷;树干NSC储量、可溶性糖和可溶性淀粉变化范围在6.23 %~7.80 %、3.86 %~5.20 %和1.54 %~2.60 %,其大小依次表现为烤树>青冈>石栎>木荷。
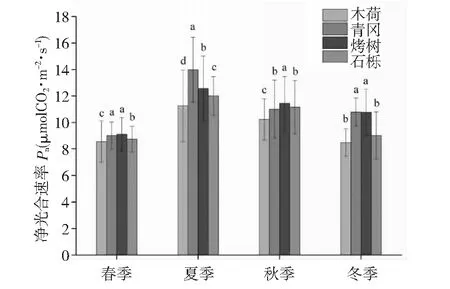
图2 天童山阔叶林不同树种叶片净光合速率季节变化趋势
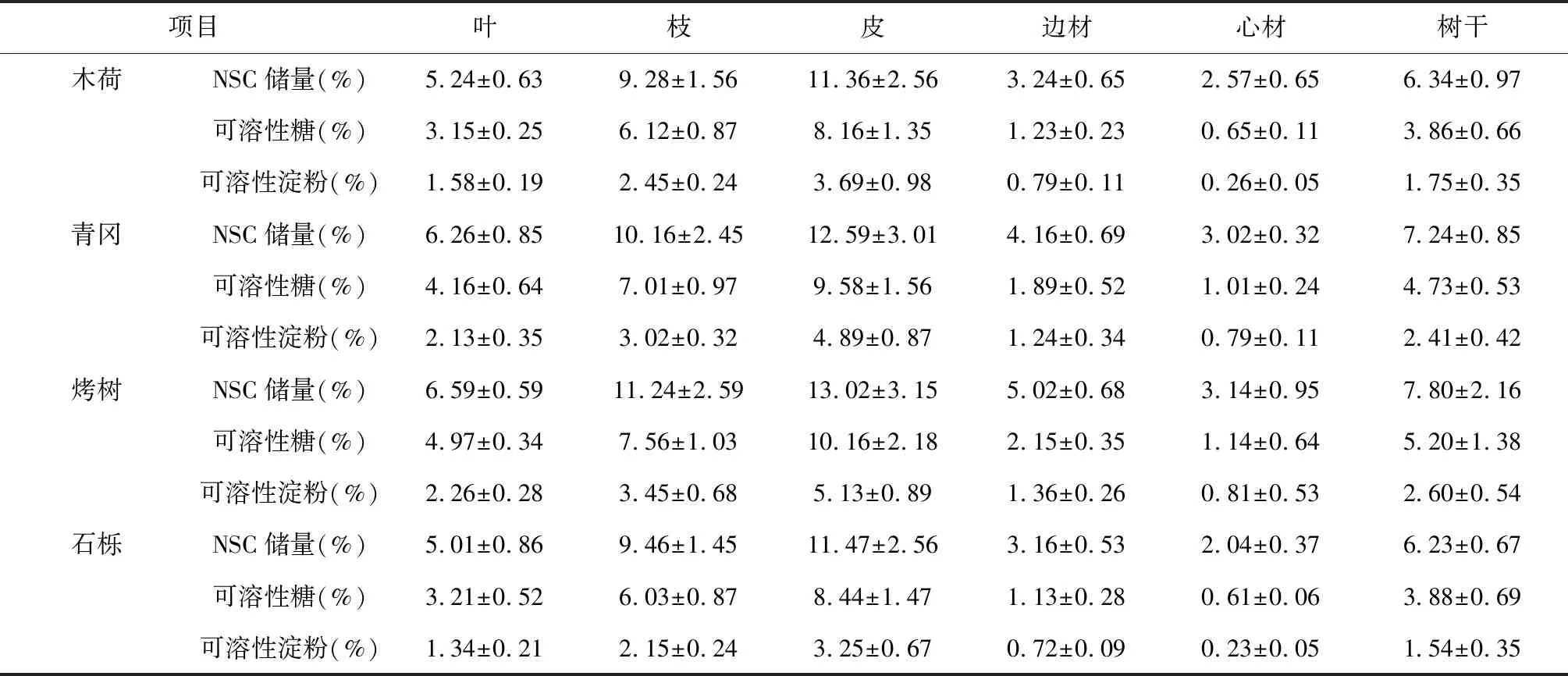
表2 天童山阔叶林不同树种地上NSC储量
2.3 天童山阔叶林不同树种地下NSC储量
由表3可知,粗根NSC储量、可溶性糖和可溶性淀粉变化范围在7.85 %~9.69 %、5.78 %~8.34 %和1.24 %~2.21 %,其大小依次表现为烤树>青冈>石栎>木荷;细根NSC储量、可溶性糖和可溶性淀粉变化范围在12.52 %~14.41 %、8.23 %~10.98 %和3.56 %~4.22 %,其大小依次表现为烤树>青冈>石栎>木荷;根系NSC储量、可溶性糖和可溶性淀粉变化范围在10.19 %~12.05 %、7.01 %~9.66 %和2.40 %~3.22 %,其大小依次表现为烤树>青冈>石栎>木荷。
2.4 天童山阔叶林不同树种NSC储量的高度变异
由图3可知,天童山阔叶林不同树种NSC储量与树干高度呈显著的指数关系(P<0.05),其中木荷、青冈、烤树和石栎指数关系R2分别为0.79、0.51、0.62和0.73;天童山阔叶林不同树种可溶性糖与树干高度呈显著的线性关系,其中木荷、青冈、烤树和石栎线性关系R2分别为0.75、0.62、0.76和0.81;天童山阔叶林不同树种可溶性淀粉与树干高度呈显著的线性关系,其中木荷、青冈、烤树和石栎线性关系R2分别为0.82、0.74、0.83和0.84。

表3 天童山阔叶林不同树种地下NSC储量
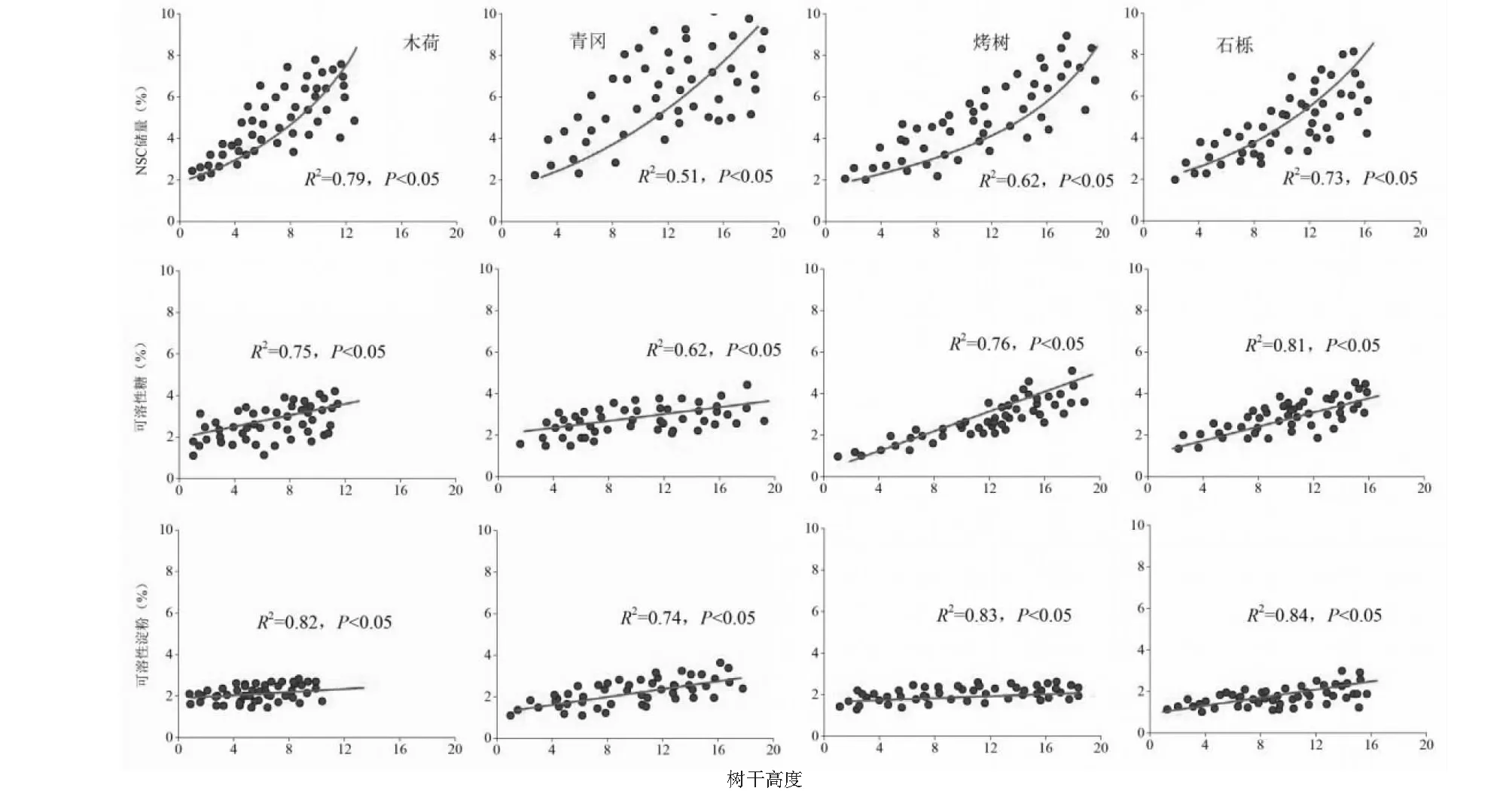
图3 天童阔叶林不同树种NSC储量的高度变异
2.5 NSC储量的估测误差
与利用4个高度的各段树芯所对应的圆环质量加权非结构性碳水化合物浓度(DL)方法估算整个树干NSC总量相比,叶、枝、皮、边材、粗根和细根NSC储量的相对误差的贡献率分别为26.3 %、21.6 %、21.3 %、29.8 %、19.8 %和21.4 %(图4),叶、枝、皮、边材、粗根和细根可溶性糖的相对误差的贡献率分别为42.1 %、38.9 %、45.3 %、35.9 %、39.8 %和37.8 %;叶、枝、皮、边材、粗根和细根可溶性淀粉的相对误差的贡献率分别为13.6 %、16.1 %、13.2 %、15.3 %、16.4 %和12.7 %。
2.6 光合速率对NSC的影响
由表4可知,木荷枝、皮、细根NSC储量与光合速率呈显著正相关;皮、细根和根系可溶性糖与光合速率呈显著正相关;枝、皮、粗根和细根可溶性淀粉与光合速率呈显著正相关。青冈枝、皮、粗根和细根NSC储量与光合速率呈显著正相关;叶、枝、皮、树干、粗根和细根可溶性糖与光合速率呈显著正相关;枝、皮、树干、粗根和细根可溶性淀粉与光合速率呈显著正相关。烤树枝、皮、树干、粗根、细根和根系NSC储量与光合速率呈显著正相关;皮、树干粗根、细根和根系可溶性糖与光合速率呈显著正相关;枝、皮、树干、粗根、细根和根系可溶性淀粉与光合速率呈显著正相关。石栎皮、粗根和细根NSC储量与光合速率呈显著正相关;皮、树干、细根和根系可溶性糖与光合速率呈显著正相关;皮、细根和根系可溶性淀粉与光合速率呈显著正相关。
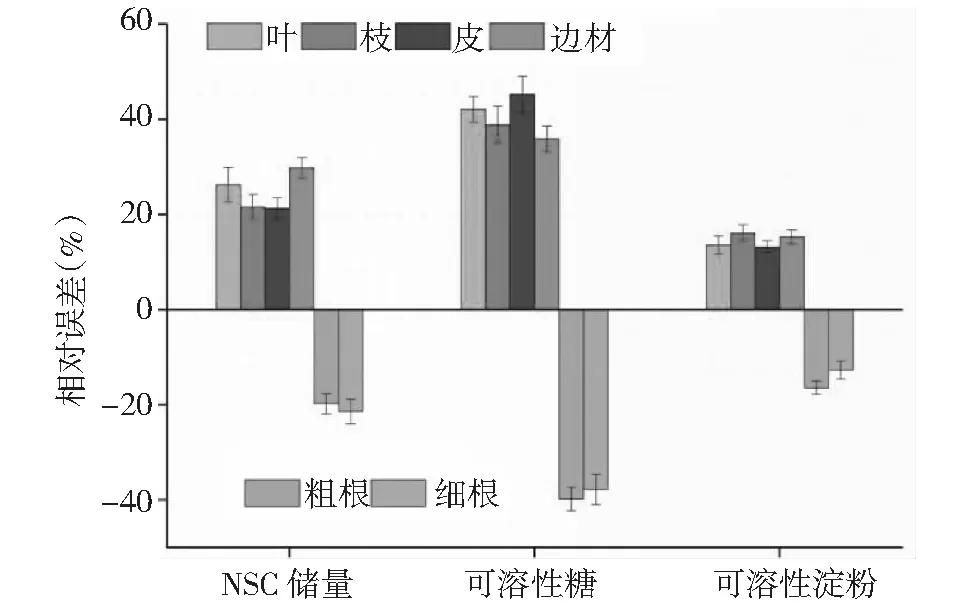
图4 NSC储量的估测误差
3 讨 论
对于青冈和烤树边材而言,其NSC存储能力明显高于木荷和石栎,与环孔及半环孔材树种相比,无孔材树种的NSC存储能力较弱[14-15],说明不同的树种具有不同的存储能力,这主要在于材性的差异,这种差异将影响到NSC的分配水平。韩风森等[16]通过对比研究得知,无梗花栎的储备碳库能够为木质生长提供55 %左右的碳含量,对于天童阔叶林碳库而言,其提供的碳含量约为41 %,明显较低,欧洲落叶松亦是如此[17],环孔材具有更高水平的NSC存储能力[17-18],这样能够为林木生长提供更为充足的碳能量来源。另外,对于环孔材的树种而言,其拥有较大的导管,有利于液流的传输,也利于碳成分的传送。
边材不仅对营养物质具有较高的存储能力,此外能够对水分等碳水化合物进行存储以备随时向树木提供能量[19],但对于芯材而言,其细胞活力明显较弱,其活性很低,能量存储能力较弱,因此能量存储主要在于边材方面[20]。通过本研究对比分析得知,对于木荷和石栎而言,其树皮的NSC存储能力明显弱于青冈和烤树,以往关于NSC存储方面的研究多在边材方面,但是树皮对其存储也有重要影响,也是NSC储库的一部分。对于青冈和烤树而言,其NSC成分中淀粉的含量超过了一半,这主要在于材性的差异,进而导致其碳储能力呈现较大差异。朱丽薇等[21]从碳存储方面进行分类,将树木碳存储分为“脂肪”型、“淀粉”型以及介于二者的中间型,对于环孔材而言,其多为“淀粉”型;对于树皮NSC存储而言,其储量是边材的3倍左右,是心材的,5倍以上,这种径向的变化较为明显,这基本与以往学者研究一致[18-20]。NSC通过树木导管等进行传输,在此过程中部门能量以淀粉的形式被薄壁细胞吸收,作为后续的长期储备,促进树干的生长发育[22]。整体来说,对于NSC存储而言,树皮不仅是其传输通道,而且的临时甚至是部分长期储库[23],具有较高的储量。
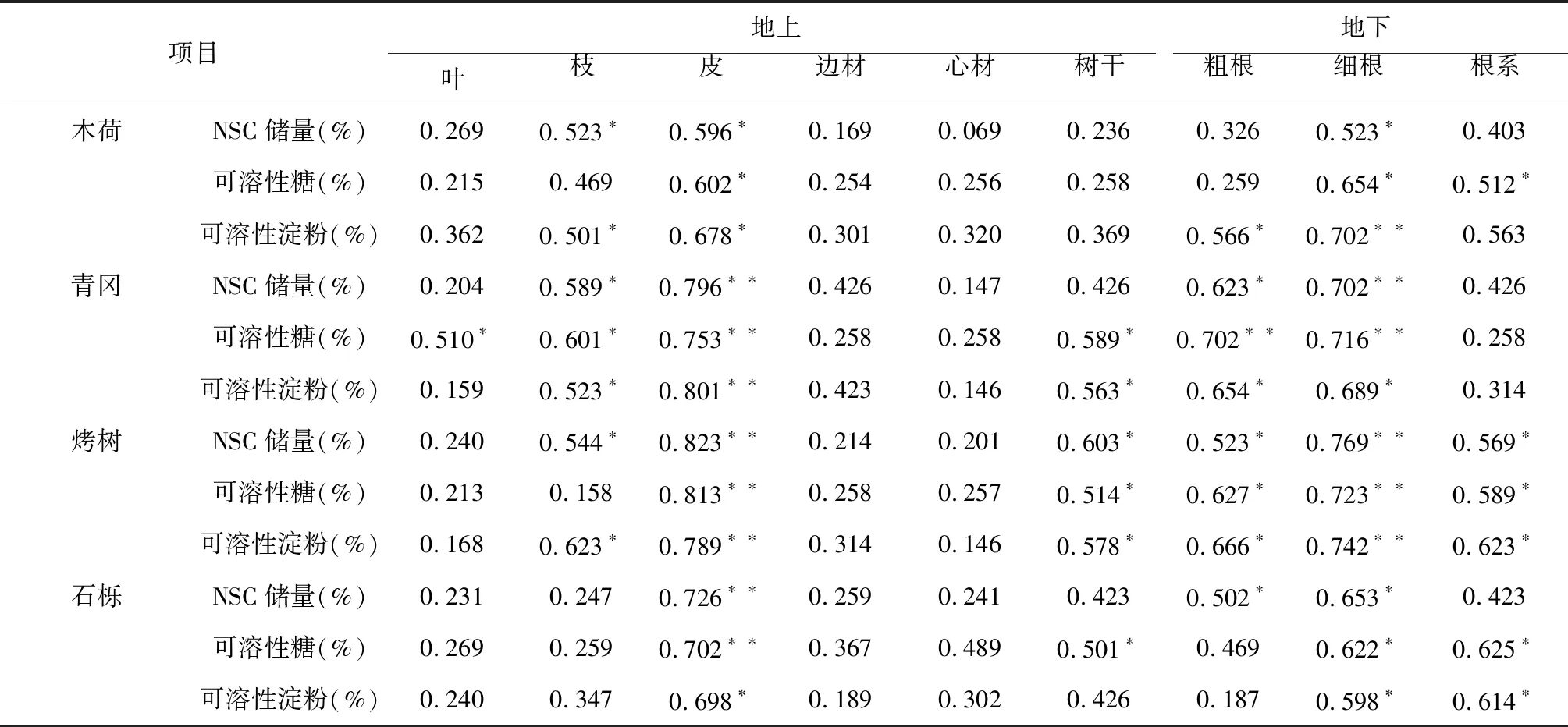
表4 光合速率对NSC的影响
注:*表示0.05差异显著;**表示0.01差异显著。
Note: * indicates significant difference of 0.05; ** mean significant difference of 0.01.
通过实验对比分析得知,树种不同,但是在树干高度不断增加的情况下,其整体NSC存储能力在不断增强,且二者的指数关系在0.05检验水平下达到显著;不仅可溶性糖与之的线性关系达到显著水平,而且淀粉亦是如此,这与以往学者的研究也基本一致[24-25]。冠基及冠中的树干部分具有更强的光合作用,这也是部分针叶林的主要生长量部分,这种径向生长过程利于NSC的存储[22-23],从而使得冠下部分具有较高含量的NSC存储。对于根颈区域而言,其边材及心材的NSC储量在高度上升的情况下出现明显下降。
4 结 论
天童阔叶林不同树种叶片光合速率在一天之内呈“M”型变化趋势,13:00气孔关闭,光合速率出现降低;光合速率在一年之内呈先增加后降低趋势,相同季节光合速率大小基本表现为青冈>烤树>石栎>木荷。叶、枝、皮、边材、心材、粗根和细根NSC储量、可溶性糖和可溶性淀粉呈一致的变化趋势。此外,NSC储量与树干高度呈显著的指数关系(P<0.05),叶、枝、皮、边材、粗根和细根NSC储量和可溶性糖的相对误差的贡献率高于可溶性淀粉相对误差的贡献率,并且不同树种地上相对误差的贡献率高于地下相对误差的贡献率。说明地上部分对NSC储量的贡献高于地下部分。相关性分析表明,光合速率对NSC存储产生显著的影响,二者具有明显的正相关关系;相关系数较高的是烤树和青冈,而石栎和木荷较低,这主要得益于其较高的光合速率水平。
猜你喜欢
草业学报(2022年9期)2022-09-16
林业科学(2022年5期)2022-08-20
热带亚热带植物学报(2022年4期)2022-08-18
东北林业大学学报(2022年6期)2022-07-25
散文诗世界(2022年5期)2022-05-23
绿色科技(2021年5期)2021-04-09
报刊荟萃(上)(2017年7期)2017-07-16
魅力中国(2016年29期)2017-05-27
江苏农业科学(2016年6期)2016-07-25
