
叶片光合生理参数变化特征与小麦受旱状态的关系
2020-05-06 01:14赵福年王润元王鹤龄
干旱地区农业研究 2020年1期
赵福年,王润元,张 凯,齐 月,陈 斐,赵 鸿,王鹤龄
(中国气象局兰州干旱气象研究所,甘肃省干旱气候变化与减灾重点实验室,中国气象局干旱气候变化与减灾重点实验室,甘肃 兰州 730020)
因水分供给不足而引发的干旱是限制全球作物产量增加的重要因素。然而,由于缺乏定量描述作物受旱阶段性特征的指标,目前依然无法准确刻画作物受旱进程[1],这导致我们无法精确模拟干旱胁迫下作物生长发育的动态过程[2],进而影响更加合理的灌溉制度的制定。尽管研究者给出了大量反映作物遭受干旱胁迫的指标(诸如土壤水分、土壤水势、叶片相对含水量、叶水势、冠层温度以及作物水分胁迫指数等[3]),但是这些指标在确定作物受旱程度时,往往以最终产量为标准,即用受旱影响作物与供水充足作物最终产量之间的偏差作为作物受旱严重程度分类的依据[4]。这种划分方法使我们无法有效地掌握干旱对作物生长发育影响的动态过程,阻碍了我们对农业干旱致害机理的认识[5]。因此,有必要在作物生长发育时期,对其受干旱胁迫影响的生理生态参数变化特征进行分析,以量化作物受旱程度。
作物本身具有较强的适应能力及应对干旱的调节能力,作物遭受轻度的干旱胁迫并不会对作物生长发育造成很大的影响[6]。在干旱胁迫发生初期,作物可通过产生大量脱落酸(有促进气孔关闭的功能),降低细胞分裂素(有加剧作物水分亏缺的功能)的含量,调节作物体内细胞分裂素和脱落酸水平及其比例关系能够在一定程度上维持植物体的水分平衡[7]。只有干旱胁迫超过一定的临界值,作物才会遭受明显的干旱影响,即作物生理参数对干旱胁迫存在阈值反应[8],而且不同生理参数的水分阈值不完全相同,这是由作物对干旱胁迫过程的适应和调剂机制不同而决定的[9]。以光合生理过程为例,在遭受干旱胁迫时,作物叶片净光合速率(Pn)的下降并不是唯一一种调节机制在控制,而是既有气孔因素又有非气孔因素的作用[10]。在干旱胁迫相对较轻时,Pn受气孔因素的影响呈缓慢的下降趋势[10];当干旱胁迫继续发展,作物叶片的光合能力受到损害,Pn将受到明显抑制[11];而当严重的干旱胁迫造成叶绿体变形,叶绿体片层膜系统受损,使希尔反应减弱,光系统Ⅱ活力下降,电子传递和光合磷酸酸化受到抑制,则会造成Pn迅速下降[12],从而使作物干物质积累明显减少,作物生长发育受到严重阻碍。
光合生理过程是作物产量形成的基础[13],因而关注与光合生理过程相关的生理生态参数变化是研究作物受旱致害过程的关键。若以作物自适应特点和作物光合生理过程受不同程度干旱胁迫影响机制不同为依据,我们可假定,在作物遭受干旱胁迫至致害过程中光合生理参数应存在不同的变化特征,据此可用以确定作物受旱的不同阶段。此外,一般认为气孔导度(gs)和Pn呈线性关系[14],然而在干旱胁迫发展过程中,气孔因素首先起作用,导致作物Pn下降,而随着干旱胁迫程度加重,非气孔导度因素的出现会导致Pn下降幅度增大,据此,我们推测非气孔因素的叠加作用可能会导致gs与Pn的关系发生改变。
基于以上两个研究假设,我们采用小区和桶栽干旱胁迫试验,以小麦为供试作物,研究其受旱的阶段性特征,从而提供一种可定量描述作物受旱过程的方法。
1 材料与方法
1.1 试验观测
试验在中国气象局兰州干旱气象研究所定西干旱气象与生态环境试验站内进行。该站位于甘肃省定西市安定区西川(104.37°E,35.35°N),海拔1 920 m,年平均气温6.3℃,年日照时数2 500 h。试验供试作物为春小麦(定西新24号)。小区试验于2017年进行,设置2个处理,分别为充足灌水和干旱胁迫处理,充足灌水处理整个生育期保证供水充足,干旱胁迫处理从小麦拔节期开始不灌水直至萎蔫。每个处理设4个重复,每个处理小区面积为2 m×1.5 m。桶栽试验分别在2014、2015年以及2017年进行,桶栽试验所用桶直径29 cm,深度45 cm。供试土壤为黄绵土,从大田采集0~30 cm层次土壤,风干过筛,装桶(每桶装土14 kg)。桶装土平均容重1.15 g·cm3,田间持水量26.8%,萎蔫系数5.5%,其中田间持水量和萎蔫系数均为重量含水量。试验设两个处理,各6组重复,共计12桶。春小麦于3月中旬播种,播种后至对比观测开始前保持12个桶子处理方式相同,土壤水分供给一致。在春小麦长至拔节初期开始设置不同处理,其中一个为充足灌水处理,保证每日的土壤含水量在田间持水量的70%以上,另一个为干旱胁迫处理,在试验开始后持续不灌水,直至小麦萎蔫。
试验期间,主要观测项目为:
(1)土壤含水量(本文均指土壤相对含水量)。每天称重测量2个处理12个桶子质量(所用称重天平最小感应为1g)。用所称得的重量减去干土重量,再除以干土重量即为桶子中土壤重量含水量(忽略小麦本身生物量的变化)。
(2)光合生理过程。采用美国产Li-6400便携式光合仪,在春小麦长至拔节期,不同试验处理实施后,于每日上午9∶00—11∶30测定各处理春小麦最上部第一片完全展开叶Pn/Qp(光响应)和Pn/Ci(CO2响应)曲线(2014年和2015年试验主要测定Pn/Qp曲线,2017年试验主要测定Pn/Ci曲线)。Pn/Qp曲线的测量:控制CO2浓度在380 μmol·mol-1,叶室温度控制在25℃,空气饱和差控制在1.5~2.5 kPa。测量叶室采用红蓝光源,光合有效辐射(PAR)设置不同的梯度自动测量,梯度水平分别为0、15、30、60、120、200、300、600、900、1 200、1 500、1 800 μmol·m-2·s-1以及2 100 μmol·m-2·s-1。每次测量用1 500 μmol·m-2·s-1的PAR适应30~40 min,待仪器读数稳定后,进入自动测量程序。Pn/Ci曲线测量时,初始CO2浓度控制在380 μmol·mol-1,叶室温度控制在25℃,空气饱和差控制在1.5~2.5 kPa之间,光强控制在1 500 μmol·m-2·s-1,开始测量之前先在初始条件下适应30~40 min,设置CO2梯度为400、200、100、50、400、600、800、1 000 μmol·mol-1和1 200 μmol·mol-1,依次自动测量。在观测叶片气体交换的同时,Li-6400还可记录观测时的叶室空气温度、相对湿度、叶气温差、叶气饱和气压差以及空气CO2浓度等气象要素指标。
1.2 光合参数计算
Farquhar等[15]在1980提出了一个计算Pn的生化模型,其以胞间CO2浓度(Ci)分别计算在Rubisco(1, 5-二磷酸核酮糖羧化酶/加氧酶)活性和RuP2(核酮糖-1, 5-二磷酸)再生限制条件下的Pn:
Pn=Vc-0.5V0-Rd=min{Pc,Pj}-Rd
(1)
(2)
(3)
式中,Vc为羧化速率,V0为氧化速率,Rd为暗呼吸速率,Pc和Pj分别为受Rubiso活性和RuP2再生限制的光合速率,Vcmax是最大羧化速率,Г*是CO2补偿点,Kc和Ko是CO2和O2的Michaelis-Menten动力参数,O是叶片胞间氧气分压,J是量子传递效率。(3)式中,J的计算依赖于光强和最大量子传递效率,依据下式计算获得:
θJ2-(I2+Jmax)J+I2Jmax=0
(4)
式中,I2为光强,Jmax为最大量子传递效率,θ为曲度。
Vcmax和Jmax反映了作物叶片的光合能力,其值由Pn/Ci曲线观测数据估算。当Ci值小于150 μmol·mol-1时,以(2)式估算Vcmax,当Ci大于150 μmol·mol-1时,用(3)和(4)式估算Jmax,具体计算过程可参照Xu等[16]的研究工作。
1.3 气孔导度模型
Pn与gs是描述植物叶片气体交换过程非常重要的两个指标,而它们之间的关系又可由内禀水分利用效率(IWUE, 即Pn/gs)或半经验气孔导度模型反映[17]。当前有很多半经验气孔导度模型,而其中尤以Ball-Berry[18]模型的使用最普遍。Ball等[18]根据前人的研究结果,发现gs的变化与Pn、叶片表面相对湿度(RHs)以及叶片表面CO2浓度(Cs)有关:
gs=a1AnRHs/Cs+b1
(5)
式中,a1为气孔导度模型斜率,由gs与PnRHs/Cs(BB参数)关系拟合获得。b1为净光合速率为零或小于零时的gs,部分文献认为其为无光照时的gs[17],可由gs与PnRHs/Cs关系拟合截距获得,或在黑暗条件下由光合仪测量获得。Leuning[19]与Medlyn等[20]此后陆续分别提出了与Ball-Berry类似的2个模型,即Leuning和Medlyn模型,它们对gs模拟精度的改进有一定的帮助。其中Leuning(1995)模型的形式为:
gs=a2An/(Cs-Γ)(1+VPDa/b2)+c2
(6)
式中,a2为气孔导度模型斜率,Г是CO2补偿点,VPDa是空气饱和气压差,b2和c2都是参数。Medlyn模型的形式为:
(7)
式中,a3为气孔导度模型斜率,VPDL是叶片与空气之间的饱和气压差,b3为参数。
1.4 反映干旱胁迫过程的指标
本研究采用土壤有效含水量(ASWC)和饱和光强下的气孔导度(gssat)来反映小麦遭受干旱胁迫的状况。其中ASWC计算如下:
(8)
式中,θa、θw以及θf分别表示实际土壤含水量、萎蔫含水量以及田间持水量。研究者发现gssat较其他水分指标更能有效地反映作物遭受干旱胁迫的状况[21],由于本研究小区试验小麦最大根系深度未知,因此以gssat作为另一个反映小麦水分供给状况的指标。已有研究表明小麦光饱和点处于1 200~1 700 μmol·m-2·s-1之间[22],本文选择光合有效辐射在1 500 μmol·m-2·s-1时的gs作为小麦的gssat。
1.5 数据分析
Pn/Qp曲线数据主要用以拟合求取3个不同气孔导度模型的参数,在Origin 9.0软件中使用非线性曲线拟合模块(Nonlinear curve fit),自定义编制与(5)、(6)以及(7)式相同的公式拟合计算获得所需参数。Pn/Ci曲线观测数据主要用以估算Vcmax和Jmax,采用R语言以Duursma等[23]给出的“plantecophys”包计算获得。同时,为证明干旱胁迫阶段对气孔导度模型参数的影响,本研究选择不同年份(2014、2017年)和不同观测来源(Pn/Qp和Pn/Ci曲线)数据,先以2014年Pn/Qp曲线数据拟合计算了不同干旱胁迫条件下不同光强的gs,并与实测值进行了对比分析;之后又将小区和桶栽2017年的数据整合,提取了饱和光强下的光合生理参数数据,分析比较了不同干旱胁迫阶段气孔导度模型参数的差异。
2 结果与分析
2.1 干旱胁迫下小麦叶片光合生理参数的变化
2.1.1 光合生理参数对干旱胁迫的响应 分析gssat与ASWC的关系(图1a),发现gssat随ASWC的变化存在3个阶段,在第1个阶段gssat变动不明显,呈轻微的下降趋势;而当ASWC减少至0.5左右时,gssat呈减小趋势,但统计检验不显著(P>0.05);而在第3阶段,当ASWC小于0.3左右时,gssat快速线性下降,且统计检验极显著(P<0.01)。同时,比较这3个阶段的gssat,发现差异极显著(图1b)。
与gssat类似,Ci/Ca值随着ASWC的变化也存在3个明显的阶段(图1c),只是变化趋势不完全相同。第1阶段,Ci/Ca相对稳定,之后快速减小,在ASWC降至0.06开始,Ci/Ca呈增大的趋势,最小值出现在ASWC为0.1时,此时Ci/Ca为0.32。同时,比较3个阶段的Ci/Ca(图1d),第2阶段与其他2个阶段差异极显著,而第1和第3阶段无显著差异。
Pn随ASWC的变化存在2个明显的阶段(图1e),在第1阶段,Pn基本保持稳定,而在ASWC降至0.35左右时,Pn随ASWC的减少呈线性减小趋势,且统计检验极显著。而且第1阶段与第2阶段Pn值的差异也极显著(图1f)。
2.1.2 光合生理参数间的关系Ci/Ca和gs是决定作物叶片Pn的重要指标,观察gssat以及Ci/Ca与Pn间的关系(图2),这2个参数与Pn并不呈简单的线性关系,而是随水分条件变化存在阶段性关系特征。
当gssat大于0.15 mol·m-2·s-1时,Pn随gssat减小缓慢下降(图2a),速率为1.64 μmol·m-2·s-1Pn/0.1 mol·m-2·s-1gs(P<0.001),而当gssat小于0.15 mol·m-2·s-1时,Pn随gssat的降低快速减小,速率增至11 μmol·m-2·s-1Pn/0.1 mol·m-2·s-1gs(P<0.001),说明在以gssat=0.15 mol·m-2·s-1为界的2个不同阶段,Pn对gssat变化的响应不同,即控制Pn变化的因素存在差异。
Ci/Ca与Pn的关系也存在2个明显的阶段(图2b),即Pn与Ci/Ca呈线性正相关关系和Pn随Ci/Ca增大而减小的阶段。在第1阶段,Pn随Ci/Ca的减小呈显著的减小趋势,但是仔细观察该阶段依然可以分为2个子阶段,即在Ci/Ca从0.8降至0.55的阶段(gssat>0.06 mol·m-2·s-1),Pn随Ci/Ca的变化降幅较小,而后下降速率加快。而当Ci/Ca降至0.32后(gssat<0.06 mol·m-2·s-1),Ci/Ca开始增大,但Pn却随Ci/Ca的增大依然在降低。
2.2 小麦叶片光合能力参数对干旱胁迫的响应
观察小麦叶片Vcmax与Jmax在不同水分条件下的变化(图3a,c),无论是在小区还是桶栽环境,小麦叶片的这2项参数对水分变化均存在明显的阈值反应。当gssat大于0.15 mol·m-2·s-1时,随gssat的降低,Vcmax和Jmax均保持相对稳定。而当gssat小于0.15mol·m-2·s-1时,随gssat的降低,Vcmax和Jmax均呈快速降低趋势(P<0.01)。而若以gssat=0.15 mol·m-2·s-1为界,再以生长环境为类别(小区和桶中),可将Vcmax和Jmax分为4组,经统计分析发现在gssat小于0.15 mol·m-2·s-1时,2个生长环境下的小麦叶片Vcmax和Jmax均小于gssat大于0.15 mol·m-2·s-1时的值,且统计检验极显著(图3b,d)。而在相同的gssat范围内,不同生长环境下除Vcmax大田和桶栽有极显著区别外,其他统计检验均不显著。
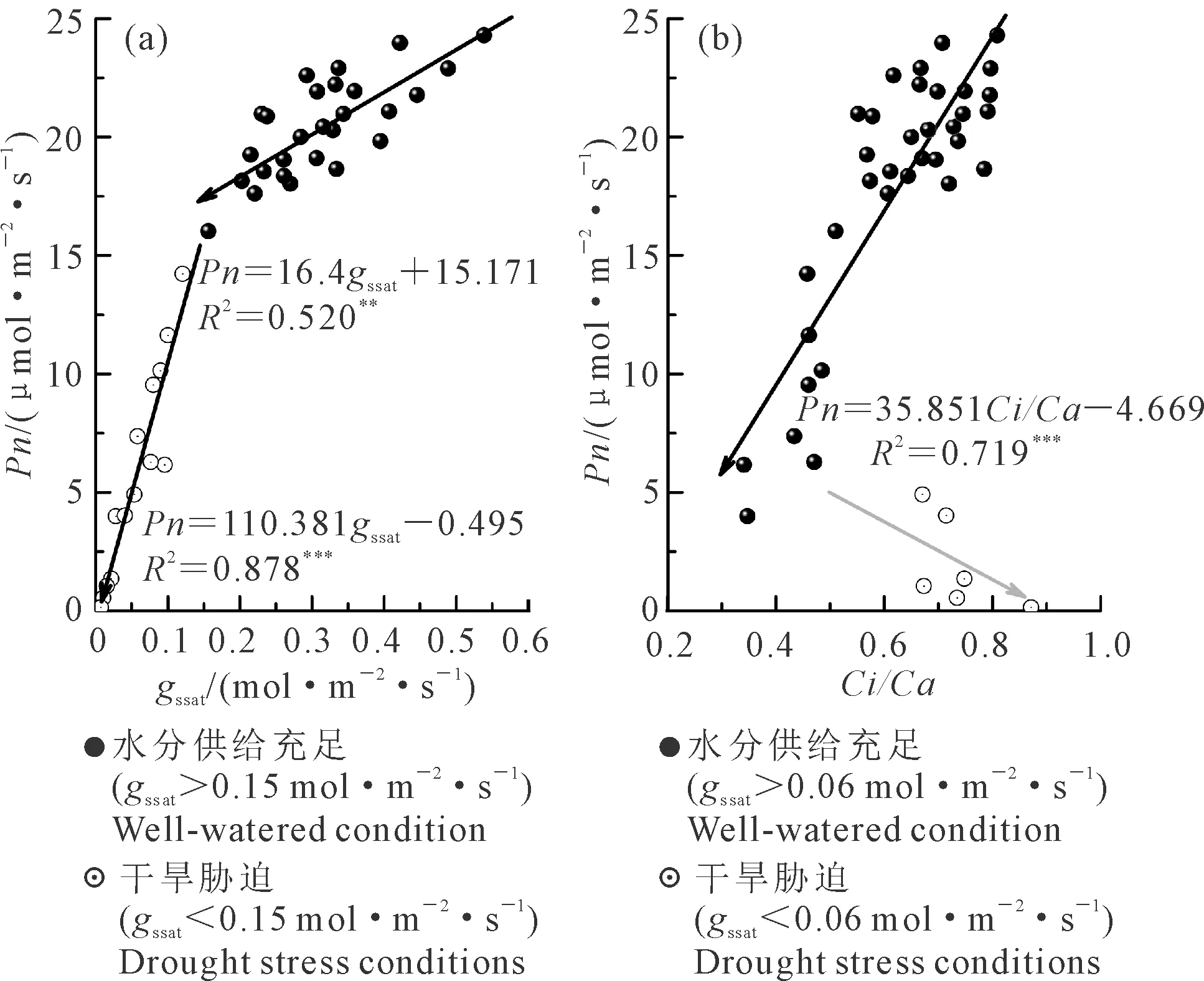
图2 不同光合生理参数间的关系Fig.2 Relationship between different photosynthetic parameters
以gssat0.15 mol·m-2·s-1为界混合分析小区和桶栽小麦Vcmax和Jmax的关系(图4a),发现当gssat大于0.15 mol·m-2·s-1时,Vcmax和Jmax关系并不明确,在相同的Vcmax值下,Jmax变幅较大。而当gssat小于0.15 mol·m-2·s-1时,Vcmax随Jmax的减小呈极显著的降低趋势,同时比较以气孔导度0.15 mol·m-2·s-1为界2个阶段的Vcmax/Jmax(图4b),发现统计检验极显著。说明不同的水分条件影响Vcmax和Jmax的相互关系。
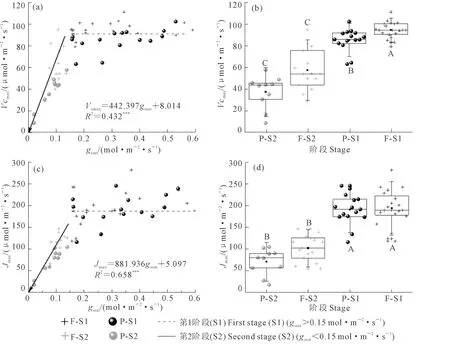
注:F-S1表示小区试验光合生理参数变化的第1阶段(gssat>0.15 mol·m-2·s-1),F-S2表示小区试验光合生理参数变化的第2阶段(gssat<0.15 mol·m-2·s-1);P-S1表示桶栽试验光合生理参数变化的第1阶段,P-S2表示桶栽试验光合生理参数变化的第2阶段。图6相同。Note:F-S1, F-S2, P-S1, and P-S2 indicate the first (gssat> 0.15 mol·m-2·s-1) and second (gssat< 0.15 mol·m-2·s-1) stages of photosynthetic parameters variation for spring wheat growing in plots and pots, respectively. The same for Fig. 6.图3 光合能力参数对干旱胁迫的响应Fig.3 Response of photosynthetic capacity to water stress
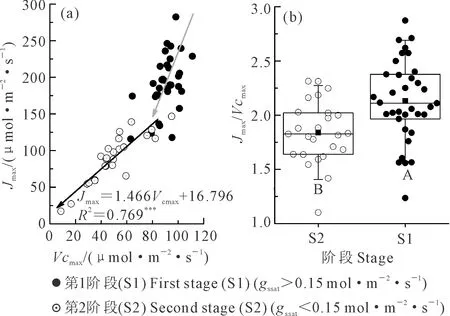
图4 不同干旱胁迫阶段Vcmax与Jmax的关系Fig.4 Relationship between Vcmax and Jmax under ifferent water stress stages
2.3 干旱胁迫下小麦叶片气孔导度与净光合速率之间的关系
若不区分干旱胁迫阶段,使用拟合获得的气孔导度模型斜率计算2014年所观测不同光强下的gs(图5a、b、c),发现3个典型的气孔导度模型实测值与模拟值均由1∶1线将数据明显分为两部分,其中一部分数据观测值明显偏大,而另一部分模拟值明显偏大。对数据进行分类发现模拟值偏大的数据来源于gssat小于0.15 mol·m-2·s-1的部分,而实测值偏大的数据来自gssat大于0.15 mol·m-2·s-1的部分。而如果以gssat在0.15 mol·m-2·s-1处为界,将数据以2个不同的供水阶段进行划分,再次分析模拟值与观测值的关系,发现模拟值与实测值的决定系数较不区分干旱胁迫阶段时有较大的提高(图5d、e、f),说明模型参数的选择对气孔导度模拟存在影响。
以Ball-Berry(1987)气孔导度模型为例(其他2个模型结果类似,图省略),整合大田和桶栽的数据,选择饱和光强下的数据拟合气孔导度模型,发现无论是生长在小区还是桶栽环境中的小麦,以gssat在0.15 mol·m-2·s-1为分界,不同水分供给阶段gssat与BB指数的关系不完全一致(图6)。在水分供给相对充足时(gssat大于0.15 mol·m-2·s-1),无论是小区还是桶栽,拟合所得的小麦气孔导度模型斜率a1均较受水分胁迫程度严重时(gssat小于0.15 mol·m-2·s-1)的值大,其中在水分相对充足时小区和桶栽小麦a1分别为10.8和10.6,而水分相对不足时a1分别为7.5和6.9,水分供给条件不同,a1统计检验差异极显著,而在相同水分条件下,不同生长环境对a1无显著影响。
3 讨 论
3.1 春小麦受旱过程的阶段性划分
gssat和Ci/Ca随水分条件变化存在明显的3个阶段,而Pn的变化则存在2个阶段,Pn与gssat以及Ci/Ca的关系也因不同水分条件存在2~3个阶段。综合来看,小麦叶片光合生理过程对干旱胁迫的响应存在4个明显的阶段,这种阶段性变化是由于作物自身在随干旱胁迫程度不断加重时相应的调节机制不同决定的。
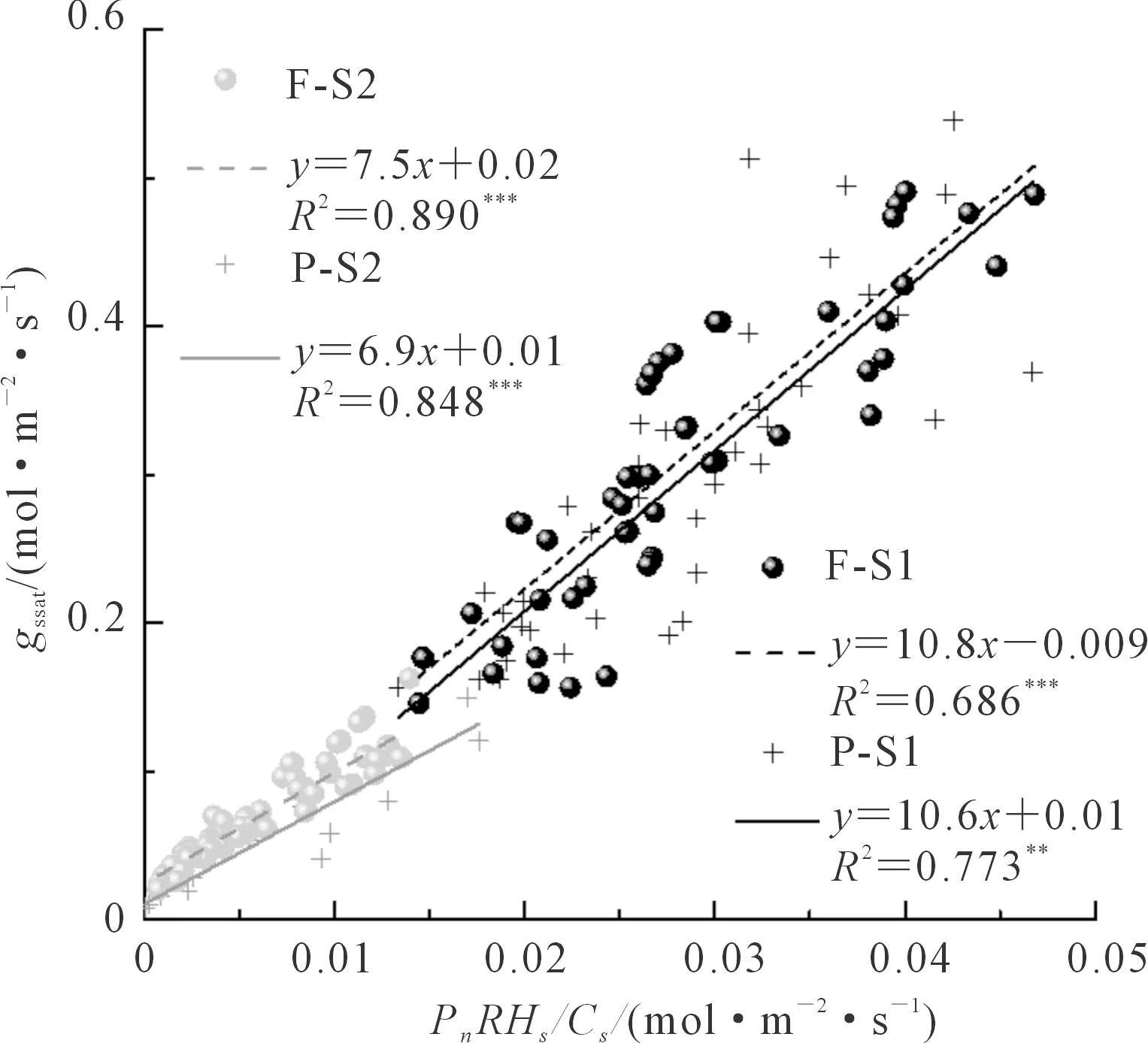
图6 不同水分条件下的气孔导度模型Fig.6 Stomatal conductance models under different water conditions
在干旱胁迫之初,随着水分供给的减少,作物能够通过自身的调节,适应水分的减少。当ASWC从1变化至0.5时(gssat>0.35 mol·m-2·s-1的阶段,第1阶段)(图1a),gssat略有降低,而Ci/Ca值却相对比较稳定,因此Pn无明显的减少趋势。而相较CO2,水分对gs的变化更敏感(CO2通过气孔的阻力是水分的1.6倍),过大的蒸腾速率在该阶段对作物的光合产出而言是无效的,该阶段的光合生理参数变化特征间接说明了为什么有的研究发现轻到中度干旱胁迫下,植物叶片尺度水分利用效率(Pn与蒸腾速率比)或IWUE存在随供水减少而增大的现象[10]。
当ASWC从0.5降至0.3时,gssat随供水的减少呈波动减少趋势(从0.35降至0.15 mol·m-2·s-1时,第2阶段),但Ci/Ca在该阶段呈极显著的降低趋势(直至ASWC降至0.1),而Pn也在该阶段随水分减少由稳定少动转变为逐渐减小。同时观察Vcmax和Jmax在该阶段随水分供给的变化保持相对稳定(图3),Pn与gssat的关系以及Ci/Ca与Pn的关系也无明显变化(图2)。据此,我们推测,在该阶段气孔因素在起主导作用而影响小麦光合生理过程,gs的变化导致Ci/Ca值出现波动,从而使得小麦供给光合作用的场所,即叶绿体可用的CO2量减少。由于光合原材料不足,达到一定程度时,小麦叶片Pn出现降低的趋势。
而在ASWC<0.3后,gssat进入一个新的变化阶段(gssat<0.15 mol·m-2·s-1时,第3阶段),即随ASWC减小呈极显著的降低趋势。此外,从Vcmax和Jmax与gssat的关系来看(图3),在该阶段光合能力从之前的相对稳定到该阶段开始随干旱胁迫程度的增加而逐渐减小。而且Vcmax和Jmax之间的关系也以gssat在0.15 mol·m-2·s-1处为界有明显的不同,gssat>0.15 mol·m-2·s-1时,Vcmax/Jmax值与许多作物水分供给充足条件下的研究结果一致[24],而当gssat<0.15 mol·m-2·s-1时,Vcmax/Jmax值明显小于水分较为充足下的值,这与部分林木的观测结果相近[25]。同时,gssat与Pn的关系也在气孔导度<0.15 mol·m-2·s-1后发生变化,Pn随gssat降低的幅度增加。这些现象充分说明,除气孔因素外,与小麦光合生理过程相关的其他代谢活动(即非气孔因素)由于干旱胁迫程度的增加开始限制Pn。
当ASWC降至0.1之后(气孔导度<0.06 mol·m-2·s-1时,第4阶段),随作物供水的进一步减少,Ci/Ca呈增加趋势,这是光合生理过程遭受干旱胁迫影响不同于上述3个阶段的新时期。从Ci/Ca与Pn的关系(图2b)也可以发现,该阶段Ci/Ca值的变化已不能反映Pn的变化。然而,过去较早的一些研究结果往往以Ci/Ca随干旱胁迫的加剧而出现增大的时期定义为作物光合生理过程遭受非气孔因素影响开始的阶段[26]。但本研究结果显示在gssat小于0.15 mol·m-2·s-1时,影响作物生理过程的非气孔因素已经出现,这与近年来的一些研究结果相近[27],即非气孔因素可能在中等干旱条件下就已开始限制光合生理过程。本研究的第4阶段,Ci/Ca的快速增大可能是由于严重的干旱胁迫导致小麦光合器官受损,使得光合生理过程出现了不可逆的破坏,这也是部分研究发现干旱胁迫到一定程度,即使恢复供水,而植物光合生理过程也无法恢复的原因[28]。
本研究对小麦受旱的阶段性划分与国外一些研究用gssat界定C3植物葡萄的受旱程度结果类似,即在葡萄生长遭受轻旱时,气孔导度介于0.5或0.7 mol·m-2·s-1到0.15 mol·m-2·s-1,中旱0.05~0.15 mol·m-2·s-1,重旱<0.05 mol·m-2·s-1[29]。然而考虑到不同试验条件,不同作物类型、土壤质地以及施肥条件等因素的影响,这种结论的相似性是偶然还是必然,还需进一步通过大量的试验结果整合分析后确定。
3.2 不同干旱胁迫程度下气孔导度模型的差异及其意义
气孔导度模型斜率在干旱胁迫条件下是否会发生变化依然存在许多争议,很多试验研究结果相互矛盾[30-31]。本研究发现以gssat=0.15 mol·m-2·s-1为界,gs与Pn的关系有明显的区别,而若不区分不同的干旱胁迫程度对气孔导度模型参数的影响,3个典型的气孔导度模型均会错估gs。出现这种现象的原因是以gssat=0.15 mol·m-2·s-1为界,当gssat大于0.15 mol·m-2·s-1时,只有气孔因素在限制光合生理过程,气孔的调节作用导致CO2进入光合场所叶绿体的数量出现波动,引起Pn变动;而在gssat小于0.15 mol·m-2·s-1时,干旱胁迫加剧,非气孔因素开始影响光合能力,而此时气孔因素并非引起Pn变动的唯一因素,因此Pn与gs的关系必然发生变化,即在相同的gs变幅下,Pn变动更剧烈,这在图3agssat与Pn的相互关系中也有反映,即随干旱胁迫的加重,小麦IWUE在增大。因此,我们在使用气孔导度模型时,需要根据干旱胁迫程度,选择合适的模型参数。否则可能出现在水分供给充足时低估gs,干旱胁迫发生时高估gs的情形(图5)。而其他一些研究[32]也表明,不准确的气孔导度模型斜率不仅影响gs的准确计算,还导致对Pn、总初级生产力、蒸腾速率以及水分利用效率的错误估计。
气孔导度模型在不同干旱胁迫阶段的斜率差异,一方面表明气孔导度模型需要根据作物所受胁迫程度调整参数从而以模拟获得准确的gs,另一方面也再次间接证明了干旱胁迫的阶段性特征,即在其他环境要素一定时,干旱胁迫条件下,以gssat=0.15 mol·m-2·s-1为界,大于该值,光合生理过程处于气孔为主要限制因子影响Pn变化的阶段,而小于该值后,转变为非气孔因素开始限制作物叶片光合生理过程的阶段。
3.3 本研究存在的不足与展望
近年来学者认为干旱胁迫条件下叶肉导度的下降是导致光合生理过程下调的重要因子[33],然而在本研究中,并没有单独考虑叶肉导度在不同干旱胁迫阶段对光合生理过程的影响,而是将其归为非气孔因素。在干旱胁迫条件下,叶肉导度的快速降低可能会影响Ci的准确测定,从而影响对Vcmax和Jmax的计算。但是由于叶肉导度的定量估计依然存在许多不确定性,因此分析叶肉导度在非气孔因素中所占的比例是干旱胁迫对光合生理过程影响研究的重要方面,同时这也是农业干旱致害过程试验研究与数值模拟亟需解决的问题。
过去的研究在确定作物受旱等级时,往往以产量作为划分标准。而本研究所涉及的光合生理过程只是决定作物产量高低的众多因素之一。而且作物不同生育期遭受干旱胁迫,对产量的影响不完全相同。例如,许多研究得出与光合生理过程相比,叶片的生长对干旱胁迫更敏感[34],在生育期前期,作物叶面积受旱而停止生长可能对产量影响更大。在自然条件下,一次降水或恢复供水后,光合等生理参数可能会迅速恢复至正常水平,但是叶面积的生长却因水分亏缺而停滞,即使恢复供水也无法达到水分供给充足条件下的水平。作物光照截获面积的减少,必然导致作物生物量积累减少。因此在研究农业干旱时,特别需要关注干旱胁迫对一年生作物生长盛期叶片生长的影响。本文只针对选定的生育期进行研究,没有观测叶面积变化对干旱胁迫的响应,在实际干旱监测中可能依然无法定量估计作物不同生育期所遭受干旱对其最终产量的影响。干旱胁迫对叶面积和光合生理过程的影响及二者对作物最终产量减损的作用还需进行更深入地分析和探讨。
4 结 论
本研究依据试验分析探讨了光合生理过程对干旱胁迫的阶段性响应特征,从而定量确定了干旱胁迫发展的不同阶段,并讨论了不同干旱胁迫阶段Pn与gs的关系及其对气孔导度模型使用的影响。小麦光合过程受干旱胁迫的影响,根据胁迫影响程度可划分为4个阶段,其临界值以ASWC表示分别为0.48、0.3和0.1,而相对应的gssat为0.35、0.15以及0.06 mol·m-2·s-1。非气孔因素开始影响光合生理过程的临界点在ASWC为0.3时,非气孔因素的出现导致Pn与gs的关系发生变化,从而在使用气孔导度模型时,需要考虑不同的干旱胁迫程度对气孔导度模型参数的影响,否则会出现水分供给充足时低估gs,干旱胁迫时高估gs的情形。本研究结果可为农业干旱等级定量划分、作物模型改进以及农田灌溉制度的确定提供帮助。
猜你喜欢
航空材料学报(2022年4期)2022-08-04
贵州农业科学(2022年4期)2022-05-06
农业工程学报(2022年3期)2022-04-16
干旱地区农业研究(2022年1期)2022-01-28
农业工程学报(2021年3期)2021-04-15
节水灌溉(2020年9期)2020-09-16
节水灌溉(2020年7期)2020-07-15
音乐研究(2019年5期)2019-11-22
江苏农业学报(2019年1期)2019-09-10
科技创新与品牌(2019年12期)2019-02-06
