
培养条件及添加剂对重组荧光藻蓝蛋白β亚基胞外分泌的影响
2020-07-07 03:15赵保国蒋苏丹汪长天伍贤军李萍萍
食品与发酵工业 2020年12期
赵保国,蒋苏丹,汪长天,伍贤军,2*,李萍萍,2*
1(南京林业大学 生物与环境学院,江苏 南京,210037)2(南京林业大学,江苏省南方现代林业协同创新中心,江苏 南京,210037)
藻蓝蛋白是存在于藻类中的捕光色素蛋白。从螺旋藻中获取的C-藻蓝蛋白(C-phycocyanin,C-PC)在医疗保健、食品添加剂、化妆品着色剂和荧光染料等方面有广泛的应用前景[1],特别是其抗氧化、抗癌、抗炎和增强免疫力的功能引起广泛的关注[2-4]。C-藻蓝蛋白由α和β亚基组成,开链的四吡咯藻蓝色素(phycocyanobilin,PCB)共价结合在蛋白质保守性半胱氨酸结合位点上。由于从分子水平上对PCB生物合成过程的解析以及对藻蓝蛋白裂合酶的鉴定,C-藻蓝蛋白α和β亚基均可在大肠杆菌体内重组产生。其中,β亚基脱辅基蛋白可在藻胆蛋白裂合酶(phycobiliprotein lyase,CpcS)的催化下结合PCB[5]。而PCB来源于对大肠杆菌血红素底物的氧化还原反应,即在血红素氧化酶(heme oxidase-1,HO1)的作用下将血红素转化成胆绿素(biliverdin,BV),在胆绿素还原酶(ferredoxin oxidoreductase,PcyA)的作用下最终合成PCB。研究表明,大肠杆菌重组藻蓝蛋白β亚基(β-subunit of C-phycocyanin,CpcB)展现出显著的抗氧化和自由基清除活性[6-8]。为获取在大肠杆菌体内重组的外源蛋白,通常需要破碎细胞,这经常会导致热原水平和样品杂质的增加以及蛋白质活性的降低。特别地,当目的蛋白在胞内过量表达时,会形成包涵体。在重组藻蓝蛋白的过程中,通常用降低表达温度的方法减少包涵体的形成,使得色素蛋白能有效合成和正确折叠[9]。但是对于大量生产和工业发酵,这会降低产量和增加生产成本。为了克服这些缺陷,采用胞外分泌重组藻蓝蛋白是有效的方法。为促进重组蛋白的胞外分泌,各种策略被广泛研究,如信号肽优化[10]、细胞质渗漏[11]、分子伴侣共表达[12]等。除此之外,还可以对培养条件进行优化,如调整温度,改变诱导剂以及使用影响细胞膜通透性的化学物,如甘氨酸、Triton X-100、十二烷基硫酸钠(sodium dodecyl sulfate,SDS)和Tween-80等[13-15]。通过这些方法可以显著强化重组蛋白的分泌。目前,藻蓝蛋白胞外分泌的特征及影响因素并不清楚,特别是藻蓝蛋白的色素化增加了胞外分泌的复杂性。本研究通过改变温度,利用乳糖作为诱导剂,利用培养基添加剂等方式促进螺旋藻重组荧光藻蓝蛋白CpcB的胞外分泌,并以藻蓝蛋白荧光强度为表征[16],探讨影响藻蓝蛋白胞外分泌的因素。
1 材料与方法
1.1 材料与试剂
含有表达质粒pETDuet-cpcB-cpcS∶∶ho1 ∶∶pcyA的重组E.coliBL21(DE3)菌株pBSA,由本实验室构建并保存[6],其中表达载体pETDuet-1,购于Novagen公司,藻蓝蛋白β亚基脱辅基蛋白编码基因cpcB来自盐泽螺旋藻SpirulinasubsalsaFACHB351,藻蓝蛋白裂合酶编码基因cpcS、血红素氧化酶编码基因ho1、胆绿素还原酶编码基因pcyA,均来自于Nostocsp. PCC 7120。
蛋白胨、酵母粉,OXOID公司;甘油、氨苄西林,盛兴生物科技有限公司;异丙基硫代半乳糖苷(isopropyl-β-D-thiogalactoside,IPTG),北京博奥拓达科技公司产品;乳糖,上海凛恩科技发展有限公司;甘氨酸和Triton X-100,广州赛国生物科技有限责任公司;SDS,上海凌峰化学试剂有限公司;Tween-80,国药集团化学试剂有限公司。
1.2 仪器与设备
Lambda 365紫外/可见分光光度计和LS 55荧光光谱仪, 美国Perkin Elmer公司;ZWY-240恒温摇床, 上海智诚分析仪器制造有限公司;MIKRO 200R高速离心机, 德国Hettich公司;BG-verMIDI蛋白质电泳仪,北京百晶生物技术有限公司;Sigma 3-18KS冷冻离心机,德国Sigma离心机有限公司;GenoSens1850凝胶成像仪, 上海勤翔科学仪器有限公司;BXM-30R立式压力蒸汽灭菌器,上海博迅医疗生物仪器股份有限公司。
1.3 实验方法
1.3.1 菌株培养及重组蛋白表达
少量接种实验室保存的重组菌株pBSA到5 mL含100 mg/L氨苄抗生素的LB培养基中,置于恒温摇床中,37 ℃、150 r/min过夜培养。转接0.5 mL过夜培养的新鲜菌液到50 mL TB培养基中,在37 ℃、220 r/min的条件下继续培养,直到OD600值为0.6,4 ℃保存。
为研究表达温度对重组荧光藻蓝蛋白胞外分泌的影响,按照上述方法培养3组50 mL菌液,添加或不添加1 mmol/L的IPTG的情况下,分别置于温度为16、25、37 ℃的恒温摇床中200 r/min培养,每隔12 h取3 mL菌液,12 000 r/min离心10 min弃沉淀,取培养基上清液,检测CpcB的荧光强度。实验独立重复3次,最终培养基中CpcB的荧光强度取3次平均值。
为研究诱导剂乳糖对CpcB胞外分泌的影响,分别转接0.5 mL过夜培养的新鲜菌液到6组50 mL TB培养基中,37 ℃、220 r/min培养,直到OD600值为0.6,冰浴30 min。各组分别加入终质量浓度为5、7.5、10、15、20 g/L的乳糖,并以未加乳糖的作为对照。置于摇床,200 r/min,25 ℃诱导表达24 h,取3 mL菌液12 000 r/min离心20 min弃沉淀,取培养基上清液检测CpcB的荧光强度和OD600值。实验独立重复3次,最终数据取3次平均值。
为探索培养基添加剂(甘氨酸,Triton X-100,SDS,Tween-80)对CpcB胞外分泌的影响,按照上述方法培养若干50 mL的重组大肠杆菌菌液冰浴。菌液分成4大组,每1大组分成6小组,加1种添加剂。其中甘氨酸的终质量浓度为15、20、25、30、35 g/L,Triton X-100的终体积分数为0.1%、0.2%、0.5%、0.7%和1%,SDS的终质量浓度为0.5、1、2、5、10 g/L,Tween-80的终体积分数为1%、1.5%、2%、2.5%和5%,并以未加添加剂的菌液为对照,置于摇床中200 r/min培养24 h,取3 mL菌液12 000 r/min离心20 min弃沉淀,取培养基上清检测CpcB荧光强度。实验独立重复3次,最终数据取3次平均值。
1.3.2 重组蛋白纯化、聚丙烯酰胺凝胶电泳(polyacrylamide gel electrophoresis,SDS-PAGE)及锌电泳
为确认重组蛋白CpcB的光谱特征和分子质量,用100 mL的TB培养基在200 r/min,25 ℃条件下表达重组菌株24 h,全部菌液12 000 r/min离心20 min弃沉淀,取上清对培养基中的CpcB进行纯化,并取少量的培养基制备电泳样品,准备SDS-PAGE电泳。由于表达载体pETDuet含有His标签,故可以利用镍离子亲和层析柱进行蛋白质的纯化,并透析去除咪唑[8]。纯化后的CpcB溶液可用于光谱分析;同时取少量CpcB溶液制备电泳样品。制备好的蛋白质样品用12%的分离胶进行SDS-PAGE电泳,电泳完成后将蛋白质胶用 1.5 mol/L 醋酸锌溶液浸泡15 min染色,置于凝胶成像仪中用紫外光照射成像,凝胶再用考马斯亮蓝染色后成像。
1.3.3 荧光检测和光谱分析
利用荧光光谱仪(Perkin Elmer LS 55,美国)检测胞外培养基CpcB的荧光强度,所用激发波长为600 nm,收集的荧光发射波长为640 nm。由于重组荧光藻蓝蛋白的荧光强度和表达量在一定范围内存在线性关系[16]。故可以通过荧光强度反映藻蓝蛋白的产量,即可以通过培养基荧光CpcB的荧光强度大致反映胞外分泌的CpcB浓度。利用荧光光谱仪检测胞外培养基CpcB的荧光发射光谱,激发波长为600 nm。光谱扫描范围300~800 nm,光谱扫描速度1 200 nm/min,狭缝宽度10.0 nm。吸收光谱用紫外可见分光光度计检测,光谱扫描范围200~800 nm,扫描速度960 nm/min,狭缝宽度1.0 nm。
2 结果与分析
2.1 温度对CpcB胞外分泌的影响
在大肠杆菌胞内合成藻蓝蛋白的过程中,最佳温度常设定在18 ℃左右[9]。本研究中,首先参照胞内合成的条件,在16 ℃的条件下加入1 mmol/L的IPTG表达CpcB直到60 h,如图1-a所示,胞外培养基中没有检测到任何CpcB荧光;在没有IPTG的条件下,如图1-b所示,也没有检测到荧光。低温下胞内合成的藻蓝蛋白不向胞外分泌,这可能与低温抑制细胞的生长率和转运蛋白的表达有关[17]。而在37 ℃,无论是添加IPTG还是没有添加IPTG,都不能检测到培养基中CpcB的荧光。较高温度下过表达的蛋白质容易形成包涵体并在内膜附近积累,阻断蛋白质的易位通道,减少分泌[18]。另外,藻蓝蛋白的色素化也可能受阻。在25 ℃没有IPTG存在的情况下,如图1-b所示,表达24 h后培养基展现明显的CpcB荧光,并且随着表达时间的增加,荧光强度逐步升高。这些结果表明温度是影响胞外分泌的关键因素,而25 ℃是促进藻蓝蛋白胞外分泌的较好温度,这与先前研究的其他蛋白胞外分泌最佳温度一致[13],表达温度应该保持细胞生长和蛋白合成的平衡以促使胞外分泌。

a-不同温度有IPTG诱导对培养基CpcB荧光强度的影响; b-不同温度无IPTG诱导对培养基CpcB荧光强度的影响
图1 温度对胞外培养基CpcB荧光强度的影响
Fig.1 Effect of temperature on the fluorescence intensity of the extracellular CpcB in medium
本研究中,诱导剂IPTG的存在使得胞外CpcB产生受阻,这可能与IPTG抑制细菌生长及自身蛋白的表达,影响重组CpcB的分泌通道有关。不过先前的研究利用IPTG诱导外源蛋白在信号肽的作用下是可以分泌的[14],有可能归因于先前研究中信号肽的强化作用,但需要进一步的证实。检测胞外培养基荧光发射光谱,如图2所示,在644 nm处展现最大荧光发射峰,这和先前CpcB的光谱特征一致[6]。
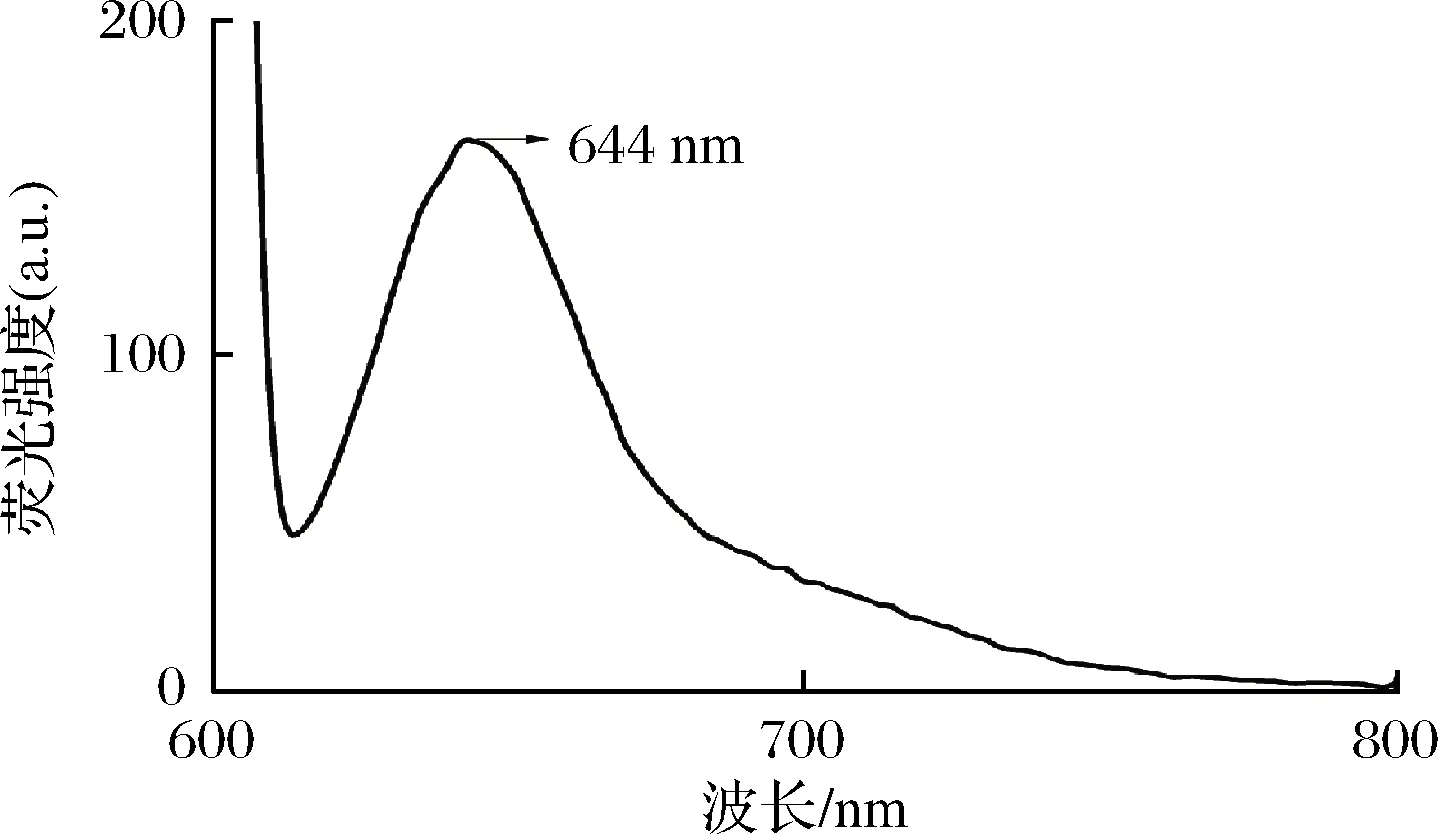
图2 大肠杆菌25℃表达24 h后胞外培养基CpcB的 荧光发射光谱
Fig.2 Fluorescence emission spectrum of the extracellular CpcB expressed byE.colifor 24 h at 25℃
为进一步鉴定培养基中的CpcB,25 ℃表达100 mL重组细胞,24 h后对培养基进行纯化,检测纯化后蛋白溶液的吸收光谱和荧光发射光谱。如图3-a﹑图3-b所示,纯化后蛋白溶液展现出621 nm的最大吸收值和644 nm的最大荧光发射值,这与先前胞内合成的CpcB光谱特征完全一致[6]。取纯化前后的胞外培养基进行SDS-PAGE电泳。如图4所示,由于对培养基中CpcB的纯化不佳,存在非特异性条带,但纯化前后培养基能看出在20 kDa左右有条带。凝胶锌离子染色后,纯化前CpcB条带有较弱的荧光,纯化后CpcB条带有较强的荧光。这些结果都表明分泌到胞外培养基中的CpcB的光谱特征和分子质量与胞内合成的一致[6]。
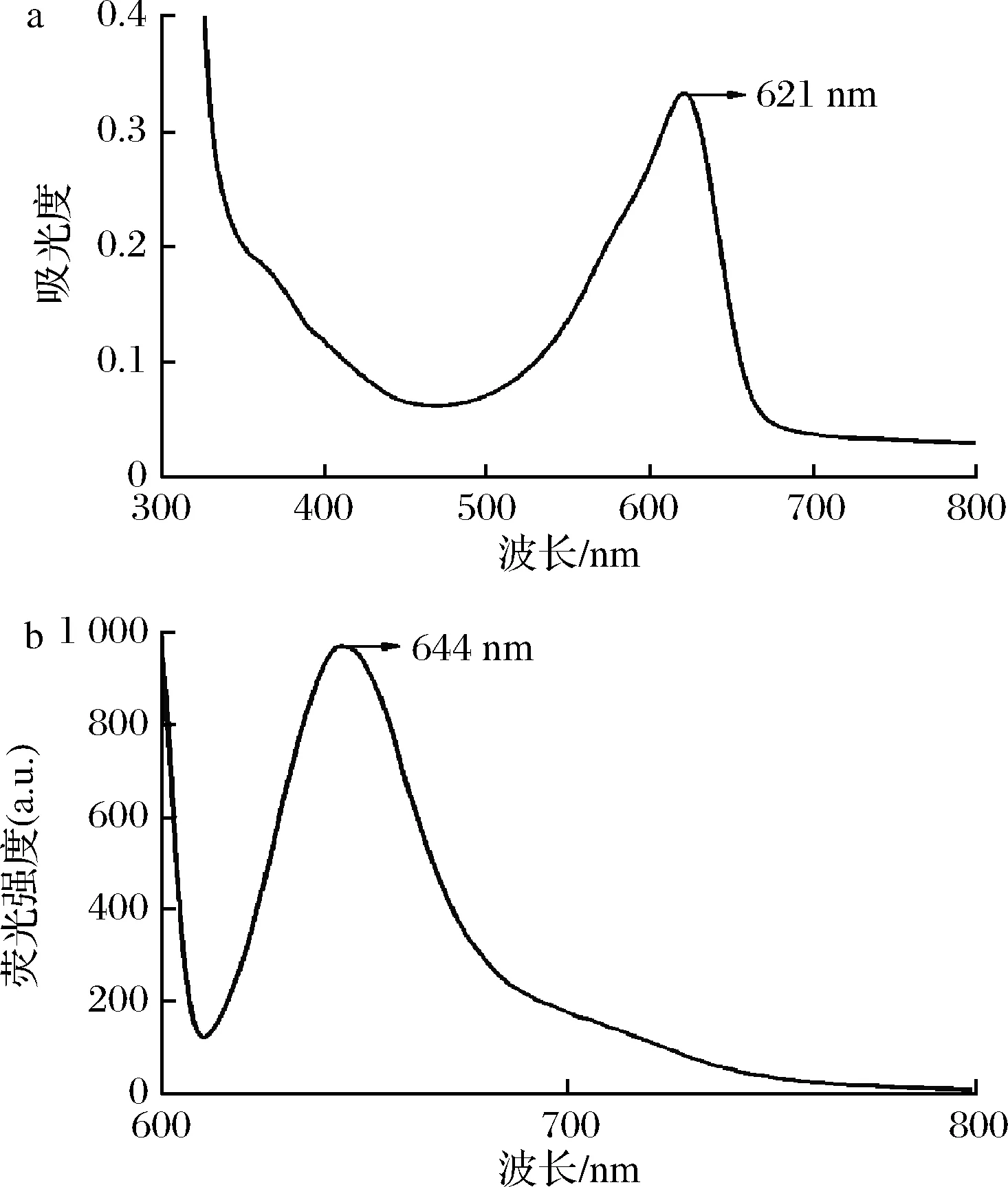
a-胞外培养基中CpcB纯化后吸收光谱;b-胞外培养基中 CpcB纯化后荧光发射光谱
图3 纯化胞外培养基CpcB的吸收光谱和荧光发射光谱
Fig.3 Absorption spectrum and fluorescence emission spectrum of the purified extracellular CpcB in medium
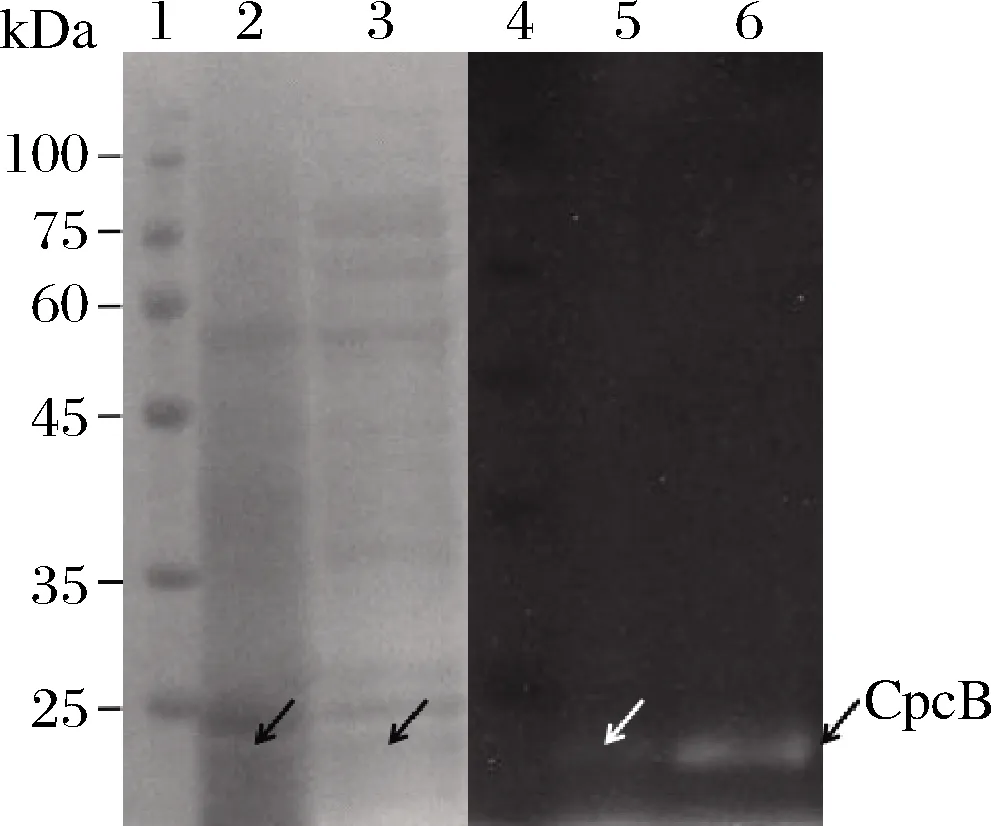
1、 4-蛋白质marker;2-培养基中CpcB纯化前考马斯亮蓝染色; 3-培养基中CpcB纯化后考马斯亮蓝染色;5-培养基中CpcB纯化前 锌离子染色;6-培养基中CpcB纯化后锌离子染色;箭头指示CpcB位置
图4 胞外培养基CpcB纯化前后的SDS-PAGE和 锌电泳分析
Fig.4 Analysis of the extracellular CpcB in medium before and after purification by gradient SDS-PAGE and chromoprotein Zn2+electrophoresis
2.2 乳糖作为诱导剂对CpcB胞外分泌的影响
基于T7为启动子的表达系统除了可以用IPTG,还可以用乳糖诱导表达。为了验证乳糖诱导重组藻蓝蛋白的表达及胞外分泌情况,按照实验方法1.3.1在25 ℃添加乳糖表达藻蓝蛋白。结果如图5-a所示,表达24 h后,与没有添加乳糖相比,添加乳糖的重组体系培养基中CpcB的荧光强度显著增加,当乳糖的质量浓度为10 g/L时,培养基CpcB的荧光强度达到最大值,是没加乳糖时的5.1倍。10 g/L的乳糖是诱导CpcB胞外分泌的最佳浓度,这与先前的乳糖诱导胞外脱乙酰几丁质酶最佳浓度一致[14]。如图5-b所示,添加乳糖后细胞OD600值也有少量增加,乳糖促进细胞的生长。乳糖无毒廉价、不抑制细胞生长、不影响分泌通道的形成,甚至可以为细胞生长提供碳源,这有益于可溶性蛋白的表达。先前的研究也表明比起IPTG,乳糖是蛋白异源表达和胞外分泌更好的诱导剂[14, 19]。
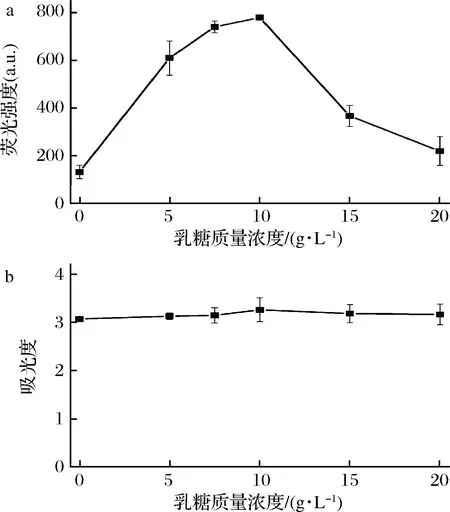
a-乳糖对荧光强度的影响;b-乳糖对细胞生长的影响
图5 乳糖诱导对胞外培养基CpcB荧光强度的影响
Fig.5 Effect of lactose on the fluorescence intensity of the extracellular CpcB in medium
2.3 培养基添加剂对CpcB胞外分泌的影响
通过补充培养基添加剂改变细胞膜通透性或增强周质空间的渗透压是增加外源蛋白胞外分泌的重要途径[13]。为了解培养基添加剂对藻蓝蛋白胞外分泌的影响,按照实验方法1.3.1分别向培养基中添加不同浓度的甘氨酸、Triton X-100、SDS和Tween-80等化学物。当添加甘氨酸时,结果如图6-a所示,甘氨酸显著增加胞外培养基CpcB的荧光强度。当甘氨酸的终质量浓度为25 g/L时,培养基荧光强度最高,是未加甘氨酸的5.1倍。甘氨酸作为增强重组蛋白胞外分泌的有效添加剂被广泛研究,如ZOU等[19]通过加入10 g/L的甘氨酸将胞外支链淀粉酶的活性提高8.3倍;HUANG等[14]加入1%的甘氨酸使得胞外脱乙酰几丁质酶活性增加2.4倍;VON ROMAN等[20]用20 g/L的甘氨酸和180 mmol/L的Tris使得SpA产量增加40倍。其机制是重组蛋白胞内合成后部分被迁移到周质空间,细胞质外膜为平衡压力释放少量的蛋白到培养基中,而甘氨酸插入肽聚糖、破坏细胞外膜的完整性,强化重组蛋白从周质空间到培养基的分泌[21-22]。表面活性剂Triton X-100和SDS也被认为是改善重组蛋白胞外分泌的有效添加剂[15, 23]。但在本研究中,添加Triton X-100和SDS到培养基中,结果如图6-b、图6-c所示,培养基CpcB的荧光强度并没有明显增强。Triton X-100对外膜的影响机制是破坏脂质双层膜[22,24];而SDS抑制脂肪酸的合成,导致膜磷脂合成不足[23]。故Triton X-100和SDS对细胞膜的破坏比甘氨酸要强很多,这可能是造成它们不如甘氨酸有效的重要原因。另外,荧光藻蓝蛋白的合成需要PCB和脱辅基蛋白CpcB的结合,推测Triton X-100和SDS有可能干扰CpcB和PCB的连接,但这需要进一步的证实。少量的Tween也能增强膜的通透性。刘军等[25]报道添加适量的Tween-80可提高胞外蛋白酶的酶活。本研究添加体积分数为1.5%的Tween-80到培养基中,如图6-d所示,培养基CpcB荧光强度增加2.5倍。重组藻蓝蛋白胞外分泌不仅受到单个因素的影响,各因素之间也相互影响,特别是不同添加剂之间的协同效应[15]。进一步的研究可对表达温度、乳糖、甘氨酸及Tween-80等因素进行协同考虑,获得最优的表达条件,高效地提升藻蓝蛋白的胞外分泌。
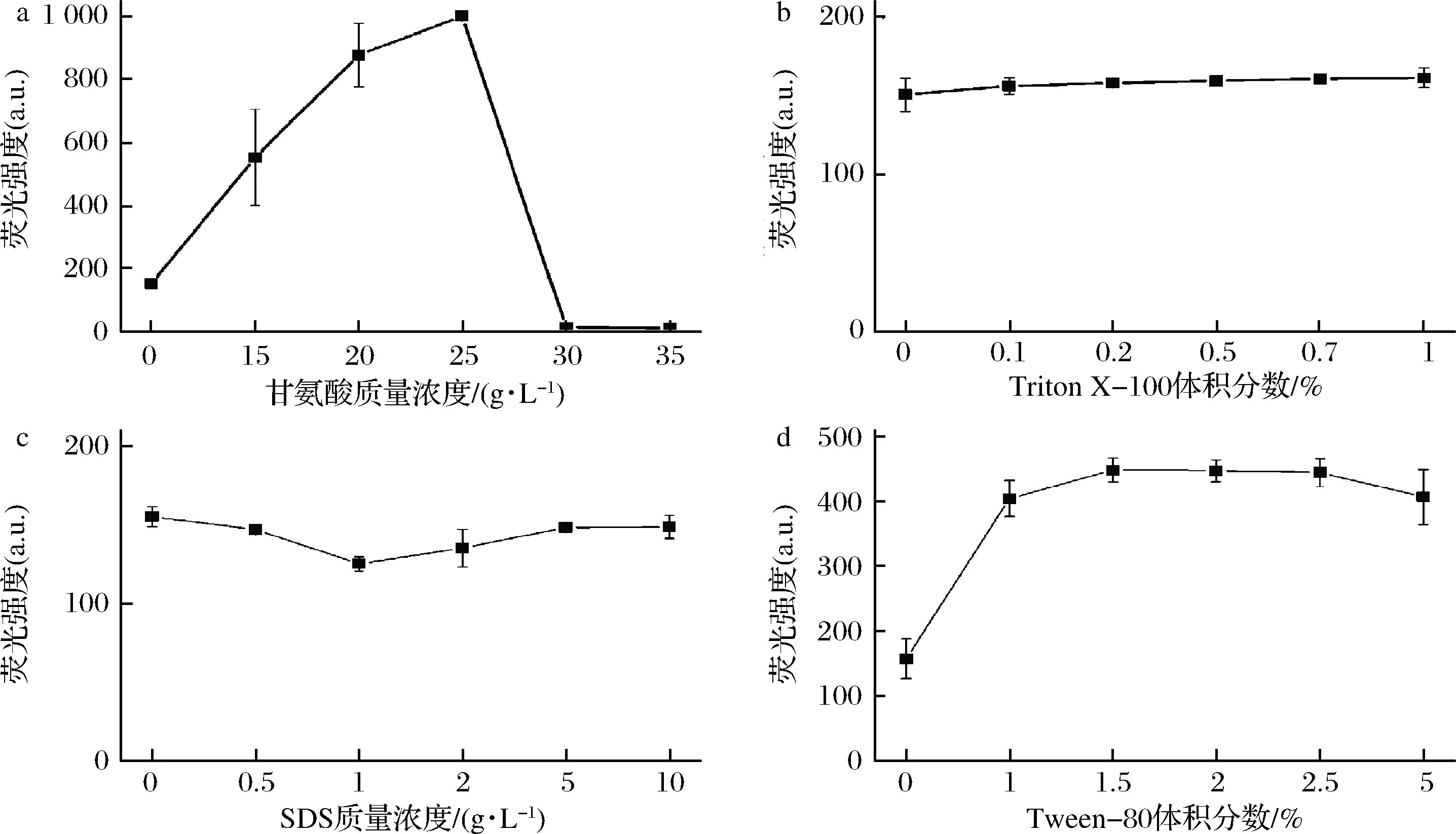
图6 培养基添加剂对胞外CpcB荧光强度的影响
Fig.6 Effect of medium additives on the fluorescence intensity of the extracellular CpcB
3 结论
藻蓝蛋白是被广泛报道的生物活性物质。为促使大肠杆菌重组藻蓝蛋白β亚基CpcB分泌到胞外培养基中,对温度、诱导剂和培养基添加剂等因素研究后发现,25 ℃未加诱导剂的条件下,CpcB表达24 h胞外培养基展现CpcB的荧光,藻蓝蛋白能分泌到培养基;加入IPTG诱导或者在16 ℃或37 ℃的条件下,胞外培养基没有明显的荧光;添加10 g/L的乳糖诱导表达,胞外培养基CpcB的荧光强度提高5.1倍;添加25 g/L的甘氨酸改变细胞膜的通透性可使胞外培养基CpcB的荧光强度提高6.3倍,添加体积分数为1.5%的Tween-80胞外CpcB荧光强度也增加2.5倍,而Triton X-100和SDS并没有改善荧光CpcB的胞外分泌。该研究为促进重组藻蓝蛋白胞外分泌,提高藻蓝蛋白异源生产的效率提供参考。
猜你喜欢
安徽建筑大学学报(2022年4期)2022-09-27
江苏卫生保健(2022年5期)2022-05-24
昆明医科大学学报(2021年4期)2021-07-23
生物化工(2020年5期)2020-10-30
农业环境科学学报(2020年9期)2020-10-09
国外畜牧学·猪与禽(2018年10期)2018-05-14
国外畜牧学·猪与禽(2018年8期)2018-05-14
国外畜牧学·猪与禽(2018年7期)2018-05-14
恋爱婚姻家庭·养生版(2016年5期)2016-05-06
人间(2015年11期)2016-01-09
