
采后硝普钠处理对生姜根茎能量代谢的影响
2020-07-08 07:09李灿婴魏美林葛永红陈延儒杨轶为励建荣
中国食品学报 2020年6期
李灿婴 魏美林 葛永红 陈延儒 杨轶为 励建荣
(渤海大学食品科学与工程学院 辽宁省食品安全重点实验室生鲜农产品贮藏加工及安全控制技术国家地方联合工程研究中心 辽宁锦州121013)
生姜,别名白姜,是多年生单子叶草本植物姜科姜属(Zingiber officinale Roscoe)姜的新鲜根茎[1], 是我国首批公布的药食两用植物资源之一。新鲜生姜根茎肉质肥厚,具特殊的香辛味,不仅富含碳水化合物、蛋白质、膳食纤维、姜油酮、姜酚和姜精油等营养成分,而且在降血脂、抗氧化和抗衰老等方面具有显著的功效[2]。生姜虽是一种非呼吸跃变型蔬菜,但刚采收的生姜表皮易脱落,根茎易失水老化,呼吸代谢旺盛,营养成分损失率高,导致根茎品质变劣,经济价值和食用品质降低[3]。 已有研究报道, 果实的衰老与线粒体能量代谢密切相关,并且能量亏缺会导致线粒体功能下降,从而削弱生物体对逆境的适应能力, 导致生理紊乱和病害发生[4-7]。
硝普钠(sodium nitroprusside,SNP)作为外源NO 供体,进入细胞后会在一定时间内通过还原反应自发分解并释放NO[8]。有研究表明,SNP 能够保持香蕉[9]、枇杷[10]、枸杞[11]、莲雾[12]、茄子[13]和芒果[14]等果蔬采后品质,延迟番茄乙烯生成[15],提高桃[16]和香蕉[17]等果实采后抗冷能力。 外源SNP 对生姜根茎采后贮藏过程中线粒体能量代谢与根茎衰老的影响尚未见研究报道。
本文以生姜根茎为试材, 采后用SNP 溶液浸泡处理, 研究常温贮藏过程中线粒体能量代谢相关 酶H+-ATPase、Ca2+-ATPase、 琥珀酸脱氢酶(SDH)、细胞色素氧化酶(CCO)活性及ATP、ADP、AMP 含量和能荷的变化,以期揭示常温贮藏过程中生姜根茎线粒体能量代谢与衰老的关系, 为阐明生姜根茎衰老的机理提供理论依据。
1 材料与方法
1.1 材料与试剂
供试生姜根茎, 采自辽宁省北镇市露地栽培大田, 愈伤后的生姜纸箱包装运抵实验室常温放置。选取适宜成熟度,无病虫害,无机械损伤,大小和色泽基本一致的生姜待用。
硝普钠(SNP)(分析纯),西亚试剂;5′-三磷酸腺苷钠盐(ATP)、5′-二磷酸腺苷钠盐(ADP)、5′-单磷酸腺苷钠盐(AMP),Sigma-Aldrich 公司;2,6-二氯酚靛钠 (DCPIP),Sigma-Aldrich 公司; 乙腈(色谱纯),美国天地公司;无机磷试剂盒,南京建成生物工程研究所;琥珀酸脱氢酶(SDH)试剂盒,南京建成生物工程研究所; 其它化学试剂均为分析纯,国药集团化学试剂有限公司。
1.2 仪器与设备
Agilent 1260 型高效液相色谱仪,美国安捷伦公司;UV-2550 型分光光度计,日本岛津公司;H-1650R 型离心机,长沙湘仪仪器有限公司。
1.3 方法
1.3.1 处理方法 采收的生姜用清水洗净表面的泥土后平摊在实验台上室温静置晾干。 将姜块随机分成2 组,室温下分别用去离子水(对照)和0.25 mmol/L 的SNP(前期预实验筛选)溶液(含0.05% Tween-20)浸泡10 min,取出自然晾干,装入塑料篮后放置在恒温恒湿培养箱中(20 ℃±1 ℃,RH 90%~95%)待用,每处理210 块生姜根茎。
1.3.2 取样方法 SNP 处理组和对照组生姜根茎分别于处理后0,3,6,9,12,15,18 d, 取皮下1~5 mm 处根茎组织,混合后切碎,锡箔纸包裹,经液氮冷冻后放入-80 ℃冰箱保存待用。
1.3.3 指标测定
1)线粒体的提取 参照文献[18]的方法并修改。 取25 g 冷冻组织用液氮研磨成粉, 加入10 mL 含0.25 mmol/L 蔗 糖、0.3 mol/L 甘 露 醇、1.0 mmol/L EDTA、 质量分数分别为0.1% BSA、0.5%PVP、0.1%半胱氨酸的50 mmol/L Tris-HCl(pH 7.5)缓冲液,继续研磨成匀浆。滤液在4 ℃、4 000×g 条件下离心10 min。 取上清液在4 ℃,14 000×g条件下离心20 min, 将沉淀用10 mL 缓冲液(10 mmol/L Tris-HCl pH 7.2, 内含0.25 mol/L 蔗糖、0.3 mol/L 甘露醇、1 mmol/L EDTA、0.1% BSA)洗涤,4 ℃、14 000×g 条件下离心20 min,再取上清液离心分离,得到的沉淀即为线粒体,加入1.5 mL悬浮液 (10 mmol/L pH 7.2 Tris-HCl, 内含0.25 mol/L 蔗糖、0.3 mol/L 甘露醇、1.0 mmol/L EDTA)悬浮,即为线粒体制备液。
2)H+-ATPase 活性测定 参照文献[18]的方法, 用南京建成生物工程研究所生产的H+-ATPase 试剂盒测定反应释放的无机磷含量。 以每小时每毫克蛋白的ATP 酶分解ATP 产生1 μmol 无机磷的量表示H+-ATPase 活性, 表示为U/mg 蛋白。
3)Ca2+-ATPase 活性测定 参照文献[18]的方法, 用南京建成生物工程研究所生产的Ca2+-ATPase 试剂盒测定反应释放的无机磷含量。 以每小时每毫克蛋白的ATP 酶分解ATP 产生1 μmol无机磷的量表示Ca2+-ATPase 活性, 表示为U/mg蛋白。
4)琥珀酸脱氢酶 (succinate dehydrogenase,SDH)活性测定 使用南京建成生物工程研究所生产的琥珀酸脱氢酶测试盒测定。 以每毫克蛋白每分钟使反应体系吸光度降低0.01 为1 个比活性单位,表示为U/mg 蛋白。
5)细胞色素氧化酶 (cytochrome oxidase,CCO)活性测定 参照文献[18]的方法。 取线粒体制备液0.1 mL, 加入质量分数为0.04%的细胞色素C 水溶液0.2 mL,重蒸水2 mL。 将试样在37 ℃预热2 min,加入0.5 mL,质量分数为0.4%的二甲基对苯二胺溶液,保温3 min 至出现红色,颜色在10 min 内稳定,于510 nm 比色,以每毫克蛋白每分钟吸光值变化0.1 为1 个活性单位, 结果以U/mg 蛋白表示。
6)ATP、ADP 和AMP 含量测定 参照文献[19]的方法。 采用HPLC 法测定ATP、ADP 和AMP的含量,色谱条件为C18色谱柱(ODS,250 mm×4.6 mm),检测波长为254 nm,流动相为乙腈(A)和磷酸钾缓冲液(B)(0.04 mol/L KH2PO4和0.06 mol/L K2HPO4混合液,pH 7.0,0.45 μm 滤膜过滤)。 线性梯度洗脱,时间7 min,延迟3 min,流速为1.2 mL/min,流动相A 的体积分数为75%~100%,流动相B 的体积分数为0~25%,进样体积为20 μL。 标准样品ATP、ADP 和AMP 购自Sigma 公司,配制0~200 mg/mL 的混合溶液制作标准曲线。
能荷 (EC)计算公式:EC=([ATP] + 0.5×[ADP])/([ATP] + [ADP] + [AMP])。
1.4 数据分析
所有指标测定进行3 次生物学重复, 数据处理采用SPSS 19.0 进行Ducan’s 多重比较分析(P<0.05),采用Microsoft excel 作图。
2 结果与分析
2.1 采后SNP 处理对生姜根茎线粒体H+-ATPase 活性的影响
H+-ATPase 是植物中的一种ATPase 质子泵。一对ATP 水解出质子转运出细胞,从而形成一个电化学质子通道用于营养物质的转运[17]。 由图1可知, 采后生姜根茎H+-ATPase 活性随贮藏时间延长逐渐降低, 但SNP 处理组生姜根茎H+-ATPase 活性在整个贮藏期间均高于对照组, 说明SNP 处理能很好的维持生姜根茎的H+-ATPase 活性。 在常温贮藏第9 天时,H+-ATPase 活性为对照组的1.33 倍; 在贮藏第18 天时,SNP 处理组H+-ATPase 活性为对照组的1.41 倍。
2.2 采后SNP 处理对生姜根茎线粒体Ca2+-ATPase 活性的影响
Ca2+-ATPase 是激发ATP 水解的初级钙离子转运蛋白。 Ca2+-ATPase 能保持钙稳态,从而延缓衰老[20]。线粒体H+-ATPase 和Ca2+-ATPase 活性对保持线粒体结构稳定和维持正常呼吸代谢具有重要作用。 由图2 可知, 整个贮藏过程中生姜根茎Ca2+-ATPase 活性总体呈下降趋势,SNP 处理显著(P<0.05)提高了生姜根茎Ca2+-ATPase 活性。在第3 天时,SNP 处理的生姜根茎Ca2+-ATPase 活性最高, 是对照组的1.81 倍。 对照组生姜根茎Ca2+-ATPase 活性始终呈缓慢下降趋势。 在贮藏第18天时,Ca2+-ATPase 活性为0.29 U/mg 蛋白,仅为第0 天时的44.8%。
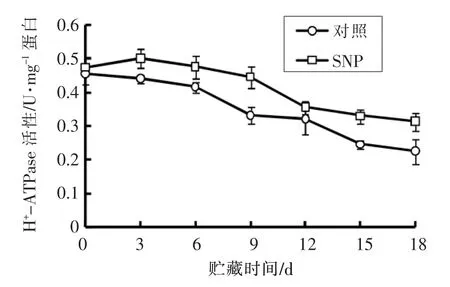
图1 采后SNP 处理对生姜根茎线粒体H+-ATPase活性的影响Fig.1 Effects of SNP treatment on the activity of mitochondrial H+-ATPase of ginger rhizomes

图2 采后SNP 处理对生姜根茎线粒体Ca2+-ATPase活性的影响Fig.2 Effects of SNP treatment on the activity of mitochondrial Ca2+-ATPase of ginger rhizomes
2.3 采后SNP 处理对生姜根茎线粒体SDH 活性的影响
SDH 是三羧酸循环(TCA)和呼吸链上的一个重要酶,催化琥珀酸氧化生成延胡索酸,是连接氧化磷酸化与电子传递的枢纽之一[17]。 贮藏过程中,对照组生姜根茎SDH 活性在整个贮藏期间呈缓慢下降趋势。而SNP 处理组生姜根茎SDH 活性随贮藏时间的延长呈先缓慢上升后下降的趋势,并在第9 天达到最高峰,SDH 活性是对照组1.42倍,随后SDH 活性随贮藏时间延长逐渐降低(图3)。说明SNP 处理生姜根茎在贮藏中后期有利于维持较高的SDH 活性, 从而有利于维持ATP 的合成能力,延缓生姜根茎衰老。
2.4 采后SNP 处理对生姜根茎线粒体CCO 活性的影响
CCO 是线粒体电子传递链末端氧化酶, 能够将电子从细胞色素C 传递给O2,以ATP 的形式为植物的多种生理活动提供能量[17]。 由图4 可知,SNP 处理组生姜根茎CCO 活性在整个贮藏期间均显著高于对照组。第3 天时,SNP 处理组和对照组生姜根茎CCO 活性降至最低,但SNP 处理的生姜根茎CCO 活性是对照的1.25 倍。在第12 天前,SNP 处理组和对照组生姜根茎CCO 活性变化趋势都比较平缓,呈缓慢上升趋势,表明这期间根茎内线粒体处于比较好的功能状态。 到第15 天时,SNP 处理组和对照组生姜根茎CCO 活性均达到最大,SNP 处理组的CCO 活性是对照组的1.28倍。
2.5 采后SNP 处理对生姜根茎ATP、ADP 和AMP 含量及能荷的影响

图3 采后SNP 处理对生姜根茎线粒体SDH 活性的影响Fig.3 Effects of SNP treatment on mitochondrial SDH activity of ginger rhizomes

图4 采后SNP 处理对生姜根茎线粒体CCO 活性的影响Fig.4 Effects of SNP treatment on mitochondrial CCO activity of ginger rhizomes
随贮藏时间延长,生姜ATP 含量呈逐渐下降的趋势,SNP 处理组生姜根茎ATP 含量在贮藏期第3~18 天内显著高于对照组。 在常温贮藏第18天时,SNP 处理组生姜根茎ATP 含量是对照组的1.96 倍(图5a)。SNP 处理组生姜根茎ADP 含量显著高于对照组,但SNP 处理组和对照组ADP 含量在采后贮藏过程中随贮藏时间延长逐渐降低,贮藏第18 天时,SNP 处理组生姜ADP 含量是对照组的1.28 倍(图5b)。 对照组和SNP 处理组生姜AMP 含量随贮藏时间延长呈逐渐上升趋势,但SNP 处理延缓了生姜AMP 含量的上升, 贮藏第18 天时SNP 处理组生姜AMP 含量仅为对照组的66.5%(图5c)。 SNP 处理组生姜能荷随贮藏时间延长逐渐降低但降低幅度趋平缓, 但对照组生姜能荷下降速度较快, 贮藏第18 天时,SNP 处理组和对照组生姜比贮藏第0 天下降了4.1%和13.7%(图5d)。
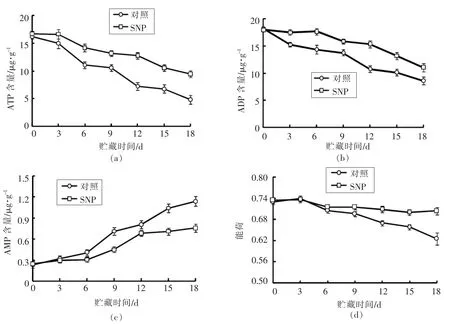
图5 采后SNP 处理对生姜根茎ATP(a)、ADP(b)、AMP(c)含量及能荷(d)的影响Fig.5 Effects of SNP treatment on the content of ATP (a), ADP (b), AMP (c)and energy charge (d)of ginger rhizomes
3 讨论
果蔬采后贮藏过程中水分蒸腾、 呼吸作用和细胞壁物质降解等会引起果实品质下降、 组织衰老以及ATP 合成能力逐渐降低,从而导致线粒体功能下降,进而引起果蔬的代谢及生理功能紊乱[4,21]。已有研究表明,病原菌侵染、衰老和生理紊乱与细胞能量状态密切相关[5,22-23]。 提高细胞ATP含量和能荷能够延缓荔枝、桃、梨等果实的衰老,并且提高果实抗病性[19,24-25]。 本研究发现,采后SNP 处理维持了贮藏期间生姜根茎较高的ATP水平和能荷水平,从而延缓了根茎的衰老速度。采后诱抗剂处理延缓了枇杷、芒果、油、桃和南果梨果实ATP 含量的下降并维持了较高的能荷水平,从而延缓果实衰老和提高抗病性[19,26-29]。此外,外源提供ATP 可减轻病害的发生,并且维持较高的能荷水平[5,24]。而氧化磷酸化解偶联剂2,4-二硝基苯酚(DNP)处理通过降低能量代谢而加重病害的发生[30]。 由此表明,采后诱抗剂处理延缓果实的衰老和提高抗胁迫能力与维持细胞较高能量水平相关。
H+-ATPase 和Ca2+-ATPase 是位于线粒体内膜上的主要呼吸代谢酶,负责合成和提供能量[31]。本研究发现,生姜根茎线粒体H+-ATPase 和Ca2+-ATPase 的活性随贮藏时间延长逐渐下降,但SNP处理延缓了生姜根茎H+-ATPase 和Ca2+-ATPase的活性的下降。 本结果与南果梨、桃、芒果等果实中的研究结果一致[19,26,28]。 由此表明,SNP 处理可以通过提升能量代谢水平来保护细胞膜完整性和离子选择渗透性,得以延缓生姜根茎衰老。SDH 是TCA 循环过程中的关键酶之一, 位于线粒体内膜上,也是线粒体的标志酶,其活性在一定程度上反应了线粒体的功能[20]。 CCO 位于线粒体电子传递链上,是呼吸链的末端氧化酶,将电子从细胞色素C 传递给O2,为氧化磷酸化提供能量[32]。 本研究发现,SNP 处理提高了贮藏过程中生姜根茎SDH 和CCO 的活性。 这与秋水仙碱对莲雾贮藏过程中SDH 和CCO 活性的影响变化趋势一致[32]。 本试验中SNP 处理均提高了同一时期内4 种酶的活性,维持了线粒体良好的功能状态进而延缓了生姜根茎的衰老。
4 结论
SNP 处理提高了采后生姜根茎线粒体H+-ATPase、Ca2+-ATPase、SDH 和CCO 的活性,延缓了生姜体内ATP、ADP 含量及能荷的下降,使生姜根茎维持较高的能量水平, 从而有效延缓生姜根茎采后衰老进程,更好地保持生姜根茎的品质。
猜你喜欢
医学研究生学报(2022年5期)2022-12-07
中华实用诊断与治疗杂志(2022年1期)2022-08-31
保健医苑(2021年7期)2021-08-13
海洋通报(2021年1期)2021-07-23
今日农业(2020年13期)2020-08-24
医药前沿(2019年35期)2019-01-05
文苑·经典美文(2017年7期)2017-07-26
中央民族大学学报(自然科学版)(2017年3期)2017-06-11
中成药(2017年3期)2017-05-17
短篇小说(2014年11期)2014-02-27
