
喀斯特天坑内孔雀藓属蒴齿亚显微结构研究
2020-07-14 09:03张朝晖王智慧
植物研究 2020年4期
吴 金 张朝晖* 王智慧
(1.贵州师范大学贵州省山地环境信息系统与生态环境保护重点实验室,贵阳 550001;2.贵州师范大学生命科学学院,贵阳 550001)
喀斯特天坑(Karst Tiankeng)是发育于特定岩溶地质、气候和水文环境下的喀斯特地貌[1],其坑口直径及深度均大于100 m。在喀斯特天坑内部,由于其独特的生态环境,具有生物避难所效应[2~4]。
苔藓植物是喀斯特天坑环境中优势类群之一[5],其结构简单,适应能力强。在当前喀斯特天坑内苔藓植物的研究中,李小芳、吴金等[5~6]报道了喀斯特天坑微环境对苔藓植物多样性具有生物避难所功能。但对于苔藓植物的主要繁殖结构——蒴齿结构在喀斯特天坑微环境的特征未见报道。
蒴齿是生长于苔藓植物孢蒴上的一种重要繁殖结构,其主要功能是促进孢子散播[7]。藓类蒴齿研究可追溯至18世纪末至19世纪初[8]。而根据现有研究报道藓类植物蒴齿的结构特征可作为种间分类的重要依据之一[9]。19世纪末,Philiber在前人研究的基础上对蒴齿的发育方式进行了较深入的研究[10],进一步丰富了蒴齿的特征及应用。为了更好地了解蒴齿结构特点以期对其充分利用,Bruch[11]、Schnepf[12]、Grout[13]、Patterson[14]、Lazarenko[15]等对蒴齿的水湿运动进行了探索,从侧面验证了蒴齿与藓类的繁殖有重要的关系。相对而言,国内关于藓类蒴齿的研究历程与国外相似,也经历了从作为藓类孢蒴的一个附属结构发展成为对藓类种类鉴别、分类和系统进化发育的鉴定依据之一[16~18]。但就目前对蒴齿的研究而言,无论是国外的研究还是国内的研究更多的是将其作为分类依据以及系统发育位置的判定[19~27],而忽略了蒴齿在生态学和环境学方面意义的研究,导致这一领域的研究成果寥寥,国内仅王晓蕊等对蒴齿与环境的相互适应机理进行了研究[28]。
经笔者多次考察及研究发现,孔雀藓属植物在贵州猴耳天坑中生境分布广泛[6],如岩面、树干和土面。随着对苔藓植物蒴齿亚显微形态结构更为详细的研究,尤其是借助扫描电镜等大型设备来寻找有用的分类指标已经成为现代苔藓学家重要的研究手段[29]。目前,国内对孔雀藓属植物蒴齿的研究仅有零星记载[6,30],为了更进一步的明确蒴齿亚显微形态结构与喀斯特天坑特殊生境的关系,探讨蒴齿结构作为系统分类的有效性。本文以孔雀藓属植物蒴齿为研究对象,利用电子显微镜扫描法,对喀斯特天坑内孔雀藓属蒴齿亚显微结构进行观察,为喀斯特天坑微环境下藓类植物蒴齿结构特征研究提供参考。
1 材料与方法
1.1 研究区域概况
猴耳天坑,位于贵州省贵阳市开阳县城关镇石头村桃子窝村民组(开阳县北部),地理坐标介于106°59′56.5″~107°00′16.6″E,27°04′57″~27°05′27″N,距离开阳县城7 km,因其坑口形似猴耳,故名“猴耳天坑”。坑口直径300 m,坑深280 m,坑底最大直径280 m,属大型塌陷型天坑(属标准竖井式天坑)[6]。
1.2 野外调查与样点设置
作者于2017~2019年对猴耳天坑进行实地调查发现,该天坑中孔雀藓属植物极其丰富,其大小在天坑内呈现各异,沿坑口至坑底分别设置了10个采样点。本研究采用线路与典型样地取样法相结合的方法[31],从坑口至坑底在有典型孔雀藓属植物的生境中设置一个10 m×10 m的样地,在样地内按S型布点法设置5个1 m×1 m的样方,每个样方内再按5点取样法设置5个10 cm×10 cm的小样方,自制1 cm×1 cm的小样方(100个小格子,用于测量盖度)[6],采集小样方内全部孔雀藓(网格法)带回室内进行鉴定,标本存放于贵州师范大学苔藓植物标本室。
1.3 材料类型
选择猴耳天坑中具有成熟孢蒴的两种孔雀藓属植物进行蒴齿研究:①黄边孔雀藓(H.flavo-limbatum):植物体中等大小,黄绿色,着生于岩面、树干和土面上;②东亚孔雀藓(H.japonicum):植物体疏松丛集生,着生于腐木、树干、岩面或土面上;仪器设备主要包括(均由贵州师范大学贵州省山地环境信息系统与生态环境保护重点实验室提供):经纬度计(Etrex 20)、海拔计(Etrex 20)、照度计(PM6612L)、温湿度计(HT-635)、光学双目生物显微镜(XSZ-107TS)、双筒解剖镜(HWG-1)、高真空镀膜机(HUS-5GB)、扫描电子显微镜(JSM-6490LV)。
表1 猴耳天坑样地基本特征
Table 1 Basic characteristics of Monkey-Ear Tiankeng sample

采集地Collectingplace样地编号SampleNo.生境Habitat盖度Coverage(%)种名Species天坑上部(海拔1287m)UpperpartofTiankeng(Altitude1287m)S1石生Stone30东亚孔雀藓H.japonicumS2石生Stone30东亚孔雀藓H.japonicumS3石生Stone20黄边孔雀藓H.flavo-limbatumS4石生Stone80黄边孔雀藓H.flavo-limbatum天坑中部(海拔1147m)MiddlepartofTiankeng(Altitude1147m)S5腐木生Rottenwood40黄边孔雀藓H.flavo-limbatumS6石生Stone30东亚孔雀藓H.japonicumS7石生Stone20东亚孔雀藓H.japonicum天坑底部(海拔1007m)BottomofTiankeng(Altitude1007m)S8石生Stone10东亚孔雀藓H.japonicumS9石生Stone20黄边孔雀藓H.flavo-limbatumS10土生Nativesoil30黄边孔雀藓H.flavo-limbatum
1.4 实验方法与数据统计
将采集到的两种孔雀藓属植物的孢蒴经处理后使其保持在半湿润半干燥状态,分别置于解剖镜下将蒴帽拔掉,切下蒴齿,待获得蒴齿后,用镊子夹住蒴齿置于事先已经粘贴好双面胶的样品台上(供扫描电镜使用),将其置于HUS-5GB高真空镀膜机内喷碳镀膜,在JSM-6490LV扫描电子显微镜下进行观察,测量、拍照并记录蒴齿的外齿层数目(外齿长、外齿宽、节片数、横脊数、中脊数)、内齿层数目(齿毛、节瘤、齿条、穿孔)、基膜高度等数据。运用Excel 2013数据统计软件分别对两种孔雀藓属植物样点—蒴齿结构—环境因子数据矩阵表进行设计用于统计分析,将预先设计的矩阵表导入Canoco 5统计分析软件中,进行排序模型的选择,结果显示4个轴中梯度最大值超过4,因此,采用非约束性排序(间接梯度分析)分析中的单峰模型除趋势对应分析(DCA)进行排序分析,排序结果用物种—环境因子关系的二维排序图表示。
2 结果与分析
2.1 孔雀藓属两个种蒴齿的形态结构特征
2.1.1 黄边孔雀藓(H.flavo-limbatum)
内外齿层数均为16枚,外齿层背面均具“Z”字形的中脊,齿片近直立,窄披针形(见图1:A),外齿长160~435 μm,宽60~120 μm,内齿长479~573 μm,从背面观(见图1:B),外齿右侧节片为33~55枚,横脊33~50条,左侧节片为33~50枚,横脊33~54条,具密集且丰富疣状突起(见图1:C);从腹面观,横隔数34~52,狭窄且通常不饱满(见图1:D);内齿层表面光滑(见图1:E),具折叠基膜,基膜高约248 μm,多达外齿层长度的1/2,齿条腹面中部具长方形竖状结构,沿竖状结构往左右两侧具横纹方形结构(见图1:F),具穿孔(见图1:G),不具节瘤,齿条间具弱齿毛(见图1:H),长约394 μm。
2.1.2 东亚孔雀藓(H.japonicum)
内外齿层数均为16枚,外齿层背面均具“Z”字形的中脊,齿片伸入齿条间,长三角形(见图2:A),外齿长126~422 μm,宽68~134 μm,内齿长191~408 μm,从背面观(见图2:B~C),外齿右侧节片约32~50枚,横脊30~46条,左侧节片约31~44枚,横脊30~46条,具稀疏疣状突起(见图2:D);从腹面观,横隔数31~54,宽且通常饱满(见图2:E);内齿层表面光滑(见图2:F),基部常愈合形成基膜(愈合程度因种而异),基膜高约60 μm,低于外齿层长度的1/3。齿条腹面中部不具长方形竖状结构,整个腹面层由左右不规则细胞组成(见图2:G),具弱穿孔或缺失,具明显节瘤,齿条间具明显齿毛(见图2:H)。
2.2 孔雀藓属蒴齿结构与喀斯特天坑环境因子关系的除趋势对应分析(DCA)
蒴齿结构对环境的变化表现出明显的反应,如蒴齿的齿片和齿条会随着环境中水分、湿度的含量做张合运动、同时也会凭借环境湿度的大小而确定孢子的散发与否等。孔雀藓属蒴齿也不例外。本研究对野外采集到的黄边孔雀藓和东亚孔雀藓进行DCA分析,两种孔雀藓属植物与空气温度、空气湿度、光照度、海拔、盖度、人为干扰度以及林冠郁闭度7个环境因子关系的DCA结果如图3~4所示。2个排序轴的特征值分别为0.446,0.140和0.084,0.002,轴1和轴2的解释量分别为76.1%、100%和98.07%、99.88%,表明其对蒴齿结构关系的分析有较好的效果或说明排序结果是可信的。
由图3可知,黄边孔雀藓蒴齿的齿片长度、宽度与喀斯特天坑内温度、海拔、盖度、人为干扰度以及林冠郁闭度呈正相关,与光照和湿度呈负相关;横脊数、节片数、中脊数与湿度具有较强的正相关性,与其他呈负相关;湿度是影响节片数和中脊数最重要的环境因子,反之与光照的相关性较弱,这就说明低温高湿有利于蒴齿节片和中脊数的发育;天坑深度对黄边孔雀藓蒴齿外齿层齿片长度起决定性作用,阴暗潮湿的环境有利于黄边孔雀藓蒴齿外齿层齿片长度的增长,环境郁闭度是影响齿片宽度增长的重要环境因子;由图4可知,东亚孔雀藓蒴齿外齿层齿片长度和宽度与人为干扰、光照、温度呈正相关,受三者的共同影响,与林冠郁闭度、天坑深度、湿度、盖度呈负相关;节片数、横脊数、中脊数与湿度、盖度、人为干扰度呈正相关,其受湿度影响最大,其生境多为湿度大、土壤含水量高的环境。由此看来,齿片长度、宽度、节片数、横脊数以及中脊数是随着环境的变化而变化的特征,在喀斯特天坑内易受环境因子的影响和干扰,是可变的、不具分类价值的特征,换言之,在运用藓类植物蒴齿作为系统演化分类依据时要做性状的筛选。

图1 黄边孔雀藓蒴齿扫描电镜形态特征图 A.黄边孔雀藓蒴齿整体观;B.外齿层背面观;C.齿片背面观;D.齿片腹面观;E~F.内齿层(齿条)腹面观;G.穿孔;H.齿毛Fig.1 The morphological characteristic of peristome scanning electron microscope in H.flavo-limbatum A.Integral view of peristome of H.flavo-limbatum; B.Dorsal view of exstomium; C.Dorsal view of dentium; D.Ventral view of dentium; E-F.Ventral view of endostomium; G.Tresis; H.Cilia

图2 东亚孔雀藓蒴齿扫描电镜形态特征图 A.东亚孔雀藓蒴齿整体观;B~C.外齿层背面观;D.齿片背面观;E.齿片腹面观;F~G.内齿层腹面观;H.齿毛Fig.2 The morphological characteristic of peristome scanning electron microscope in H.japonicum A.Integral view of peristome of H.japonicum; B-C.Dorsal view of exstomium; D.Dorsal view of dentium; E.Ventral view of dentium; F-G.Ventral view of endostomium; H.Cilia
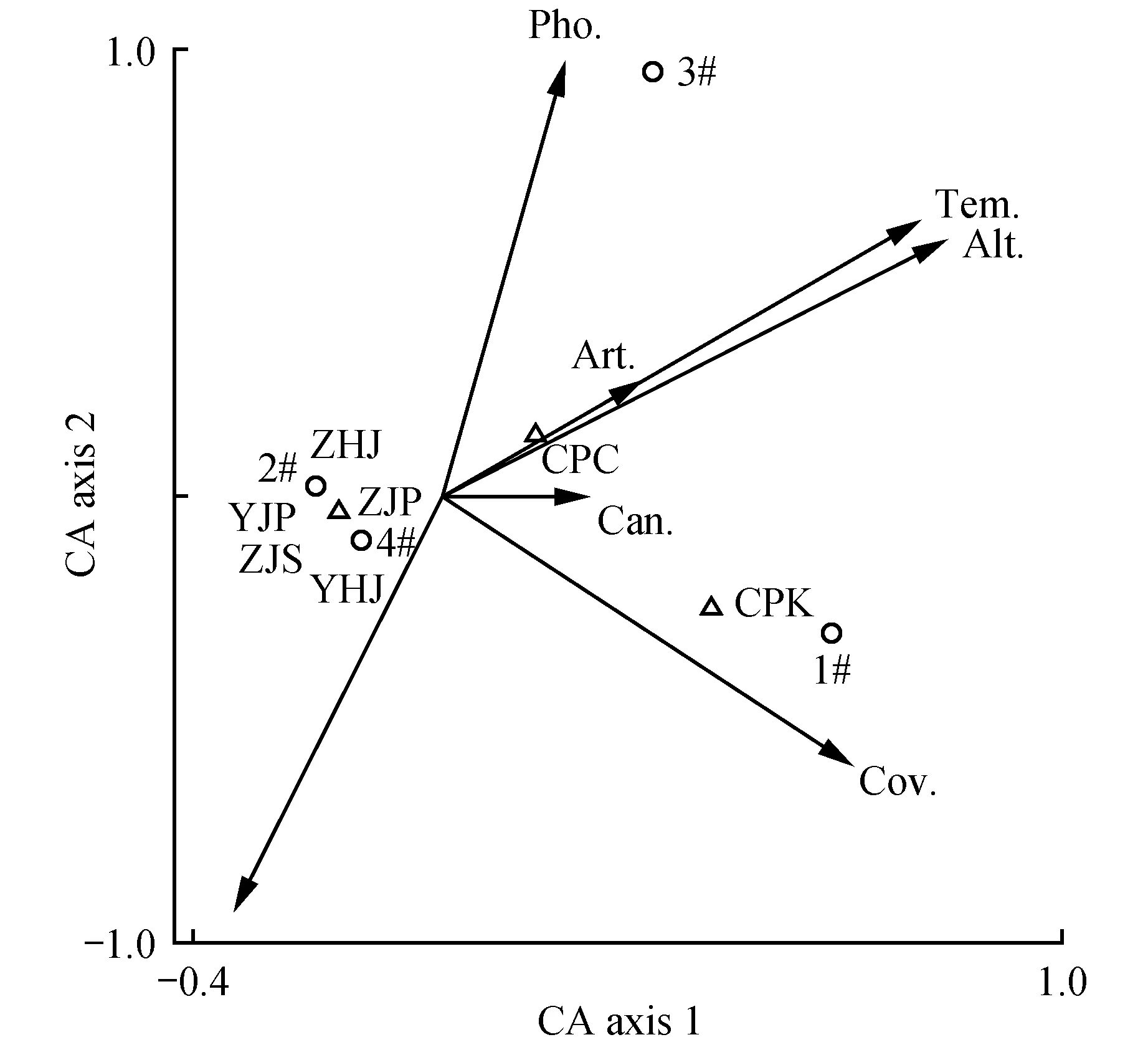
图3 黄边孔雀藓—环境因子DCA二维排序图 1#~4#表示采集到带有蒴齿的黄边孔雀藓(H.flavo-limbatum)数目CPC.齿片长度;CPK.齿片宽度;YJP.右节片数目;ZJP.左节片数目;YHJ.右横脊数目;ZHJ.左横脊数目;ZJS.中脊数目;Tem..空气温度;Hum..空气湿度;Pho..光照度;Alt..海拔;Cov..盖度;Art..人为干扰度;Can..林冠郁闭度 下同。Fig.3 Two-dimensional sequencing diagram of H.flavo-limbatum -environmental factor DCA 1-4 show the number of H.flavo-limbatum collected with peristome CPC. Dentium length; CPK. Dentium width; YJP. The number of right plates; ZJP. The number of left plates; YHJ. The number of right transverse raphe; ZHJ. The number of left transverse raphe; ZJS. The number of middle raphe; Tem.. Air temperature; Hum.. Air humidity; Pho.. Iilluminance; Alt.. Altitude; Cov.. Coverage; Art.. Artificial interference; Can.. Canopy density The same as below.

图4 东亚孔雀藓—环境因子DCA二维排序图 1#~4#表示采集到带有蒴齿的东亚孔雀藓(H.japonicum)数目Fig.4 Two-dimensional sequencing diagram of H.japonicum -environmental factor DCA 1-4 indicates the number of H.japonicum collected with peristome
4 讨论
近年来,随着相关学者对苔藓植物资源价值的认识,逐渐从配子体研究转向孢子体研究过渡到蒴齿亚显微形态结构研究,尤其是蒴齿表面的细节特征的研究在分类中是极其重要的[22]。本研究在扫描电子显微镜(SEM)下对孔雀藓属2种植物蒴齿的观察结果表明:2种植物蒴齿为双齿层,内外齿层数均为16枚,外齿层背面均具“Z”字形的中脊,纵隔不明显,密被细疣,具明显或不明显的节或具横脊。以上特征表明这属于两者的共性特征,是稳定的、不可变的特征,可作为高阶(如属级)的分类依据,但不可作为低阶分类单元的判定依据。而两种植物蒴齿在具有一定共性的同时,它们的表面特征在细节上存在显著的差异,如齿片长度、宽度等,会受到生境中环境因子的影响,因此,在运用藓类植物蒴齿作为系统分类依据时要做性状的筛选,换言之,蒴齿被用于系统分类的依据是可行的[32],但并不是蒴齿上所有的结构特征都可用于系统分类。
喀斯特天坑的形成造成了区域生境的异质性以及环境多样性,巨大的塌陷规模和相对封闭的环境使天坑内部形成一个独特的生境系统,为植物的生长繁殖发育提供了良好的栖息地。而蒴齿形态结构特征同苔藓植物孢子体的演化、对环境的适应及孢子的传播方式都有着极其密切的关系。在本研究中,基于蒴齿结构—环境因子的二维排序图,发现环境因子对蒴齿表面的细微结构特征存在显著的影响。首先,天坑深度对黄边孔雀藓蒴齿外齿层齿片长度起决定性作用;阴暗潮湿的环境有利于黄边孔雀藓蒴齿外齿层齿片长度的生长,环境郁闭度是影响齿片宽度生长的重要环境因子;其次,湿度是影响节片数和中脊数最重要的环境因子,而对光照的需求较低,即高温环境不利于蒴齿节片和中脊的发育,低温高湿利于蒴齿节片和中脊的发育;最后,东亚孔雀藓蒴齿外齿层齿片长度、宽度与人为干扰、光照、温度具有较强的相关性,受三者的共同影响,节片数、横脊数、中脊数与湿度、盖度、人为干扰度呈正相关,其受湿度的影响最大,其生境多为湿度大、土壤含水量高的环境。综上所述,导致2种孔雀藓属植物蒴齿结构呈现不同变化的原因是由于受到喀斯特天坑这种隐域且封闭圈地化生境的影响,其内部环境多为湿度大、温度低、降雨丰沛、热量充足,适宜植物生长发育繁殖。对于大多数苔藓来说,孢子的散播是由蒴齿控制的。当这些苔藓生境变潮湿时,齿片的张开程度各不相同[14]。究其原因,是因喀斯特天坑中封闭型隐域生境的形成使得天坑区别于周围地带的小气候和适合植物生存的独特微生境[33],为孔雀藓属植物蒴齿等结构的生长发育提供了条件。
在Vitt的研究中,对蒴齿的结构与微环境变化的适应做了详细的描述。依据Vitt的观点,苔藓植物在潮湿的环境中,其蒴齿高度发达(双层蒴齿),较为原始,而在干旱的环境中,其蒴齿简单(单层蒴齿),是进化的类群,这与本研究的结果是一致的[34],即潮湿的环境中,蒴齿呈现高度发达较为原始的双齿层。本文所观察的2种孔雀藓属植物多分布于热带亚热带地区,多属石灰岩型藓类,大多为石生、稀树生,其双层蒴齿是适应这种潮湿阴暗环境的适应性表现。同时,该研究表明了将藓类蒴齿作为系统间发育分类的重要指标的必要性和采用现代化手段研究方法的重要性,这将对苔藓植物的分类和系统演化有着重要意义。
5 结论
①两种植物蒴齿为双齿层,内外齿层数均为16枚,外齿层背面均具“Z”字形的中脊。2种孔雀藓属植物外齿层齿片曲向存在差异,黄边孔雀藓齿片近直立、呈窄披针形,东亚孔雀藓齿片伸入齿条间、呈长三角形;黄边孔雀藓外齿长度、宽度、内齿长度、节片数、横脊数与东亚孔雀藓均明显不相同;黄边孔雀藓齿条腹面中部具长方形竖状结构,沿竖状结构往左右两侧具横纹方形结构,不具节瘤,东亚孔雀藓齿条腹面中部不具长方形竖状结构,整个腹面层由左右不规则细胞组成,具明显节瘤;基膜高度:黄边孔雀藓多达外齿层长度的1/2,东亚孔雀藓基膜低于外齿层长度的1/3。②2种孔雀藓属植物蒴齿结构受湿度的影响最大,其次是温度和光照度,其生境多为湿度大、土壤含水量丰富的环境,而湿度主要是通过温度、光照、降雨和生境等环境条件来实现。③2种孔雀藓属植物蒴齿共性特征是稳定的、不可变的特征,可作为高阶科属的分类依据,但不可作为低阶分类单元的判定依据。而细微结构如齿片长度、宽度等,会受到生境中环境因子的影响,在运用藓类植物蒴齿作为系统分类依据时要做性状的筛选。
猜你喜欢
小哥白尼(野生动物)(2022年4期)2022-07-16
今日农业(2021年15期)2021-10-14
检察风云(2021年13期)2021-09-14
阅读与作文(小学高年级版)(2018年12期)2018-12-18
小天使·一年级语数英综合(2018年9期)2018-10-16
红领巾·萌芽(2017年5期)2017-06-23
爆笑show(2016年7期)2017-02-09
科学大众(中学)(2016年5期)2016-05-10
少儿科学周刊·儿童版(2015年10期)2015-11-07
少儿科学周刊·儿童版(2015年1期)2015-07-07
