
光照对晒烟变黄期叶绿素荧光及抗氧化指标的影响
2020-09-25 08:47向欢王俊陈勇汤卫荣张凡刘雷
中国烟草学报 2020年4期
向欢 ,王俊,陈勇,汤卫荣,张凡,刘雷
1 四川农业大学农学院,成都市温江区惠民路211号 611130;
2 四川省烟草公司德阳市公司,德阳市长江东路176号 618400
晒烟具有突出的香气风格特色和良好的应用价值[1-4]。已有大量研究表明晒烟调制措施对烟叶质量和风格特色有显著影响[5-9],但与之相关的晒烟调制生理研究明显较少[10-11]。光照是晒烟调制中必需的因素,也是晒烟与晾烟和烤烟在调制方法上最明显的差异因素[12-13];它不仅影响晒烟叶品质特色[6-9],还影响晒烟的叶绿素含量和衰老相关生理指标[10-11,14];然而调制期的光照与晒烟叶生理生化指标变化间的关系仍不清楚。
调制变黄期的晒烟叶处在光照、水分和温度等环境因素的逆境胁迫下,烟叶叶绿素荧光参数指标会因此受到影响。叶绿素荧光参数对光照、水分和温度等胁迫很敏感,通过测定叶绿素荧光参数可以实时、无损地了解叶片光合能力、光合系统能量传递状况以及叶片所受胁迫的程度[15];监测叶绿素荧光参数已成为研究植物逆境反应的重要方法[16]。目前,关于晒烟调制期光照对叶绿素荧光参数和抗氧化指标的影响以及这些指标变化之间的联系鲜见报道[17]。因此,本文测定了晒烟调制变黄期间不同光照射处理下晒烟的叶绿素含量、光合性能、叶绿素荧光参数、活性氧含量及抗氧化物酶活力,分析它们的变化规律和相互间的关系,以探索光照在晒烟调制变黄期的影响和作用机理。
1 材料与方法
1.1 材料与试剂
供试晒烟品种为“什烟1 号”,由四川省德阳市烟草公司提供。H2O2测定试剂盒(A064-1-1)产自南京建成生物工程研究所;各种常规化学试剂(AR)产自成都市科龙化工试剂厂。
1.2 实验方法
1.2.1 晒烟栽培和调制
实验在2018 年进行,地点为四川省什邡市师古镇大泉坑村,栽培晒烟的田间行距100 cm、株距45~50 cm。烟叶采收调制期在5~6 月,以适熟的中部烟叶为实验材料,烟叶采收标准为主脉呈现浅白色,叶片呈淡绿色至黄绿色。用钢管搭建晒烟调制棚架、用无色透明塑料薄膜覆盖棚顶,用多层黑色遮阳网遮盖棚顶部的一半作为遮光处理(CK)区域、另一半留作光照处理区域;悬挂黑色遮阳网将CK 区域围绕,以避免外来散射光的干扰。用LI-6400XT 便携式荧光仪测得晴天正午时棚下光照处理区域光合有效辐射约为300~350 μmol·m-2·s-1,CK 区域的光合有效辐射约为10~20 μmol·m-2·s-1。将晒烟按单叶编竿方法编好,烟竿平分为两组分别置于遮光处理区域和光照处理区域进行调制;其它调制条件和操作与当地晒烟生产习惯相同。实验期间天气状况以晴天为主。
1.2.2 荧光参数测定
在实验第1~7 d 内上午10:00 开始,以LI-6400XT(LI-COR,USA)便携式光合荧光仪配上6400-40 荧光叶室用于测定烟叶的PS Ⅱ实际光化学效率(PhiPS2)、非光化学淬灭(qN)、光化学淬灭(qP)、电子传递速率(ETR)、PSII 最大光化学效率(Fv/Fm)等荧光参数。每次测定时从两个处理下的中央区域各选取3~5 片烟叶,在每片烟叶的中段区域内选择以主脉为对称轴的左右2 个位点做重复测定。
1.2.3 生化指标测定
在实验第1~9 d 内,每天从两种处理中抽取已测过叶绿素荧光参数的烟叶,截取其中段区域(去除主叶脉)约50 g,经天平称重后分成约5 g /份、用锡箔纸包裹、编号并记录重量后放入液氮中保存,用于测定叶绿素、H2O2、MDA 的含量和CAT、POD 的酶活性。参考熊庆娥的方法测定叶绿素和MDA 含量以及POD 和CAT 酶活性[18]。使用南京建成生物工程研究所H2O2测定试剂盒(比色法)测定H2O2含量。使用UV-2600(Shimadzu, Japan)紫外-可见分光光度计测定以上各指标的吸光度。
1.3 数据分析
用Excel 2013 求数据均值、误差并制图,用DPS7.05 分析样本间差异显著性,用SIMCA-P13 软件做两处理间的正交偏最小二乘法判别分析(OPLS-DA)。
2 结果与分析
2.1 叶绿素含量分析
图1 显示:调制4 d 后CK 和光照处理的叶绿素含量分别比各自第1 d 时的含量降低了10%和15%,这表明在晒烟调制变黄初期的烟叶组织中仍然保有较多的叶绿素,具有接收光能的物质基础。在实验的第5~7 d 内两种处理下的叶绿素下降速都明显加快,到7 d 时CK 和光照处理的叶绿素含量已分别减少了57%和67%;由此可推测晒烟叶的光合能力将因叶绿素含量下降而降低。

图1 不同天数内CK 和光照处理下烟叶叶绿素含量Fig. 1 Contrast of chlorophyll content under CK and light treatment with time
2.2 叶绿素荧光参数分析
图2a 表明两个处理下晒烟叶的实际光合效率(PhiPS2)都在下降,在实验的第2~7 d 内CK 的PhiPS2 一直高于光照处理。两处理的PhiPS2 都是在调制的前4 d 内有大幅下降,到第4 d 时CK 的 PhiPS2 比其第1 d 时低35%,而光照处理的PhiPS2降幅更大、比第1 d 时低了60%。图2b 中两个处理下烟叶的光合系统电子传递速率(ETR)同样有大幅下降,在第2~5 d 内CK 的ETR 高于光照处理。到第4 d 时CK 烟叶的ETR 比第1 d 时下降了46%,而光照处理的ETR 较第1 d 时下降了约75%。可见在晒烟调制1~4 d 内光照处理下烟叶的PhiPS2 和ETR 都比在CK 中下降得更快;因此同CK 相比,光处理下烟叶所接收的光能中将会有较多的光能因PhiPS2 和ETR 下降而无法参与光合反应,将有利于过剩光激发能的形成。

图2 不同天数内CK 和光照处理下烟叶的PhiPS2 和ETR Fig. 2 Contrast of PhiPS2 and ETR under CK and light treatment with time
两种处理下烟叶光化学淬灭值(qP)都在下降(图3a),在前4 天内最明显;CK 处理第4 d 的光化学淬灭值(qP)较其第1 d 的qP 值已降了31%,而此时光照处理下的qP 值较其第1 d 下降了52%。在2~7 d 内光照处理的qP 值一直显著低于CK、到第7 d 时比同期CK qP 值低38%。图3b 中,CK 处理下烟叶第5d的非光化学淬灭(qN)值升到峰值、较其第1 d 的qN值上升了71%,而光处理下烟叶第4 d 时qN 值达到峰值、比其第1 d 升高了110.5%,显著高于同期的CK处理。qP 和qN 的变化都表明调制中晒烟叶的光能利用率持续下降,而光照处理的降幅比CK 更大。在第4~7 d 内两个处理的qN 都转为下降,显示光合系统中以热的形式耗散掉的光能在减少;这或与同期内叶绿素含量明显下降、并导致烟叶获取的光能下降有关。

图3 不同天数内CK 和光照处理下烟叶的qP 和qN Fig. 3 Contrast of qP and qN under CK and light treatment with time
由图4 可见,在2~7 d 内CK 和光照处理下晒烟烟叶的Fv/Fm值均保持下降趋势,到第7 d 时CK 的Fv/Fm值比其第1 d 下降了49%,而光照处理的Fv/Fm值下降了70%。正常烟株上的烟叶Fv/Fm值维持在0.8~0.84 之间,遇到逆境胁迫越重、则Fv/Fm值降幅越大。在第2~7 d 中光照处理的Fv/Fm值一直显著低于CK 烟叶的Fv/Fm值,到第7 d 时比同期CK 的Fv/Fm值低43%,说明光照处理下烟叶光能转化率下降幅度比CK 大;由此推测光照下烟叶所承受的调制逆境胁迫比CK的更严重。

图4 不同天数内CK 和光照下烟叶的Fv/Fm Fig. 4 Contrast of Fv/Fm under CK and light treatment with time
2.3 H2O2、MDA 含量及抗氧化酶活性分析
两种处理下烟叶中H2O2含量都呈先增后减的变化趋势(图5);CK 中H2O2含量上升相对较缓,至第5 d 达峰值、约为第1 d 含量的4 倍;光照处理的H2O2含量上升相对较快,在第3 d 时就达峰值、约为其第1 d 含量的6 倍,极显著高于同期对照。在调制第2~5 d 内,光照处理的H2O2含量显著高于CK,表明光照处理下烟叶中的氧化胁迫大于CK。在第6~9 d 内CK 和光照处理的H2O2含量不再随晒烟调制时间的延长而上升,转而大幅下降;说明随着调制时间的延长烟叶细胞对调制逆境胁迫的反应能力在下降。

图5 不同天数内CK 和光照处理下烟叶的H2O2 含量Fig. 5 Contrast of H2O2 content under CK and light treatment with time
由图6 可见,CK 的MDA 含量持续上升至第7 d出现峰值12.23 μmol·mg-1,光照处理下的MDA 含量在调制3 d 后便已极显著高于遮光对照,并于第5 d达峰值19.06 μmol·mg-1,比CK 提前2 d 时间。两个处理下的MDA 含量峰值相差7.69 μmol·mg-1。此变化表明光照处理下晒烟叶细胞发生细胞膜脂过氧化的进度明显大于CK,细胞膜结构受损状况将比CK 更严重。
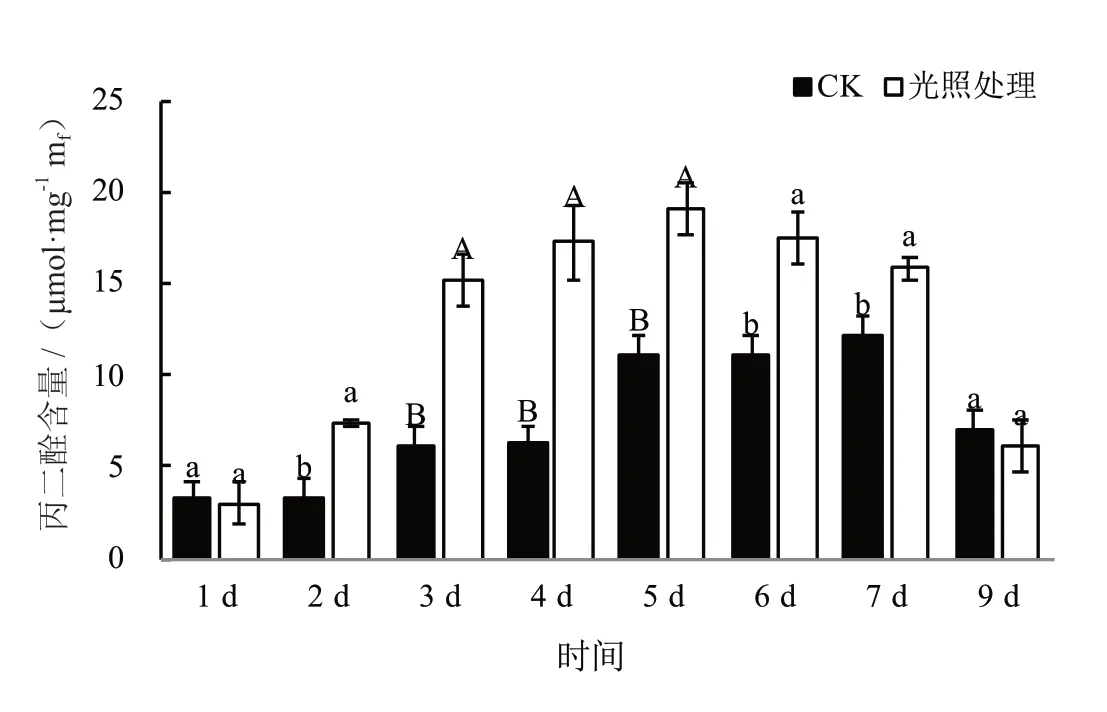
图6 不同天数内CK 和光照处理下MDA 含量Fig. 6 Contrast of MDA content under CK and light treatment with time
图7a 中CK 的POD 酶活性在第6 d 时升到峰值55.04 U·g-1,而光照处理下调制3 d 后烟叶POD 酶活性升到峰值59.57 U·g-1,它比CK 中的峰值提前3 d出现。图7b 中CAT 酶活性变化趋势与POD 相似,CK 的CAT 酶活性峰值晚于、且低于光照处理。CAT和POD 的酶活性的变化暗示:光照处理下烟叶的抗氧化反应比CK 中出现得更早、更剧烈。

图7 不同天数内CK 和光照处理下烟叶中POD 和CAT 酶活性 Fig. 7 Contrast of POD and CAT activity under CK and light treatment with time
在第6~9 d 中CK 和光照处理的CAT 和POD 的酶活性未随调制时间增长而继续升高,反而均出现大幅下降,到第9 d 时CAT 和POD 酶活性已低于它们第1 d 的水平,显示此时烟叶的抗氧化能力都已明显衰弱。
2.4 处理间OPLS-DA 分析
为分析遮光CK 处理和光照处理下烟叶中各种被测定指标间的相互关系及其对光、暗处理的响应状况,本文以各指标为变量对1~7 d 内光照处理和遮光对照烟叶样本群体进行了正交偏最小二乘判别分析(OPLS-DA),得到了具有显著性的OPLS-DA 模型,其对总方差的解释能力为R2X(cum)= 0.959、R2Y(cum)= 0.621,预测能力为Q2(cum)= 0.398。绘制变量载荷图(图8a)和变量的VIP(Variable Importance)图(图8b)。

图8 CK 和光照处理的OPLS-DA 模型中变量的载荷和VIP 分析Fig. 8 Loading plot and VIP plot on OPLS-DA model under CK and light treatment
变量载荷图(图8 a)中各指标变量所处的相限位置和彼此间邻近程度的反映了指标变量与两个处理间、以及指标彼此间的相关程度;其中 PhiPS2、ETR、qP、Fv/Fm和叶绿素(Chl)这5 个与光合作用密切相关的指标集中出现在与CK 相近的左上区域中、彼此间相互接近,说明它们彼此间相关性强;而H2O2、MDA、CAT 和POD 这4 种与细胞抗氧化有关的指标则较为集中地出现在图中右下部更靠近光照处理所在的位置、且与qN 较近;这反映这些抗氧化指标以及qN 之间的相关性较强;同时还表明在CK 处理下烟叶的总体光合能力大于光照处理下的烟叶,而光照处理下烟叶的抗氧化反应比CK 中的更强烈。
图8b 中PhiPS2、H2O2和MDA 这3 个 指 标 的VIP 值都在1.3 以上、明显高于其它指标,说明它们是对光照处理反应最为敏感、变化最突出的指标。因为以上指标是代表叶细胞光合作用和抗氧化能力的重要指标,由此可知在晒烟调制1~7 d 内,光照处理可以对烟叶细胞的光合作用和抗氧化能力产生重大影响。
3 讨论
前人研究表明光照能促进晒烟调制初期叶绿素含量的下降[14,23]。本研究也显示光照处理下叶绿素含量下降速度比CK 快。在吴疆的研究中叶绿素含量在调制1~4 d 内就已明显降低,而本文中叶绿素含量下降相对较缓。此差异很可能是实验条件或取样方法不同所致,如本研究中什邡烟区的生态环境、栽培品种、种植季节、栽培方法和采收、调制方法等多方面均与前者研究地万源烟区不同,而这些条件的差异都能影响晒烟叶内物质成分[2,5,7,10-12,14]。
叶绿素荧光参数是反映植物光合能力和植物所承受的胁迫程度的重要指标[24-28]。本研究中发现晒烟叶在调制变黄期1~7 d 内有明显的荧光信号,表明此时烟叶的叶绿素和光合系统仍然保有一定的工作能力。光照处理下晒烟叶的PhiPS2、ETR、qP 和Fv/Fm值都在调制过程中保持下降趋势,此现象与烟叶叶绿素荧光参数在干旱或高光强胁迫下的变化规律相符合[19-21]。CK 中PhiPS2 和ETR 等多个与烟叶光合能力呈正相关的荧光参数在调制过程中也在下降,但降幅低于光照处理。本研究中的Fv/Fm和其它多项荧光参数变化状况都表明光处理下烟叶所受胁迫程度更重。
H2O2和MDA 是植物逆境代谢的代表性物质,其含量高低与植物细胞所受胁迫程度密切相关[29-30]。调制初期CK 和光照处理中H2O2和MDA 浓度都有明显上升,这与宫长荣、陈秋芳的研究结论相符[11,31];CK 的H2O2和MDA 浓度都明显低于光照处理,这说明调制期间的光照处理使烟叶细胞中的过氧化物浓度提高了、使细胞膜结构受损更重。
光照处理下烟叶中快速大量形成的H2O2还激发了烟叶抗氧化酶活性的上升,CAT 和POD 酶活性峰值比CK 中出现得更早、更高。1~7 d内光、暗两种处理下烟叶中CAT 和POD 酶活性都是先升后降,这与陈秋芳、王宏辉研究结果相符[31-32]。结合前面分析的各项指标不难看出在5~7 d 内烟叶细胞在结构和功能上已受损或衰弱了;与此同时CAT 和POD 酶活性也快速下降,细胞最终将丧失抗氧能力。
本研究认为在晒烟调制初期,烟叶不仅有能力接收光合作用所需的光能,还接收到光照中的辐射热能和大量超过烟叶光合作用所能利用的“过剩光激发能”,它们可造成光胁迫、光破坏和叶内氧化胁迫。其作用机理如图9 示意:晒烟叶在变黄期受到失水、高温、强光等逆境因素的胁迫,导致叶细胞的叶绿素含量和光合能力不断地下降。因此,烟叶光合系统接收的一部分光能将无法利用、转而成为“过剩光激发能”,经光合系统转化后释放大量自由电子,它们与O2结合形成ROS 又使细胞内过氧化物(H2O2等)浓度上升。过氧化物的增加能促使细胞抗氧化反应加剧,使细胞内CAT 和POD 等抗氧化酶活性快速上升。然而,由于切断了烟叶的水分和养分物质补充,细胞抗氧化系统清除ROS 能力难以持久、继而衰弱。在另一方面,由于烟叶所处的调制逆境状况不断加重,使叶细胞中生成、积累更多的ROS。ROS 可使细胞膜脂发生过氧化产生MDA,造成细胞毒害、损害细胞结构和功能[33-34]。因此,光照处理下烟叶细胞的光合能力和抗氧化能力都因高浓度ROS 持续作用而明显下降,最后失去对光照等环境逆境因素的反应能力。

图9 光照对调制变黄初期的晒烟叶影响机制示意图Fig. 9 Mechanism schemes of light effect on sun-cured tobacco leaf in the early stage of yellowing period
郑昕、时向东等人的研究表明光照可提高烟叶香气物质含量、安全性、香气质和感官质量,但是过强的光照能使叶内物质转化不充分、烟叶品质下降[5,9]。本研究显示调制初期的光照可明显改变烟叶的光合能力和抗氧化反应,继而将影响到烟叶细胞中复杂的物质代谢乃至烟叶品质的形成。因此,如果进一步研究晒烟变黄期的光照条件与烟叶细胞中过剩光激发能的关系、细胞内过氧化物与烟叶品质的关系,将有助于合理控制晒烟调制过程的光照条件、优化晒烟品质。
4 结论
在调制变黄初期晒烟叶的叶绿素仍可接收环境光能并传递给光合系统。调制胁迫使晒烟叶的光能利用率明显下降,导致叶片所接收的部分光能无法被利用、转而成为过剩光激发能。这可促进H2O2和MDA 等物质的形成和积累,CAT 和POD 等抗氧化酶活性也因此升高。光照对调制变黄期烟叶的PhiPS2、MDA和H2O2含量影响最大。调制变黄期的光照可明显降低烟叶光合系统的工作能力、加剧烟叶中氧化胁迫状态,促进胞内物质转化。
猜你喜欢
中国机械工程(2022年8期)2022-05-09
湖南文理学院学报(自然科学版)(2021年3期)2021-10-23
儿童时代·幸福宝宝(2021年1期)2021-03-29
西南农业学报(2020年9期)2020-12-10
阅读(科学探秘)(2020年8期)2020-11-06
小资CHIC!ELEGANCE(2019年40期)2019-12-10
西南农业学报(2019年9期)2019-11-11
活力(2019年15期)2019-09-25
绿色科技(2019年2期)2019-05-21
中国果业信息(2019年1期)2019-01-05
