
嘉陵江中游银鮈食物组成及肠道异速生长模式初步研究
2020-10-09 09:11张富斌
广东农业科学 2020年8期
张富斌
(西华师范大学环境科学与工程学院/国家淡水渔业工程技术中心(武汉)西南分中心,四川 南充 637009)
【研究意义】消化道器官是鱼类从外界环境获取营养物质以满足自身能量需求的基础。肠道作为消化道器官的重要组成部分,其形态结构的变化往往受到食物组成、食物质量以及动物能量需求等因素的影响[1]。目前,国内外学者对消化道形态组织[2]、内分泌细胞[3]、微生物[4]等方面进行了相关研究,但关于消化道发育模式则鲜有报道[5]。
【前人研究进展】异速生长又被称作相对生长,是指生物在早期生长发育过程中,由于受到基因和外界环境因素的影响,身体各部分功能器官的生长速度具有不同步性[6]。生物体存在异速生长的现象自被报道后引起广大学者的密切关注[7]。越来越多研究发现,生物体在代谢速率[8]、胚胎生长和发育[9]及种群最大生长尺度等方面均存在异速生长现象。而国内对鱼类异速生长的研究大部分聚焦仔鱼在早期生存和环境适应中表现出来的器官异速生长[10-12]。面对复杂多变的外界环境,鱼类器官的异速生长能够提高其主动摄食和逃避敌害的能力,从而实现自我保护。
【本研究切入点】银鮈(Squalidus argentatus)又名亮壳、银色颌须鮈等,隶属鲤形目(Cypriniformes)鲤科(Cyprinidae)鮈亚科(Gobioninae)银鮈属(Squalidus)[13],广泛分布于我国各主要水系[14],产漂流性卵[15]。银鮈易捕获,在多数地区捕获渔获量大,而且肉质厚爽、鲜嫩味美,具有一定的经济价值。目前关于银鮈的研究大部分集中在繁殖生物学[16]、胚胎发育[17]、生长和死亡率[18]等方面,关于其食物组成及肠道发育模式的研究鲜有报道。
嘉陵江是长江流域面积最大的河流,但由于梯级航电枢纽建设(目前已建和在建水利枢纽共15级),渠化明显,鱼类栖息地破坏严重,原有自然流态河道多已成为库区,使得鱼类群落结构发生了较大变化[19],特别是对于产漂流性鱼卵的鱼类,库区的形成不利于其繁殖,致使其资源量必然随着航电枢纽的建设而减少。嘉陵江中游苍溪江段作为嘉陵江航电工程开发的缩影,在该江段附近共有3级航电枢纽,分别为亭子口航电枢纽、苍溪航电枢纽以及沙溪航电枢纽,鱼类生境破坏严重。
【拟解决的关键问题】本研究以采集于嘉陵江苍溪江段的银鮈作为研究对象,分析其食物组成、肠道形态结构,并探讨其异速生长模式与食物组成的关系,旨在探讨银鮈广泛分布的原因,同时为研究鱼类对环境适应性提供科学的参考依据。此外,由于人类影响,嘉陵江银鮈的生存必将受到威胁,本研究结果能在一定程度上为银鮈及其他鱼类的资源保护提供指导。
1 材料与方法
1.1 研究区域概况
嘉陵江由北向南流经陕西、甘肃、四川和重庆三省一市,干流全长1 119 km,流域面积达1.6×105km2,年径流量670亿m3[13]。嘉陵江上游包括源头至广元昭化江段,中游为昭化至重庆合川江段,合川江段至河口为下游江段[20]。本研究样品采自嘉陵江中游苍溪江段(图1),采样范围为31°38′51″~31°48″24″N,105°52′30″~105°55′47″E。苍溪江段受人类活动影响严重,在约35 km的河道上建有3级梯级航电枢纽,相应江段几乎全部成为库区,对鱼类栖息地生境影响严重。
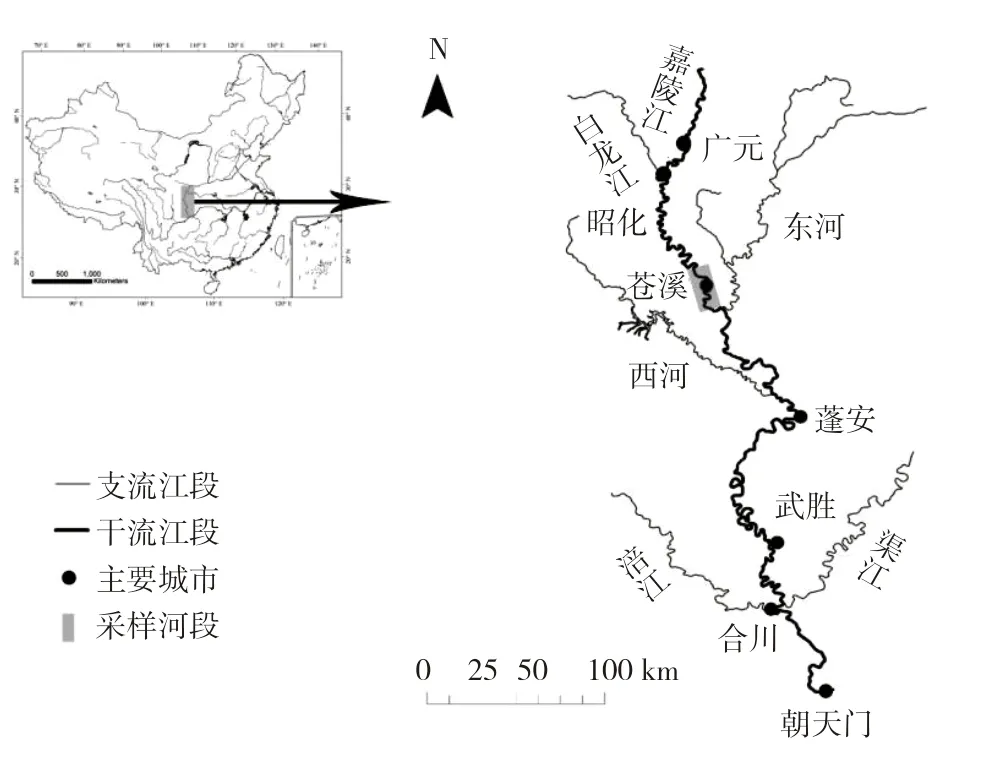
图1 嘉陵江中游苍溪江段银鮈采集地Fig.1 Sampling sites of Squalidus argentatus in midstreamof the Jialing River
1.2 样品采集
于2016年6月和8月,利用定置刺网(8 m×1.2 m×2 cm)在嘉陵江中游苍溪江段共采集银鮈46尾。为避免鱼类食物被消化,每次放置网具的时间不超过4 h。鱼类一旦被捕获,立即带回野外实验室进行处理。
1.3 样品处理
在野外实验室,首先测量鱼体的全长、体长、体重。随后立即对鱼进行生物学解剖,记录鱼肠充塞度(0~5,0表示空肠,1表示肠道饱满度为20%,5表示肠道饱满度为100%,依此类推),并将肠充塞度≥3的肠道内容物取出保存于10%的福尔马林溶液中,以待后续进行鱼类食物组成观察。然后,立即将肠道洗净,利用游标卡尺和电子天平分别测量鱼类前肠、中肠和后肠的长度及重量,鱼体整体肠道长度与重量分别等于前肠、中肠及后肠三区的长度与重量之和。
1.4 食性观察
为了避免因食物消化给食物鉴定带来误差,仅选用肠充塞度≥3的样本用于食性分析。在室内鉴定时,先用0.5 mm网目筛网将肠道内容物过滤。过滤液全部收集用作藻类及浮游动物类群鉴定,而留在筛网上的大型食物类群(如水生昆虫、软体动物等)则主要通过肉眼或解剖镜(Nikon SM-6)进行鉴定计数,并在电子天平上直接称量食物重量。藻类及浮游动物的鉴定通过显微镜(Leica DM500)进行鉴定计数,小型食物的种类通过体积转换法[21]进行计算,具体鉴定方法参照文献[22-23]。
食物的鉴定主要参照《中国淡水藻类系统、分类及生态》[24]、《Aquatic Insects of China Useful for Monitoring Water Quality》[25]、《淡 水微型生物与底栖动物图谱》[26]以及《水生生物学》[27]等文献资料。
1.5 数据分析
1.5.1 肠道指数分析 肠道作为银鮈(为无胃鱼)的消化器官,对银鮈生长发育起着至关重要的作用。本研究对银鮈肠道发育通过比肠长(Relative gut length,RGL)、Zihler指数(Zihler index,ZI)、比肠重(Relative gut mass,RGM)以及单位长度肠道质量(W/L)4个指数来描述和衡量。其中,RGL和ZI侧重于鱼体肠道长度的描述,而RGM和W/L更偏向于肠道质量和发达程度的衡量。各指数计算方法参照文献[5]。
银鮈前肠、中肠及后肠间各肠道指数的差异通过单因素方差分析(One-Way ANOVA)进行检验,采用SPSS 22.0软件计算分析。
1.5.2 异速生长模式分析 银鮈异速生长模型采用幂函数方程(Y=aXb)计算。本研究中以银鮈全长为自变量,其整体肠道或肠道各分区长度为因变量,a为Y轴截距,b为异速生长指数[28]。为了优化分析,对数据统一进行Log转换,其后通过R软件(Segmented分析包)[29]进行统计分析。通过t检验,当b>1时,表明肠道为快速生长;当b<1时,为慢速生长;若b与1无显著性差异,则可认为是等速生长。异速生长曲线图采用Origin 9.0绘制。
1.5.3 食性分析 银鮈食物组成采用出现率、数量百分比、质量百分比、相对重要指数、相对重要指数百分比等5个指数衡量,各指数计算公式如下:
出现率(%)=(含有某种食物的肠道数量/解剖后有食物的肠道数量)×100
数量百分比(%)=(某种食物出现的个数/所有食物的总个数)×100
质量百分比(%)=(某种食物的质量/所有食物的总质量)×100
相对重要指数=出现率百分比×(数量百分比+重量百分比)
相对重要指数百分比(%)=(相对重要指数/相对重要指数的总和)×100
2 结果与分析
2.1 样品采集情况
本研究共采集银鮈46尾,所有肠充塞度均≥3,全部用于食性分析。46尾银鮈全长介于96.0~152.0 mm之间,平均130.5 mm;体长介于79.0~123.0 mm之间,平均106.7 mm;体重介于8.2~36.7 g之间,平均22.8 g。
2.2 银鮈肠道形态特征
解剖发现银鮈属于无胃鱼类,肠道盘曲方式简单,呈“S”型。前肠、中肠和后肠的比肠长(RGL)平均值分别为0.2219、0.1811、0.3705,比肠重(RGM)平均值分别为0.0070、0.0056、0.0058,Zihler指数(ZI)平均值分别为0.8412、0.6862、1.4046,单位肠道质量(W/L)平均值分别为0.0067、0.0051、0.0034(表1)。单因素方差分析表明,银鮈前肠、中肠与后肠三区的RGL、RGM、ZI、W/L之间均存在显著差异。LSD多重比较结果显示,银鮈的RGL和ZI具有相似结果,其中中肠最小、前肠次之、后肠最大;RGM比较结果为前肠最低、中肠其次、后肠最高,但后肠与中肠差异不显著;W/L两两比较结果显示后肠最低、中肠次之、前肠最高,上述结果可能与各肠道分区的功能有关。
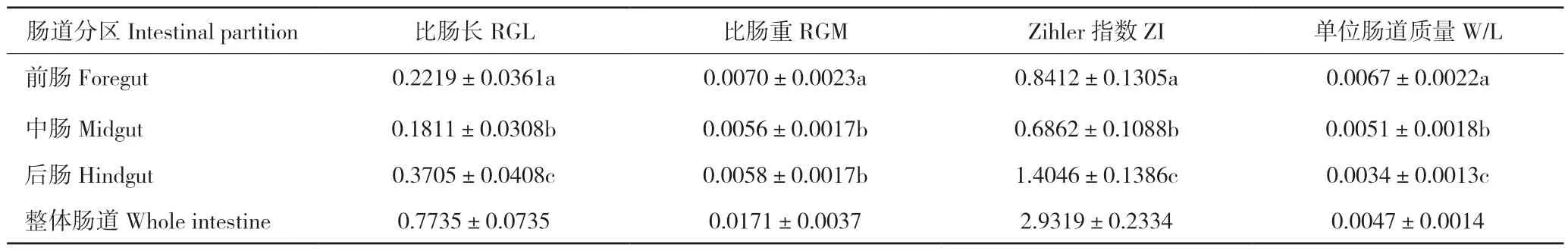
表1 银鮈肠道指数Table 1 The intestinal index of Squalidus argentatus
2.2 银鮈肠道异速生长模式分析
对银鮈整体肠道以及肠道各分区异速生长整体分析表明,整体肠道(b=1.5388>1,P<0.05)、前肠(b=1.8288>1,P<0.05)以及中肠(b=1.8496>1,P<0.05)均为快速生长;而后肠的异速生长指数(b=1.2418)与1比较差异不显著,故可认为是等速生长(表2)。
t检验结果表明,整体肠道和前肠出现了生长拐点(图2)。其中,整体肠道出现生长拐点对应的鱼体全长为131.00 mm,对应鱼体肠道长度为83.45 mm(表3)。整体肠道在拐点前,异速生长指数为1.7491(P<0.05),为快速生长;拐点后,异速生长指数为1.4305(P>0.05),为等速生长(图2A)。而前肠出现生长拐点时对应的鱼体全长为129.39 mm,对应的前肠长度为23.28 mm(表3)。前肠在拐点前,异速生长指数为2.6565(P<0.05),为快速生长;拐点后,异速生长指数为0.5463(P>0.05),表现出等速生长(图 2B)。而中肠和后肠未出现生长拐点(图2C和图2D)。
2.3 银鮈食物组成分析
由于前肠为主要的摄食消化场所,因此,本研究以前肠拐点为依据,将银鮈划分为拐点前和拐点后进行食性观察并分别进行统计。其中拐点前(全长<129.39 mm)共包括24尾,拐点后(全长≥129.39 mm)共包括22尾。
拐点前,银鮈食性主要包括藻类、水生昆虫及有机碎屑,其中藻类包括硅藻、蓝藻、绿藻以及黄藻。从出现率来看,拐点前有机碎屑出现率最高(87.5%),其次是舟形藻和平板藻,出现率均为62.5%;从数量百分比来看,拐点前羽纹藻最高(24.86%),其次是舟形藻(24.43%)和卵形藻(15.28%);从质量百分比来看,拐点前碎屑最高(76.84%),其次是摇蚊幼虫(21.16%)和舟形藻(0.31%);从相对重要指数百分比来看,拐点前有机碎屑最高(57.41%),其次是舟形藻(11.52%)和羽纹藻(10.54%)(表4)。总体而言,拐点前,银鮈的主要食物包括有机碎屑、摇蚊幼虫以及硅藻等。
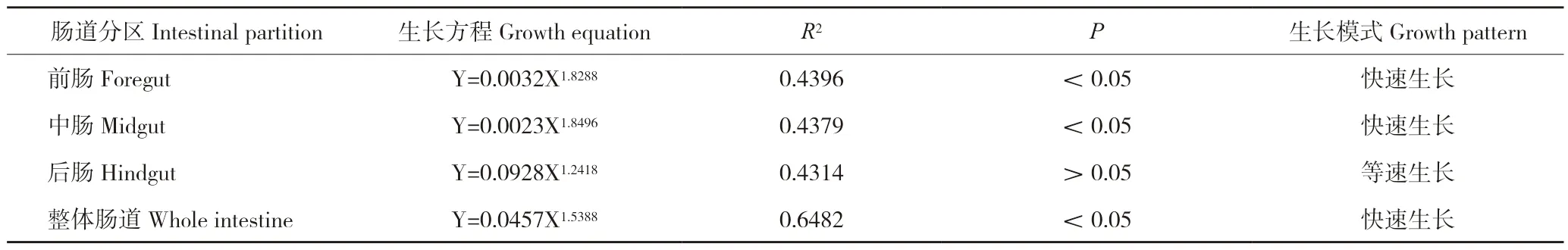
表2 银鮈肠道生长方程Table 2 The growth equation of Squalidus argentatus
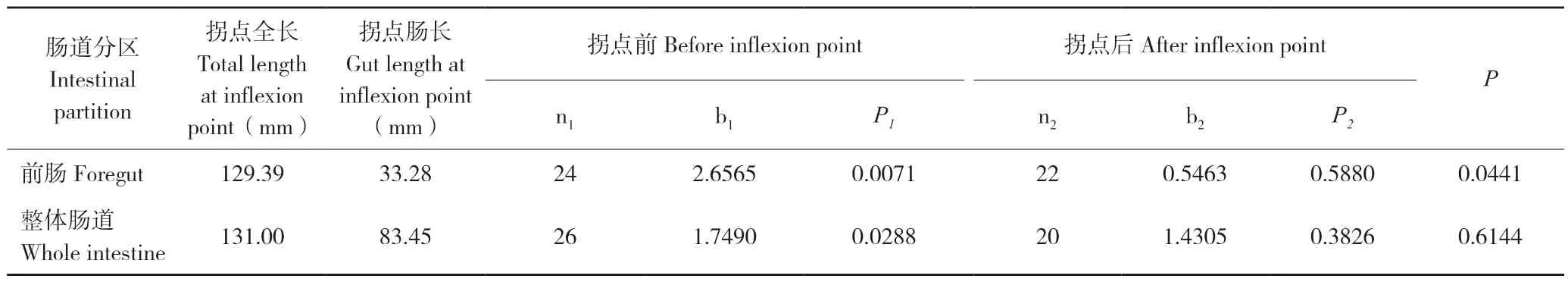
表3 银鮈肠道异速生长模式Table 3 The allometric growth pattern of Squalidus argentatus
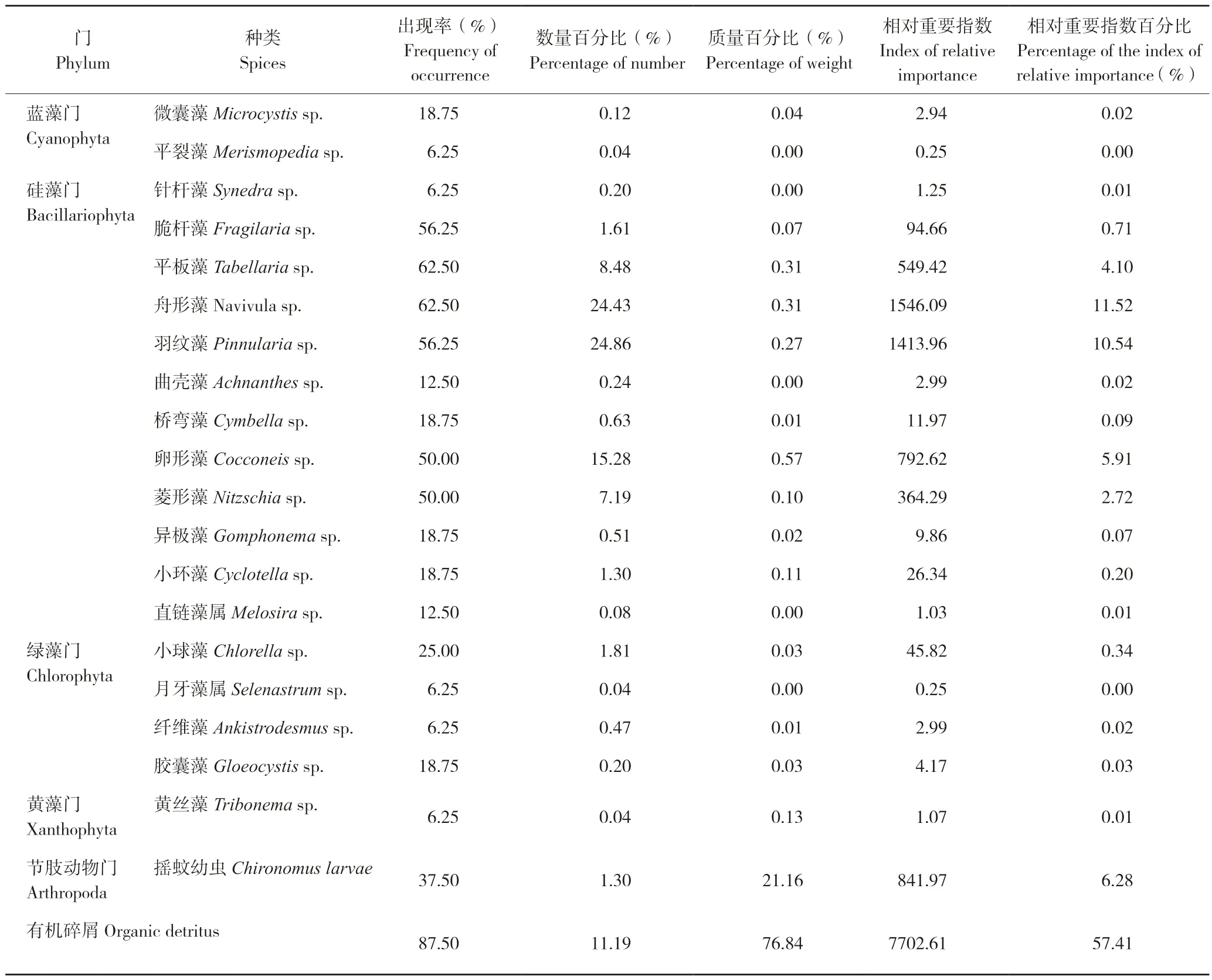
表4 拐点前银鮈食物种类组成Table 4 Food composition of Squalidus argentatus before the inflection point
拐点后,银鮈的食性包括藻类、原生动物、软体动物、节肢动物以及有机碎屑,其中藻类包括硅藻、蓝藻、绿藻、甲藻以及黄藻。从出现率来看,拐点后有机碎屑最高(64.29%),其次是菱形藻、卵形藻、脆杆藻、小环藻(均为42.86%);从数量百分比来看,拐点后羽纹藻最高(40.30%),其次是舟形藻(29.15%)和有机碎屑(12.60%);从质量百分比来看,拐点后有机碎屑最高(90.41%),其次是淡水壳菜(6.64%)以及摇蚊幼虫和变形虫(均为0.51%);从相对重要指数百分比来看,拐点后有机碎屑最高(80.30%),其次是羽纹藻(10.57%)和舟形藻(2.55%)(表5)。总体而言,拐点后,有机碎屑、硅藻以及淡水壳菜是银鮈的主要食物组成,相较于拐点前,拐点后银鮈摄入的肉食性饵料有所减少。
综上所述,嘉陵江中游苍溪江段的银鮈食物来源广泛,属于杂食性鱼类。通过比较银鮈拐点前和拐点后的食性可以发现,拐点后银鮈摄入的食物种类和类群更加丰富,包括软体动物以及原生动物,有机碎屑对拐点后的银鮈食性贡献增加,达80.30%。
3 讨论
3.1 银鮈肠道特征
鱼类消化道的形态结构与其食性具有很大相关性。比肠长(RGL)一般可视为形态结构的指标[30],RGL值较低的多为典型的肉食性鱼类,RGL值较高的多为典型的植食性鱼类,RGL值介于二者之间的通常为杂食性鱼类。杂食性鱼类的RGL值往往与饵料中植物与动物的比例有关。例如,研究发现哲罗鱼的RGL为0.47,肠道一般没有折叠盘旋,属于典型的肉食性鱼类[31];而华鲮的RGL为11.10,肠道高度盘曲,属于典型的植食性鱼类[32]。本研究结果表明,银鮈整体肠道的RGL为0.7735,而食性鉴定结果显示,其食物主要由有机碎屑、硅藻、摇蚊幼虫、淡水壳菜组成,食物种类繁多,属于杂食性鱼类。
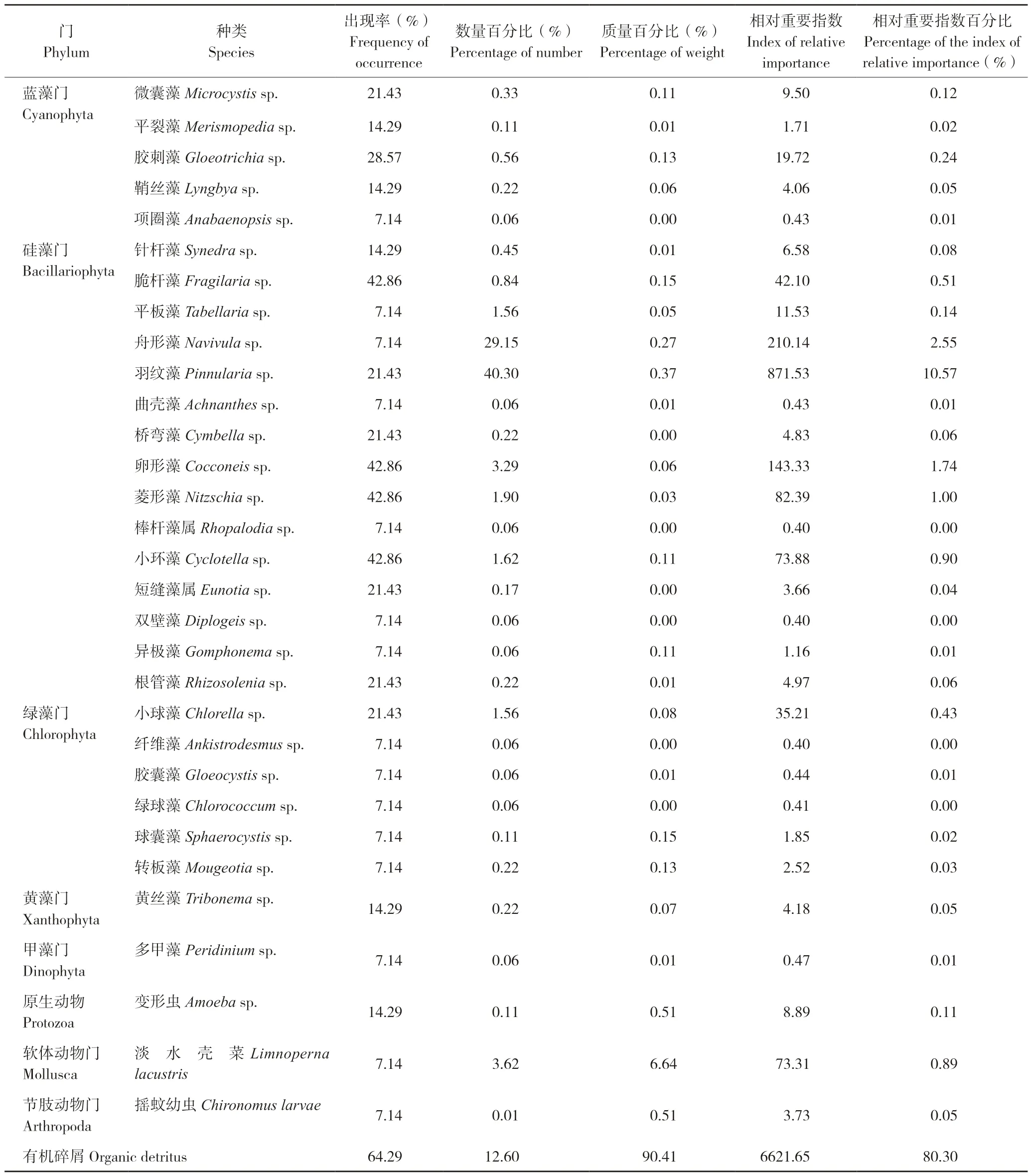
表5 拐点后银鮈食物种类组成Table 5 Food composition of Squalidus argentatus after the inflection point
3.2 银鮈肠道形态和异速生长特征
银鮈属无胃鲤科鱼类,其进化程度不如高等脊椎动物。因此,肠道并没有严格意义上的分区。本研究采用大部分学者意见,将其肠道划分成前、中、后三部分[33-34]。从形态学指数来看,银鮈前肠最重且最发达,中肠和后肠差异不显著。李茜[35]研究表明,异育银鲫肠道前部的横截面积较大且管径较粗,中肠与后肠部分横截面积变小,以至于很难区分二者的分界线。本研究结果发现,银鮈中肠和后肠也存在相似特征,这与李茜[35]的研究结果基本一致。
在鱼类中,重要器官优先发育,次要器官延缓发育,这是生物体适应环境的表现[36]。鱼类异速生长的发育模式提高了仔鱼逃避天敌和觅食的能力,从而帮助鱼类更好地适应复杂生境,保证其存活率[11]。本研究结果显示,银鮈的肠道存在异速生长模式,其前肠和整体肠道在幼鱼阶段快速生长,到后期变为等速生长。无胃鱼类,前肠是其最重要的食物消化场所,也是其摄取营养物质的重要场所。本研究发现,银鮈前肠在幼鱼时为快速生长,这一策略能够帮助银鮈幼鱼最大限度地快速消化食物,获取自身生长发育所需的营养,促进其在幼鱼时期较快速生长发育,以尽快适应复杂环境,同时也可以提高存活率。总体而言,银鮈肠道的异速生长模式可能与其对生活环境的适应息息相关,这可能也是其能广泛分布的原因之一。本研究仅对银鮈肠道的生长模式进行了探讨,后续有待开展更多相关研究以期更好地了解其生长模式与生境适应关系。
3.3 银鮈异速生长和食物组成变化间关系及生物学意义
研究表明,肠道的异速生长可能与其功能分化以及能量摄入相适应[5]。本研究中,银鮈前肠的快速生长可以有效提高银鮈对饵料的消化能力,促进摄食频率的提高,从而满足鱼体快速生长所需的大量能量。而银鮈中肠粘膜层上的细胞表面具有密集的微绒毛,且粘膜压缩形成许多皱褶。这种组织结构的分化使饵料在中肠的停留时间延长,同时增大了饵料与消化液的接触面积,通过中肠的快速生长,饵料被吸收更加充分,从而提高了能量的吸收效率。银鮈后肠发达程度最低,管径最小,长度最长,维持等速生长使食物残渣排出体外。在银鮈肠道发育过程中,拐点后食物出现了变形虫、淡水壳菜,且藻类种类增多。据此我们推测,由于银鮈在拐点后已为成体,其消化能力和对环境的适应能力均有所增强,使得银鮈前肠生长速度变慢,此后银鮈通过改变食物类型,一定程度上可以减少了种内食物竞争;此外,在拐点前,银鮈摄入更多的肉质饵料(摇蚊幼虫,质量百分比21.16%),小个体鱼类摄入更多优质饵料能够快速生长,从而更快适应环境,这一策略有助于该物种种群的壮大,这与异齿裂腹鱼(Schizothorax oconnori)[21]和拉萨裸裂尻鱼(Schizopygopsis younghusbandi younghusbandi)[37]的生长策略相同。上述现象可能是银鮈肠道自身发育及长期与环境适应的结果。
4 结论
嘉陵江中游银鮈前肠、中肠、后肠在其长度、质量、发达程度等方面均存在显著差异。其中,银鮈前肠和整体肠道存在异速生长,中肠保持快速生长,后肠保持等速生长。其前肠与整体肠道表现出异速生长模式,这可能与肠道各分区功能密切相关,这一结果应是其对环境长期适应的结果。食性分析结果表明,嘉陵江中游银鮈食物来源广泛,属于杂食性鱼类,主要摄食有机碎屑、藻类、软体动物、水生昆虫。在前肠拐点前,银鮈(全长<129.39 mm)摄入更多肉食性饵料(质量百分比21.16%),而拐点后(全长≥129.39 mm),肉食性饵料比例降低,这可能是银鮈肠道发育及长期与环境适应的结果。
猜你喜欢
儿童时代·幸福宝宝(2020年9期)2020-09-08
当代水产(2020年4期)2020-06-16
汽车与新动力(2019年5期)2019-11-07
汽车观察(2019年2期)2019-03-15
红岩春秋(2017年6期)2017-07-03
探索科学(2017年5期)2017-06-21
股市动态分析(2015年29期)2015-09-10
海外星云 (2013年3期)2013-02-20
雕塑(1997年3期)1997-06-23
雕塑(1997年3期)1997-06-23
