
滇东南典型常绿阔叶林土壤酶活性的海拔梯度特征
2020-11-27 02:58吕晶花任玉连杨罗平王东旭
林业科学研究 2020年6期
李 聪,吕晶花,陆 梅*,任玉连,杜 凡,陶 海,杨罗平,王东旭
(1. 西南林业大学生态与环境学院,云南 昆明 650224;2. 西南林业大学林学院,云南 昆明 650224;3. 贵州大学生命科学学院,贵州 贵阳 550025)
土壤酶是一类主要来源于土壤中动植物体、微生物体的细胞分泌物释放的生物催化剂[1],它不仅在土壤养分循环及能量代谢方面扮演重要的角色,而且是评价土壤肥力、土壤质量和健康状况的一个重要生物学指标[2]。土壤酶活性的时空变化受植被类型[3]、土壤生物[4]、土壤水热条件[5]及土壤养分[6]等生物与非生物因素的共同调控。研究表明,海拔变化会引起区域小气候、植被群落和土壤理化性质等环境因子的梯度效应,进而显著影响土壤酶活性[7-8]。任玉连等[9]研究发现,随着海拔梯度变化,植物群落组成、凋落物性质、微气候及土壤理化性质等环境因子均发生相应改变。姚兰等[10]认为,森林植被类型能够直接或间接地改变土壤活性有机碳/氮组分及含量,从而显著影响土壤酶活性。因此,土壤酶活性能够对海拔引起的气候、植被类型和土壤理化性质等环境因子变化做出响应。然而,调控不同区域土壤酶活性的主导因子可能存在一定差异性,使土壤酶活性对微气候、植被与土壤沿海拔梯度变化的响应存在地区差异。因此,开展不同地理区域森林土壤酶活性的海拔梯度变化及影响机制研究,不仅有助于理解环境因子变化对森林土壤酶活性及土壤生态学过程的作用机制,而且有助于探明森林土壤对全球变化响应的大小、方向与过程。
云南文山国家级自然保护区地处典型南亚热带季风气候区,森林植被群落结构随海拔升高变化明显,且不同海拔植被带的水热条件、群落多样性及土壤理化性质存在显著的空间异质性,是研究亚热带不同海拔梯度森林植被与土壤相互作用的理想场地。因此,本研究选择文山自然保护区3个不同海拔的典型植被类型(亚热带季风常绿阔叶林、半湿润常绿阔叶林和湿性常绿阔叶林)为研究对象,研究土壤酶活性随海拔变化的规律及其影响因素,进而阐明土壤酶活性对海拔高度变化的响应特征,为理解该保护区不同海拔“水热-植被-土壤”之间相互作用对土壤酶活性影响的生态学过程及机制提供理论与数据支撑。
1 研究区概况
文山国家级自然保护区(23°21′00″~23°24′00″ N,104°41′00″~104°41′00″ E)地处滇东南低纬度高原,属中山山原地貌。该区位于南亚热带季风湿润气候区,具气温高、雨量多及湿度大等特点。保护区地势起伏平缓,相对高差683 m。年均温差沿海拔升高由21.1℃降至12.4℃,年均降水量由1 050 mm增至1 630 mm,沿海拔自下而上形成了亚热带季风常绿阔叶林、半湿润常绿阔叶林、湿性常绿阔叶林、苔藓常绿阔叶林及山顶苔藓矮林的天然植被垂直分布景观和森林生态系统[11]。
2 研究方法
2.1 样地设置
选取文山自然保护区内3个不同海拔的典型植被带,即亚热带季风常绿阔叶林(ME)、半湿润常绿阔叶林(SE)和湿性常绿阔叶林(HE)。每个植被带随机布设3个重复样地,样地大小30 m ×30 m(间距约 > 200 m)。样地基本情况详见表1。

表 1 样地基本情况Table 1 The basic situation of the sample plots
2018 年10月,按照上述每个植被类型的3个重复样地,在每个样地中采用对角线法选取3个1 m × 1 m取样点,除去地表覆盖的凋落物,挖掘宽1 m、深0.8 m的土壤剖面,沿土层自上而下分层采样(0~10、10~20、20~30、30~40、40~50 cm),剔除石砾、根系等杂物后,用四分法取适量土壤装入无菌自封袋中,低温带回实验室。将各土壤样品进行如下处理:约200 g用于土壤含水量测定;约1 kg冷藏于冰箱中,于2周内测定土壤酶活性;约1 kg置于室内自然风干、磨细过筛,用于测定土壤基本性质。各植被带内3个重复样方测定的均值作为各植被类型土壤酶活性和理化性质指标。
2.2 土壤理化指标测定
气温与降水量数据来源于《云南文山自然保护区综合科学考察报告》[11]。土壤理化指标测定参照中华人民共和国土壤检测方法林业行业系列标准[12]。土壤含水量采用烘干法;土壤容重采用环刀法;土壤pH值采用(水土比 1:2.5)电位法;总有机碳采用重铬酸钾氧化-外加热法;全氮采用半微量凯氏定氮法;土壤全磷采用酸溶-钼锑抗比色法;速效磷采用氟化铵-盐酸浸提比色法;速效氮采用碱解-扩散法;全钾采用碱溶-火焰光度计法;速效钾采用乙酸铵浸提-火焰光度计法。每个土壤样品至少重复测定3次,取平均值。土壤温度采用地温计测定。
2.3 土壤酶活性测定
土壤酶活性测定方法参照《土壤酶及其研究方法》[13]。过氧化氢酶(Cat)采用KMnO4滴定法,以1 g·h-1土壤消耗0.1 mol·L-1KMnO4的毫升数表示H2O2酶活性;脲酶(Ure)采用苯酚-次氯酸钠比色法,以培养1 d后1 g土壤中生成NH3-N 的毫克数表示脲酶活性;蔗糖酶(Suc)采用3,5-二硝基水杨酸比色法,以培养1 d后1 g土壤中生成葡萄糖的毫克数表示蔗糖酶活性;酸性磷酸酶(Acp)采用磷酸苯二钠比色法,以1 g·h-1土壤中释放的酚毫克数表示磷酸酶活性。每个土壤样品至少测定3次,取平均值。
2.4 植物多样性
植被多样性指数采用如下测度[14],其计算公式如下:
Margalef丰富度指数(R):R= (S- 1)/lnN
Shannon-Wiener多样性指数(H):
H= -∑(PilnPi)
Simpson多样性指数(D):D= 1 - ∑(Pi/N)2
Pielou均匀度指(J):J=H/lnS
式中:Pi=Ni/N,即某个物种的相对多度;Ni为种i的株数;S为种i所在样方的所有物种的总株数[15]。
2.5 数据处理
采用Microsoft Excel 2013对实验数据预处理、数据统计和作图;采用单因素方差分析(Oneway Anova)对各环境因子(水热条件和植物多样性、土壤理化性质)进行多重比较和差异性分析。采用前向选择法对各环境因子进行约束性分析,然后使用蒙特卡洛检验对环境因子进行重要性排序;根据重要性排序的结果,筛选出对酶活性具有显著影响的环境因子,采用Canoco5.0中的T-value双序图进行单一因子检验,明确其与各酶活性的关系,若某个土壤酶活性指标的箭头连线落入或者穿过黑色圈表示呈显著正相关,落入或穿过白色圈为显著负相关[16]。
3 结果与分析
3.1 不同海拔植被带水热条件及物种多样性特征
图1表明:不同植被带年均气温、年均降水量及土壤温度沿海拔梯度的变化差异显著(P<0.05),其中,年均气温和土壤温度随海拔升高而降低,即亚热带季风常绿阔叶林 > 半湿润常绿阔叶林 > 湿性常绿阔叶林,年均降水量随海拔升高而增加,即亚热带季风常绿阔叶林 < 半湿润常绿阔叶林 < 湿性常绿阔叶林。
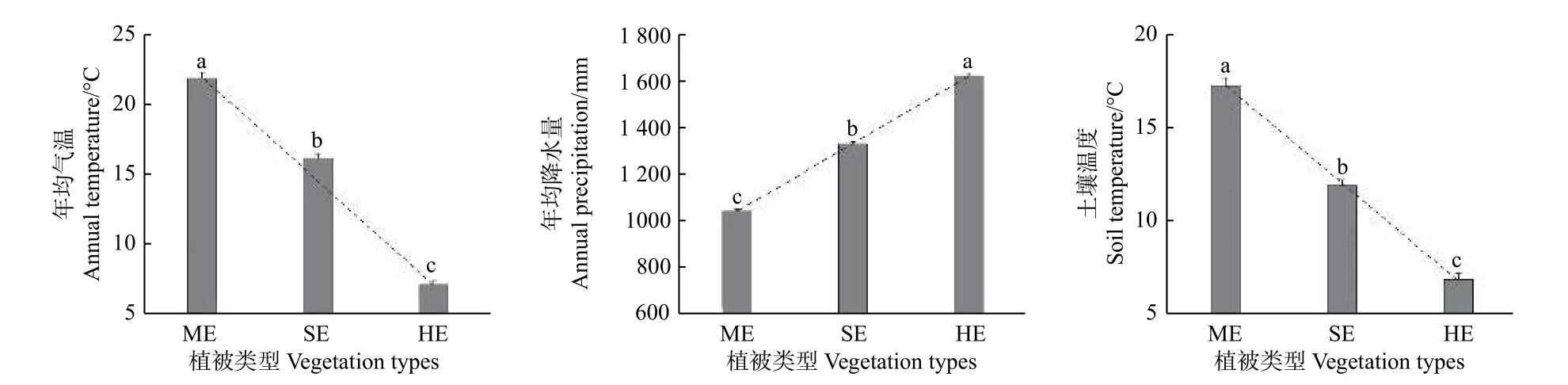
图 1 不同植被带水热条件的海拔变化特征Fig. 1 The hydrothermal conditions of different vegetation zones at different altitudes
表2表明:不同植被带Margalef(R)、Shannon-Wiener(H)、Simpson(D)和Pielou(J)指数沿海拔梯度的变化趋势不同,R、H和J指数均随海拔升高而减少,其中,3个植被带的R、H指数差异显著(P< 0.05),亚热带季风常绿阔叶林和半湿润常绿阔叶林的J指数差异不显著(P> 0.05);D指数沿海拔升高而增加,且3个植被带间差异显著(P< 0.05)。
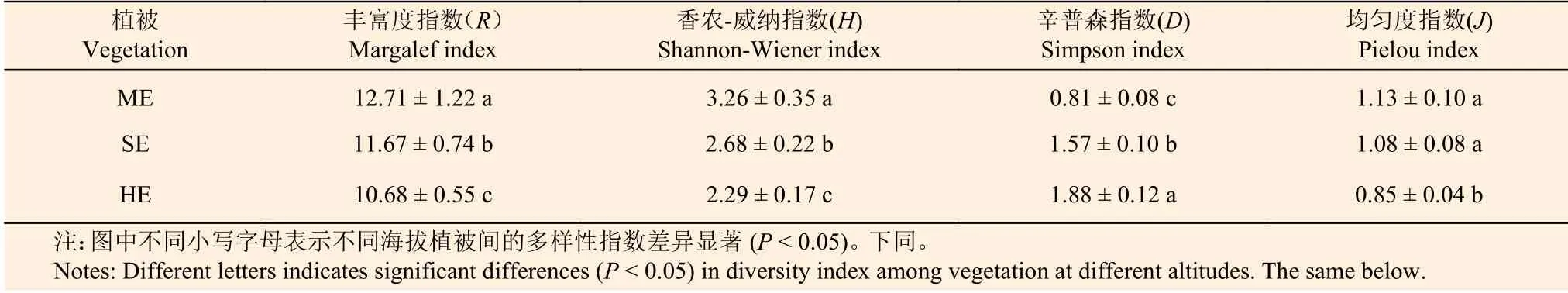
表 2 不同海拔样地植物多样性指数Table 2 The diversity index of plant communities at different altitudes
3.2 不同海拔植被带土壤理化性质特征
3.2.1 土壤理化性质沿植被带变化 由表3可知:各海拔植被带土壤理化性质沿海拔梯度呈不同变化规律,且差异显著(P< 0.05);土壤容重和pH值分别随海拔升高而降低;土壤含水量、总有机碳、全氮、全钾、速效氮和速效钾随海拔升高而增加,土壤全磷和速效磷随海拔升高先降低后增加。湿性常绿阔叶林土壤含水量、总有机碳、全氮、全磷、全钾、速效氮、速效磷和速效钾的含量分别是亚热带季风常绿林的1.53、2.61、2.70、1.66、2.26、1.77、2.18、1.71倍。
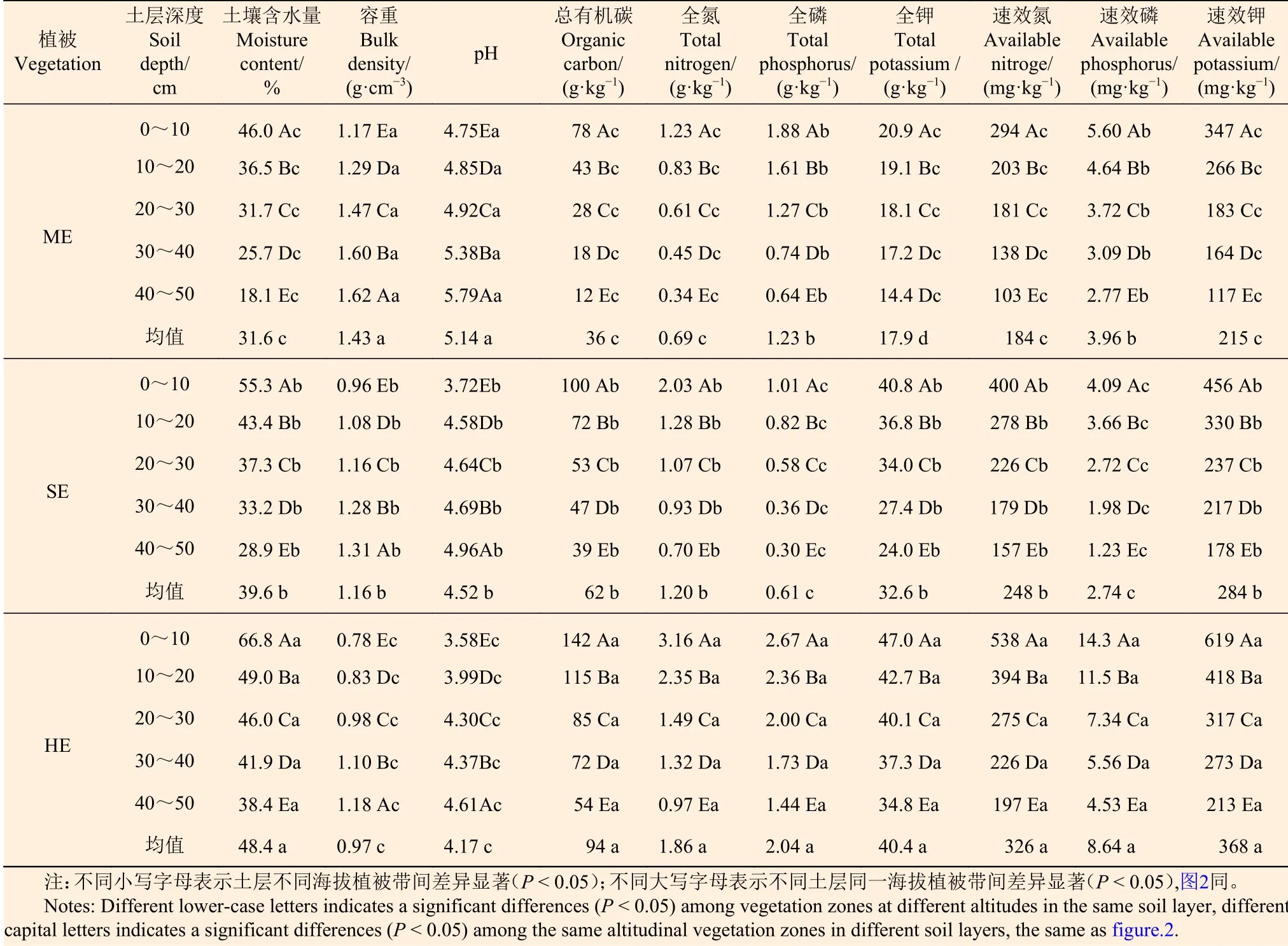
表 3 不同海拔植被带土壤理化性质特征 Table 3 Soil physicochemical properties of vegetation zones at different altitudes
3.2.2 土壤理化性质的垂直变化 各植被带土壤理化性质沿土层深度呈现不同变化规律,且差异显著(P< 0.05)(表3)。土壤含水量、总有机碳、全氮、全磷、全钾、速效氮、速效磷和速效钾的含量均表现为随土层加深而减小,0~10 cm最大,40~50 cm最小;与0~10 cm相比,40~50 cm的上述各指标,亚热带季风常绿阔叶林分别减小27.9 %、66 g·kg-1、0.89 g·kg-1、1.24 g·kg-1、6.5 g·kg-1、191 mg·kg-1、2.83 mg·kg-1、230 mg·kg-1,半湿润常绿阔叶林分别减小26.4 %、61 g·kg-1、1.33 g·kg-1、0.71 g·kg-1、16.8 g·kg-1、243 mg·kg-1、2.86 mg·kg-1、278 mg·kg-1,湿性常绿阔叶林分别减小28.4 %、88 g·kg-1、2.19 g·kg-1、1.23 g·kg-1、12.2 g·kg-1、341 mg·kg-1、9.77 mg·kg-1、406 mg·kg-1。土壤容重和pH值均表现为随土层加深而增加,40~50 cm最大,0~10 cm最小。
3.3 不同海拔植被带土壤酶活性特征
3.3.1 不同植被带土壤酶活性的比较 土壤脲酶、过氧化氢酶和蔗糖酶活性在不同海拔植被带间差异显著(P< 0.05),且随海拔上升呈增大趋势(图2)。脲酶、过氧化氢酶和蔗糖酶活性均为湿性常绿阔叶林 > 半湿润常绿阔叶林 > 亚热带季风常绿阔叶林。
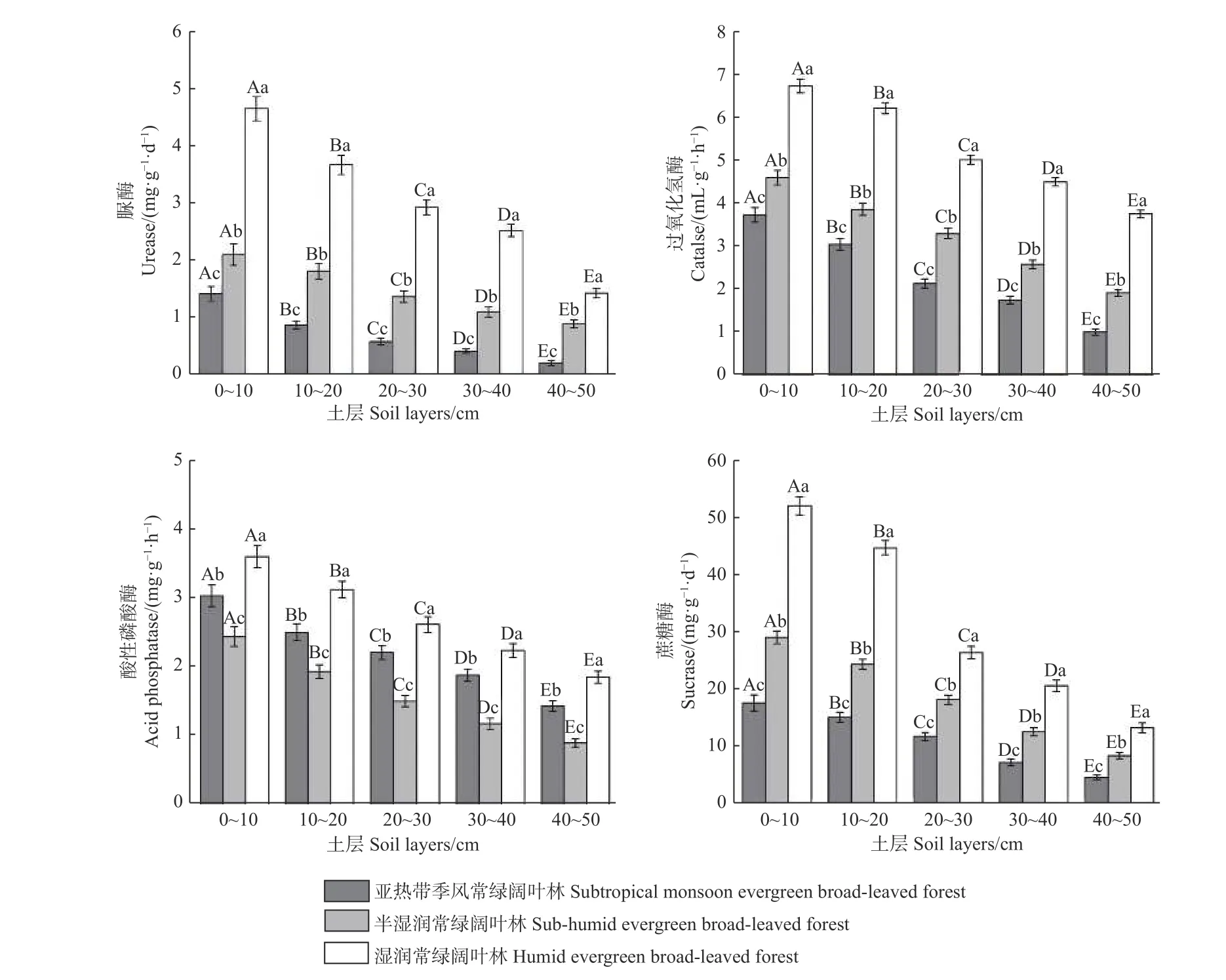
图 2 不同海拔植被带土壤酶活性变化Fig. 2 Soil enzyme activities of different vegetation zones at different altitudes
不同海拔植被带酸性磷酸酶活性差异显著(P<0.05),且随海拔上升呈先减小后增大趋势(图2),其活性表现为湿性常绿阔叶林 > 亚热带季风常绿阔叶林 > 半湿润常绿阔叶林。
3.3.2 土壤酶活性随土层的变化 土壤酶活性沿剖面的变化均差异显著(P< 0.05),且随土层加深而降低,表现为0~10 cm土层最高,40~50 cm土层最低。土壤脲酶、过氧化氢酶、酸性磷酸酶、蔗糖酶其活性呈现:0~10>10~20>20~30>30~40>40~50 cm。同土层的酶活性沿海拔上升呈显著不同的变化(P< 0.05),土壤过氧化氢酶、脲酶和蔗糖酶活性沿海拔升高呈增加趋势,酸性磷酸酶活性呈先降低后增加趋势。同一植被带下,土壤酶活性均随土层加深逐渐降低(图2)。
3.4 土壤酶与环境因子的相关关系
3.4.1 环境因子对土壤酶活性的冗余(RDA)排序 水热条件、植物多样性、土壤理化性质等环境因子对4种土壤酶活性在第1排序轴解释量为93.83%,第2排序轴解释量为3.97%,即前2轴对酶活性特征解释值为97.80%。表明前2轴能较好的解释土壤酶活性与环境因子的关系,并且主要由第1排序轴决定。第1、第2排序轴中,水热条件和植物多样性因子与土壤酶活性的相关系数分别为0.999和0.998,进一步反映出土壤酶活性与环境因子关系密切。
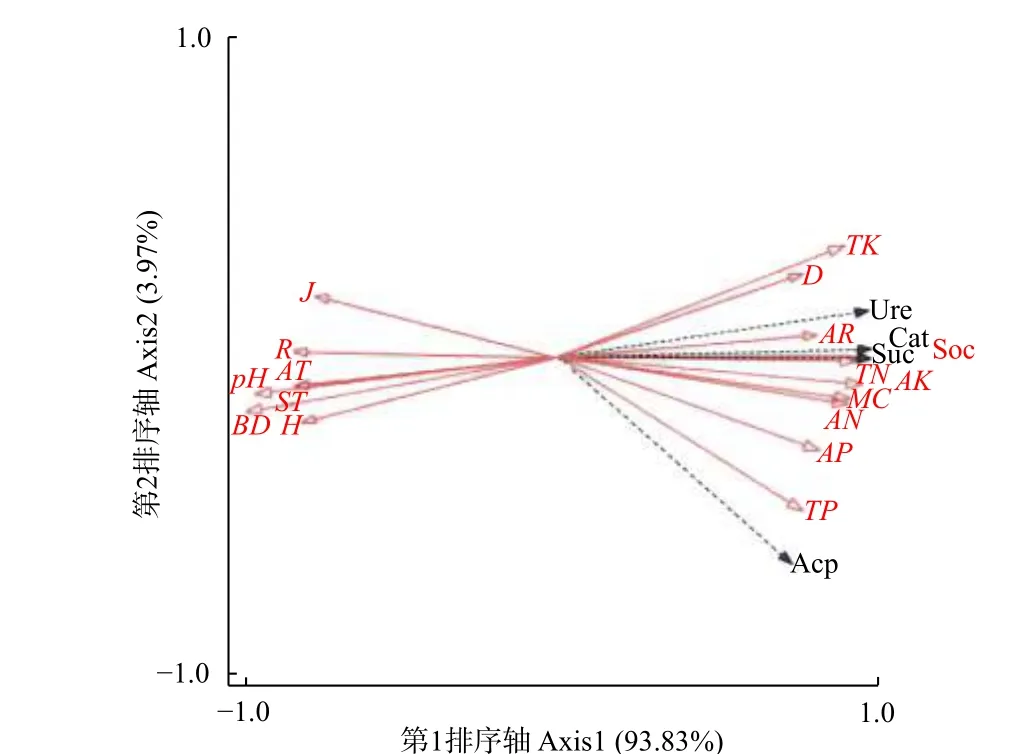
图 3 土壤酶活性与环境因子的冗余度分析Fig. 3 Redundancy analysis of soil enzyme activity and environmental factors
从土壤酶活性与环境因子的二维排序图(图3)可知:土壤全钾(TK)、pH和容重(BD)的箭头连线较长,能较好解释土壤酶活性的差异,其中,Margalef(R)、Shannon-Wiener(H)、Pielo-u指数(J)及年均气温(AT)、土壤温度(ST)与过氧化氢酶(Cat)、脲酶(Ure)、蔗糖酶(Suc)和酸性磷酸酶(Acp)夹角较大(> 90°),呈显著负相关(P< 0.05);Simpson指数(D)、土壤全钾(TK)、速效钾(AK)、含水量(MC)、速效氮(AN)、速效磷(AP)与上述4个酶夹角较小(< 90°),呈显著正相关,其中,过氧化氢酶、蔗糖酶与总有机碳(SOC)和全氮(TN)夹角最小,脲酶与年均降水量(AR)夹角最小,酸性磷酸酶与全磷(TP)夹角最小,表明总有机碳和全氮对过氧化氢酶、蔗糖酶存在极显著正效应(P<0.01),年均降水量对脲酶存在极显著正效应,全磷对酸性磷酸酶存在极显著正效应,而4种酶活性与pH和容重(BD)夹角最大,说明4种酶活性与pH和容重存在显著负效应。
综上可知:不同环境因子对土壤酶活性的影响差异较大。使用蒙特卡罗检验对17个因子进行大小排序得出:总有机碳(SOC)> 全氮(TN)> 容重(BD)> 速效钾(AK)>pH> 含水量(MC)>速效氮(AN)> 全钾(TK)> 有效磷(AP)> 全磷(TP)> Pielou指数(J)> 年均降水量(AR)>年均气温(AT)> 土壤温度(ST)> Simpson指数(D)> Margalef index指数(R)> Shannon-Wiener指数(H)。相关分析显示:各环境因子对土壤酶活性均影响显著(P< 0.05)。
3.4.2 单一因子对酶活性的影响 根据蒙特卡洛检验排序的大小,使用T-value双序图筛选出对土壤酶活性有显著影响的12个环境因子进行深入分析,以明确酶活性与主要环境因子的关系。由图4a、b、d可知:脲酶(Ure)、过氧化氢酶(Cat)、蔗糖酶(Suc)和酸性磷酸酶(Acp)的箭头均穿过了黑圈,说明土壤总有机碳(SOC)、全氮(TN)速效钾(AK)与上述4个酶均呈显著正相关,对各酶活性影响较大。由图4f~h、l可知:过氧化氢酶、脲酶、蔗糖酶的箭头穿过了黑圈,说明土壤含水量(MC)、速效氮(AN)、全钾(TK)和年降水量(AR)和上述3个酶均呈显著正相关,对3个酶活性变化起促进作用。由图4c、e、k可知:脲酶、过氧化氢酶和蔗糖酶的箭头均穿过了白圈,说明容重(BD)、pH和Pielo-u指数(J)与脲酶、过氧化氢酶和蔗糖酶呈显著负相关,并对3个酶反向变化的影响较明显。由图4i、j可知:酸性磷酸酶箭头完全落入了黑圈,说明速效磷(AP)和全磷(TP)与酸性磷酸酶呈极显著正相关,表明速效磷和全磷是影响酸性磷酸酶的正向变化主要因子。由图4c、l可知,酸性磷酸酶箭头完全落入白圈,说明容重(BD)和年均降水量(AR)与酸性磷酸酶呈极显著负相关,表明年均降水量和土壤容重对酸性磷酸酶反向变化的影响最明显。
4 讨论
4.1 不同植被带土壤酶活性沿海拔的变化
高原山区垂直气候沿海拔梯度的变化,能够引起植被类型、凋落物输入质与量、土壤微生物及理化性质的显著改变,从而形成土壤酶的结构及活性在不同海拔高度的显著差异性[17]。本研究中,土壤过氧化氢酶、脲酶及蔗糖酶的活性均随海拔升高呈显著增强的趋势,且湿性常绿阔叶林的最大。湿性常绿阔叶林郁闭度最大,凋落物输入量大,根系及分泌物较多、腐殖质层较厚,导致土壤碳、氮及磷等养分含量的显著增加,其分解中释放的土壤中有机质、养分元素可以为酶促反应提供良好的基质[18]。土壤过氧化氢酶与蔗糖酶随海拔的增加,能够促进土壤有机物的转变与积累[19],土壤脲酶的增加能够分解土壤中的尿素,提高土壤的供氮能力[20]。因此,土壤酶活性随海拔的增加可能促进高海拔样地土壤养分循环、土壤生物活动、植物生长及群落演替。
土壤酸性磷酸酶参与土壤磷循环和代谢,其活性的高低能够反映土壤磷的生物转换方向及强度[21]。本研究中,土壤酸性磷酸酶活性随海拔升高呈先减小后增强的变化趋势,这与任玉连等[9]研究结果不同,这可能与不同地区土壤空间异质性有关。半湿润常绿阔叶林土壤磷含量显著低于亚热带季风常绿阔叶林和湿性常绿阔叶林,表明受到的磷限制程度低于其它2种植被带,无需微生物分泌更多的磷酸酶来维持速效磷供给[22]。因此,土壤酸性磷酸酶活性沿海拔的变化反映了植物生长需求及土壤供磷水平之间的平衡关系。
不同植被带相同土层酶活性随海拔上升呈增加趋势。一方面,随着海拔上升,同一土层容重变小,根系分布逐渐增多,植物根系分泌物的增加促进土壤微生物的代谢产酶能力[23];另一方面,相同土层酶活性的海拔变化规律与养分元素一致,表明高海拔植被带土壤剖面的水热条件和肥力状况良好,促进了土壤酶活性。在土壤层次上,由于受生物、淋溶等共同作用,使土壤中养分物质在不断地进行交换,导致各土层间土壤理化性质的显著差异[24],从而对土壤酶活性的垂直分布造成影响。研究发现,各海拔植被带下4种土壤酶活性沿土层逐渐减小,这与土壤含水量和土壤碳、氮和磷等养分含量垂直变化趋势相一致。因此,不同海拔土壤酶活性垂直变化与土壤微生物及土壤养分沿土层分布密切相关。
4.2 不同海拔水热条件及植物多样性对土壤酶活性的影响
海拔是重要的山地地形因子之一,海拔的变化会引起水热条件的梯度变化[25],Zuccarini等[26]研究发现,土壤水分会对酶活性产生显著的影响,升温时只有伴随充足的水分条件才能增加土壤酶活性。本研究发现,降水量和土壤含水量对脲酶、过氧化氢酶和蔗糖酶存在显著正向影响,但降水量的增多会对酸性磷酸酶产生显著负效应。这与酶的底物变化有关,水分沿海拔的增加能够促进凋落物分解和高分子化合物形成[27],积累的有机质增加了土壤脲酶、过氧化氢酶和蔗糖酶的反应底物,但大量降雨加剧了土壤磷素的淋溶作用,导致研究区整体处于低磷状态(全磷均值为1.29 g·kg-1),使磷酸酶对磷的底物利用率较低。前人研究表明[28-29],温度可直接或间接影响酶活性,适宜的温度对酶活性具有促进作用。本研究“环境因子对酶活性影响的排序”表明,土壤温度对酶活性总体解释性较弱,低海拔植被带较高的土壤温度未能显著影响到土壤酶的活性。可能由于文山低海拔植被带日照时间长,强光合作用促进植被呼吸速率[30],导致生境热量循环加快,刺激了微生物对土壤碳氮等酶促底物的分解速率,进而限制低海拔植被带土壤酶活性。
植被多样性不仅影响着森林群落结构组成,还对土壤基质及养分库起稳定作用[31],从而引起土壤酶活性对植被多样性的变化做出响应。本研究中,植被多样性沿海拔梯度对土壤酶活性产生不同程度的负向影响,其中,均匀度指数最明显,这与杨秉珣等[32]研究结果相似。保护区主要以典型亚热带阔叶林为主,群落组成差异较小,均匀度指数对群落分布的影响较其他指数强,随着海拔上升,植被由草本、灌木群落向乔木群落进行演替,地表覆盖物和地下根系腐解物逐渐增多,伴随着低温环境,微生物的呼吸作用和分解能力受到抑制,土壤养分物质以积累为主[33],从而使土壤酶的底物和活性随海拔上升均有不同程度的增加。
4.3 不同海拔植被带土壤理化性质对土壤酶活性的影响
土壤理化性质影响着土壤酶的活性及稳定性[34],但不同土壤理化因子对土壤酶活性影响程度有差别。土壤有机质和矿质元素是土壤中酶促底物的主要供源,是土壤酶的载体[35]。本研究中,土壤有机质和矿质元素(氮、磷、钾)对土壤各酶活性存在显著的正效应,其中,总有机碳和全氮最明显,表明土壤养分沿海拔的增加促进了样地酶活性,这与许全等[36]研究结果相似。
土壤pH能够决定土壤有机分子功能基团的离子化,影响酶和底物的构象[28],本研究发现,土壤pH与各个酶活性呈显著负相关,表明土壤酶活性随土壤pH的升高而降低,这与Sinsabaugh等[37]等研究结果不同,这可能是由土壤的空间异质性,造成土壤pH和酶活性的关系不同所致。土壤容重通过改变土壤紧实度来影响土壤水分条件、空气组成和热量状况,进而调控土壤酶活性[38]。研究表明,土壤容重与各酶活性呈显著负相关,表明土壤酶活性随土壤容重增加而降低,这与吕瑞恒等[6]和马剑等[7]研究结果相似。
土壤理化性质的海拔差异对土壤酶活性分布规律产生重要影响,土壤酶活性的增强,则能够改变土壤理化性质,有利于土壤养分元素的积累,促进植物生长及群落演替[39]。因此,文山国家级自然保护区“海拔高度-植被群落-土壤理化性质-土壤酶”之间存在复杂的耦联关系。
5 结论
不同植被带土壤酶活性沿海拔梯度呈现不同的变化特征,较高海拔的湿性常绿阔叶林土壤酶活性显著高于较低海拔的亚热带季风常绿阔叶林与半湿润常绿阔叶林群落。土壤过氧化氢酶、脲酶和蔗糖酶活性随海拔升高呈增加趋势,而土壤酸性磷酸酶活性则呈先下降后上升的趋势。不同海拔植被带各土壤酶活性均随土层加深而降低。
冗余分析和单一因子检验表明,各环境因子对酶活性具有不同的影响。土壤有机质和矿质元素(氮、磷、钾)是调控土壤酶活性变化的主要因子;容重和pH与土壤酶活性呈显著负相关,表明较低的容重和pH可能有利于提高土壤酶的活性;植被多样性和水热条件的海拔差异是驱动酶活性变化的重要因子。
猜你喜欢
生物技术通报(2020年12期)2020-12-21
腐植酸(2020年1期)2020-11-29
绿色中国(2019年18期)2020-01-04
浙江理工大学学报(自然科学版)(2018年6期)2018-11-07
农家科技下旬刊(2017年8期)2017-11-13
中国组织化学与细胞化学杂志(2017年1期)2017-06-15
科技创新导报(2016年21期)2016-12-17
天然产物研究与开发(2016年6期)2016-06-05
医学研究杂志(2015年8期)2015-06-22
中国医学科学院学报(2013年6期)2013-03-11
