
丛枝菌根真菌对植物繁殖的影响研究进展
2021-01-07 05:13王磊闫兴富唐占辉
广西植物 2021年12期
王磊 闫兴富 唐占辉


摘 要: 叢枝菌根真菌(arbuscular mycorrhizal fungi, AMF)与宿主植物所形成的互惠共生体系是生态系统中广泛分布的共生体系代表类型之一。AMF除能够促进宿主植物生长发育外,也可以对宿主植物的繁殖过程产生多方面影响。研究宿主植物在AMF共生状态下繁殖策略的变化规律,对于深入理解植物繁殖适合度的变化具有重要理论意义。该文综述了AMF对宿主植物繁殖影响的相关研究,包括AMF的侵染对宿主植物繁殖分配、花部特征、虫媒传粉和花期的影响。目前已有研究发现某些AMF能够促进宿主植物增加繁殖资源投入,提高花朵产生的数量或花冠直径,同时增加花粉数量和花蜜量来影响访花昆虫的行为,以及造成开花提前及花期延长,但其作用的具体机制尚不明确,且因宿主植物的差异,并未有完全统一的结论。然而,由于AMF与植物共生的普遍性,其在植物繁殖过程中发挥的重要作用不可忽略。今后除了在以上各方面开展更深入的研究外,还需在AMF对宿主植物繁殖性状的影响机制、AMF共生条件下植物有性繁殖过程中雌雄功能的资源分配,以及对无性繁殖和后代适应性的影响等方面进行更深入的研究。
关键词: 丛枝菌根真菌(AMF), 繁殖分配, 花期, 花部综合征, 传粉
中图分类号: Q948
文献标识码: A
文章编号: 1000-3142(2021)12-2110-11
收稿日期: 2021-01-18
基金项目: 国家自然科学基金 (31470446,31960228) [Supported by the National Natural Science Foundation of China (31470446,31960228)]。
作者简介: 王磊(1996-),硕士研究生,研究方向为植物繁殖生态学,(E-mail)wangl788@nenu.edu.cn。
通信作者: 唐占辉,博士,副教授,硕士研究生导师,研究方向为植物繁殖生态学,(E-mail)tangzh789@nenu.edu.cn。
Research progress on the effects of arbuscular
mycorrhizal fungi on plant reproduction
WANG Lei1, YAN Xingfu2, TANG Zhanhui1*
( 1. State Environmental Protection Key Laboratory of Wetland Ecology and Vegetation Restoration/School of environment, Northeast Normal
University, Changchun 130117, China; 2. College of Biological Science and Engineering/Key Laboratory of Ecological Protection of
Agro-pastoral Ecotones in the Yellow River Basin,
National Ethnic Affairs Commission of the People’s Republic
of China, Beifang Minzu University, Yinchuan 750021, China )
Abstract: The mutually beneficial symbiosis system formed between arbuscular mycorrhizal fungi (AMF) and host plants is one of the representative types of symbiosis systems widely distributed in various ecosystems. In addition to promoting the growth and development of host plant, AMF can also have various effects on the reproduction process of host plants. Studying the changes of reproduction strategies of host plants infected by AMF has important theoretical significance for in-depth understanding of changes in plant reproductive fitness. In this article, we review the related researches on the effects of AMF on the reproduction of host plant, including the effects of AMF infection on reproductive allocation, floral characteristics, insect-pollination and anthesis of host plants. Although current studies have found that AMF can promote the increase of host plant reproduction input, increase the size and number of flowers, increase the number of pollen and nectar secretion, affect the behavior of flower visitors, and cause early flowering and prolonged flowering, the specific mechanism of AMF affecting reproduction of flowering plant is still unclear, and there is currently no unified conclusion due to differences between host plants. However, due to the ubiquity of AMF symbiosis with plant, its important role in the process of plant reproduction cannot be ignored. In the future, more researches should be conducted to investigate the influence mechanism of AMF on the reproductive traits of host plants, the resource allocation of male and female functions in the process of plant sexual reproduction, asexual reproduction and offspring fitness in addition to conducting more in-depth studies in the above-mentioned aspects.
Key words: arbuscular mycorrhizal fungi (AMF), reproductive allocation, anthesis, floral traits, pollination
丛枝菌根真菌(arbuscular mycorrhizal fungi, AMF)是自然界大量存在的能够侵染植物根系且与其形成菌根共生体的土壤真菌。AMF菌丝与宿主植物营养根系共生所形成的丛枝菌根是生态系统中广泛分布的植物-真菌互惠共生体系的代表,其对植物群落生产力的形成和生物多样性的维持具有重要作用(Oehl et al., 2011;黄艳飞等, 2019)。已有研究表明,全球约80%的陆生植物能够与AMF形成共生体系,且部分植物对AMF具有高度的依赖性(Berruti et al., 2016)。在AMF与宿主根系形成的共生体系内,AMF菌丝一端侵染进宿主根系表皮细胞内形成丛枝结构,用无机化合物(氮和磷酸盐)与宿主细胞的碳水化合物和脂肪酸进行交换以及分泌菌根因子激活宿主相关基因的表达(Jiang et al., 2017; He et al., 2019),另一端则形成根外菌丝扩大宿主根系吸收养分的范围(Smith SE & Smith FA, 2011)。AMF从宿主体内获取含碳有机物以满足自身的生长和繁殖,研究显示宿主体内约20%的光合产物用于AMF的消耗,其中一部分用于AMF自身的生长代谢,另一部分则以小分子有机化合物的形式释放到土壤中,以诱导菌根际细菌在菌丝附近或表面定殖,从而改善土壤微生物群落结构,进而改变宿主植物根际微环境(Hoeksema et al., 2010;Bharadwaj et al., 2012)。AMF的共生可为宿主根系提供更多的营养物质和水分,增大宿主对营养物质的吸收和利用效率。此外,AMF也可以通过影响宿主体内相关代谢途径和基因的表达等调节宿主植物体内激素的合成与分配,促进宿主植物的生长和发育(Zhang et al., 2014; Battini et al., 2016; 祝英等, 2016)。因此,两者在长期的协同进化过程中所形成的互惠互利的共生关系,对植物生长乃至繁殖发挥着重要的调控作用。
繁殖是植物生命活动中的重要环节,是植物种群更新和扩散的关键过程,其在维持植物群落结构稳定和生态系统功能等方面具有重要的作用(郭伟等, 2010; 孙士国等, 2018)。植物繁殖过程中的资源分配策略(营养器官与繁殖器官的资源投入比例)、繁殖方式(有性繁殖与无性繁殖)、花部综合特征演化和传粉等研究一直都是植物繁殖生态学关注的热点问题(张婵等, 2020)。植物繁殖性状易受到周围环境中生物和非生物因素的显著影响,从而导致其繁殖性状及繁殖策略发生不同程度的改变,以便响应这些因素的变化(Li & Zhang, 2019; Tang, 2020; Kenny et al., 2020)。AMF可以通过改善宿主植物对环境中营养元素的吸收能力,促进宿主植物营养器官生长和生物量增加,同时可以影响宿主体内某些次生化合物的合成,而这些促进效应和次生化合物的产生会进一步对宿主植物的繁殖性状产生一定的影响(张延旭等, 2019; Bennett & Meek, 2020),尤其对植物繁殖分配、花部特征、昆虫传粉过程和花期等方面的影响,可直接或间接改变宿主植物的繁殖适合度,有利于提高植物对环境的适应能力(Scagel & Schreiner, 2006; 胡文武, 2015; Jimenez-Leyva et al., 2017; 宋洁蕾等, 2019)。
AMF作为土壤中不可忽略的重要生态因子,其对植物繁殖过程的影响受到越来越多的关注。本文综述了近年来有关AMF对宿主植物繁殖影响方面的相关研究,包括AMF的侵染对宿主植物繁殖分配、花部特征、虫媒传粉以及花期这4个与植物繁殖机制密切相关的表观特征的影响,以及对其内在的影响机制进行了总结,并对今后AMF对植物繁殖适合度影响的研究工作做出了展望,以期为植物与菌根真菌共生影响植物繁殖的相关研究提供参考。
1 AMF对植物繁殖分配的影响
植物繁殖分配是指植物将总资源量投入到繁殖结构中(花和果实或种子)的比例,繁殖分配是植物资源分配策略的核心内容之一(Tian et al., 2012; 张婵等, 2020)。AMF促进宿主植物生物量的增加,可直接或间接影响宿主的繁殖分配,最终影响其繁殖适合度(Merlin et al., 2020)(图1)。有研究已证实接种AMF后的紫茎泽兰(Ageratina adenophora)在显著增加生物量的同时也增加了其花序和种子数量(宋洁蕾等, 2019)。同样,在接种AMF后,玉米(Zea mays)干物质产量显著提高的同时,果穗质量也有一定程度的增加(Sabia et al., 2015)。在室外条件下,接种AMF可显著提高番茄(Solanum lycopersicum)果实的产量(Miranda et al., 2015; Bona et al., 2017),但也有研究指出AMF对温室条件下生长的番茄果实产量并没有明显的影响(Schubert et al., 2020),可能的原因是在室外条件下生长的番茄受到磷酸鹽的限制,从而增加菌根以适应养分缺乏的外部环境,而实验室条件下的高营养环境可能会限制菌根的形成。众所周知氮和磷是限制植物营养生长和繁殖的重要因素,而AMF可以改善其宿主对氮和磷的吸收状况,从而在一定程度上可以促进宿主对繁殖的资源投入(Koide & Dickie, 2002),如碱蓬(Suaeda physophora)在接种AMF后地上部分的磷含量明显提高,其种子数量显著增加(Yang et al., 2016)。Campo et al.(2020)研究了AMF接种对水稻(Oryza sativa)生长、籽粒数量和穗数的影响,发现AMF侵染后的水稻生长更好,且籽粒数和穗数显著增加,这可能与AMF促进细胞分裂素等有利于植物花芽分化的激素合成有关。
大量研究结果发现接种AMF可以提高宿主植物对繁殖资源的投入,可使种子产量增加,但也有研究发现接种AMF并不会明显提高植物的种子产量(表1),如对大豆(Glycine max)单独接种AMF后发现其种子数量、直径、表面积等并没有受到显著影响。AMF对宿主植物种子产量的影响受到植物与AMF间的亲和力以及它们与土壤中某些微生物协同增效作用的影响(Igiehon & Babalola, 2021)。值得注意的是,在接种AMF后番薯(Ipomoea batatas)的块根和马铃薯(Solanum tuberosum)的块茎产量有了明显的增加,并显著提高了匍匐茎形成块茎的比例,这表明AMF对宿主植物的无性繁殖也会产生影响,能够在一定程度上使宿主植物加大对无性繁殖的资源投入(Yu et al., 2013; Nurbaity et al., 2019),但这种效应并不普遍且因宿主而异,藏红花(Crocus sativus)在AMF侵染后其作为无性繁殖器官的球茎(大小和直径)就无显著变化(Caser et al., 2020)。虽然AMF能够促进宿主植物对土壤中氮、磷等营养元素的利用和光合产物的积累,促进宿主植物生长及生物量积累,从而促使宿主植物在繁殖上的资源投入,但更多的是从种子的绝对增产方面进行了报道,很少用繁殖分配这个相对指标来衡量AMF对宿主植物繁殖资源分配方面的影响。当前,宿主植物的繁殖分配对AMF侵染的响应机制仍缺乏深入探讨,并且克隆繁殖的植物对无性繁殖的资源投入如何响应AMF侵染的研究尚有所欠缺。此外,AMF的定殖能够增强宿主植物抵抗土壤真菌、细菌、病毒和其他生物(如植食性线虫)的能力(Affokpon et al., 2011),这在一定程度上也可能会对宿主植物的繁殖分配策略产生一定的影响,这方面的研究也需进一步关注。
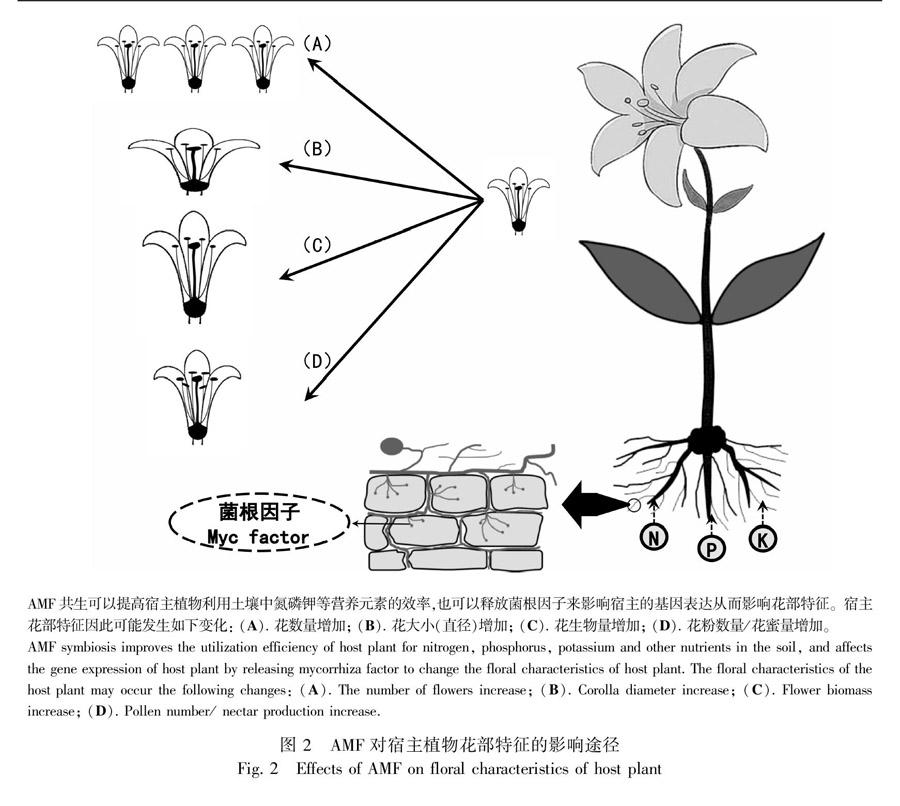
2 AMF对植物花部特征的影响
花作为植物重要的有性繁殖器官,其形态特征对访花者访花和花粉传递过程具有重要的影响(雷虓等, 2019)。AMF与宿主植物的共生可以改变植物花的大小(直径)、开花数量等花部特征(Bennett & Meek, 2020)(图2)。多数研究支持AMF侵染能够增加植物花的大小或数量(表1),如接种AMF明显增加了曼陀罗(Datura stramonium)花的数量(Aguilar-Chama & Guevara, 2012),在番茄和马蹄莲(Zantedeschia spp.)接种AMF的研究中也得到了相似的结果(Bryla & Koide, 1990; Scagel & Schreiner, 2006)。对于大多数植物而言,由于资源的限制作用,植物花的数量与植株生物量有一定相关性,AMF与植物共生后可增强宿主植物对磷元素的吸收,促进植株营
养生长和生物量积累,从而使宿主提高了花的生产,可见AMF共生后对植物所提供的磷元素增加是引起植物繁殖资源向花器官投入的重要因素之一(Scagel & Schreiner, 2006)。除此之外,植物体内钾含量的增加也被认为可以促使花数量的增加(Dufault et al., 1990),而AMF的侵染可以提高宿主植物对土壤中钾的吸收,从而提高宿主体内钾含量,间接使宿主花数增加(Gaur et al., 2000),且相关研究表明AMF共生后可诱导宿主细胞钾转运体基因的表达,促进宿主对钾的吸收(Liu et al., 2019)。Bona et al.(2015, 2017)认为AMF是通过调控宿主植物体内的光合产物和植物生长素(IAA)的含量,使宿主植物花数量增多。IAA作为AMF与宿主植物共生的重要信号物质,是调节植物生长发育的重要植物激素,有研究表明,AMF能够使IAA合成关键酶基因的表达上调,增加宿主体内IAA的含量,从而促进宿主花数的增加(赵方贵等, 2019)。植物花数的增加在一定程度上可能吸引更多的传粉者来帮助其传粉,有助于提高传粉效率。由此可见,AMF可以通过影响宿主植物的花部特征来提高其对环境的适应性(图1)。
也有研究认为,AMF对宿主植物的花数没有直接影响或在某种程度上减少了宿主植物的花数(表1)。Püschel et al.(2014)对多种植物接种AMF后的花数进行统计发现,辣椒(Capsicum annuum)、Gazania splendens、新几内亚凤仙(Impatiens hawkerii)、盾叶天竺葵(Pelargonium peltatum)、马蹄纹天竺葵(P. zonale)等植物的花数并没有显著变化,而贯叶连翘(Hypericum perforatum)在接种AMF后单株花的数量减少(Lazzara et al., 2017)。Bolin et al.(2018)發现Clarkia xantiana在接种AMF后花数量也有所降低,分析认为可能的原因是AMF对宿主植物具有一定的偏好性,在某些植物中表现为共生效应,在另一些植物中可能表现为寄生效应,花数的减少是由于AMF的寄生作用使得宿主植物对繁殖资源的分配减少,说明某些AMF表现为偏害共生作用。AMF对宿主植物花朵数量的影响可能既依赖于AMF与宿主植物之间对氮和光合产物的竞争,又依赖于AMF增强了宿主植物对营养不足或其他环境胁迫的响应能力,而这些依赖性又可能取决于环境或实验条件下土壤基质的氮磷比(N/P)。我们认为,AMF与宿主植物的亲和力、植物生长基质条件以及外在胁迫是造成AMF侵染后宿主植物花数产生变化的主要影响因素,今后这方面的研究要特别重视这些影响因素的共同作用。
AMF对宿主植物花部特征的影响还体现在对花的大小(花直径)和单花质量两个方面(表1,图2)。万寿菊(Tagetes erecta)在接种AMF后花的直径增加了约11%,花直径的增加间接促进了传粉者对宿主访花频率的提升(Gange & Smith, 2005)。一些研究也发现接种AMF处理后的马蹄纹天竺葵、金盏菊(Calendula officinalis)和鸟尾花(Crossandra infundibuliformis)的花序干重均有明显的增加(Vaingankar & Rodrigues, 2015; Giulia et al., 2015; Heitor et al., 2016)。此外对千叶蓍(Achillea millefolium)、Aster laevis、圆叶风铃草(Campanula rotundifolia)、春飞蓬(Erigeron philadelphicus)、Solidago missouriensis的生长基质施用杀真菌剂后,这些植物的花序大小等没有显著改变,但相同处理的卷耳(Cerastium arvense)花序大小则均有所增加,这表明了AMF对植物花大小的影响因物种而异(Cahill et al., 2008)。花大小的改变可以影响访花昆虫的种类,在一定程度上影响植物-传粉者间的相互作用,改变植物的繁殖适合度。
虽然AMF对某些植物表现为寄生效应,但是这种寄生效应可能在一定程度上促进宿主植物在繁殖器官上的资源投入以满足自身的繁殖需求(Heitor et al., 2016)。实际上,AMF定殖对花数量或大小的影响也可能取决于植物的繁殖系统(Nicholas & Nicole, 2015)。AMF定殖可增加Geranium sylvaticum两性花的花朵大小(Varga & Kytviita, 2010),而另有一项以Cucurbita foetidissima为材料的研究结果则显示AMF仅仅促进其雄花数的增加(Pendleton, 2000)。此外AMF定殖也可增加雌雄同株植物黄瓜(Cucumis sativus)的雄花大小(Kiers et al., 2010)。AMF对植物花部特征的影响在不同物种间表现出较大的变异性和多样性, 这取决于AMF的种类以及宿主植物的特性,目前这方面的研究还不足以支撑总结出一般的普适规律。特别是对具有复杂性系统类型的植物,AMF对雌雄同株性系统植物中两性花和单性花,以及雌雄异株性系统植物中雌花和雄花的花部特征影响可能存在更大的差异。性系统复杂的植物,不同的基因型个体在利用资源方面的策略也有所不同,在此基础上,AMF的共生可能会进一步增加其花部特征的变化和多样性,这方面在将来的研究中也应该特别关注。
3 AMF对植物虫媒传粉的影响
植物有性繁殖时花粉成熟以后会通过自然界中的各种传粉媒介将花粉传递到柱头,根据传粉媒介的不同可将有性繁殖植物的花分为风媒花、水媒花和虫媒花等(任明迅等, 2004)。传粉行为在植物的繁殖过程中具有重要的地位,若植物的传粉过程受到限制则会使植物结实率降低,导致植物种群更新受阻甚至濒危(王爱芝等, 2020)。风媒植物和水媒植物的传粉均依靠自然作用力进行,而虫媒植物吸引传粉者则依赖其繁殖器官的性状。植物吸引传粉者的方式大体上可以分为以下几种:(1)通过醒目的花朵展示来吸引传粉者进行传粉;(2)通过花朵散发的特殊气味来吸引传粉者傳粉;(3)花朵具有特殊的“蜜导”来吸引和指示传粉者在吸食花蜜的过程中完成传粉;(4)通过特化的传粉系统来吸引特定的传粉者进行传粉或为传粉者提供一定的回报以吸引传粉者为其传粉(Waser & Price, 1981; Micheneau, 2006; 顾垒和张奠湘, 2009; 张婵等, 2012; 邢剑锋等, 2015; 张亭等, 2018)。本文前面已经述及AMF可以改变植物花序或花的数量和大小,而这些变化均可以间接影响植物对传粉者视觉上的吸引,从而影响传粉者的活动和访问频率。
植物花的颜色对传粉者来说也是一个重要的视觉信号,花色由植物次生代谢产物花色苷含量决定,包括酚类化合物花青素和类黄酮类物质(Weston & Mathesius, 2013)。在AMF共生情况下有些植物会改变次生代谢产物的产生,特别是改变与植物花朵颜色相关的次生代谢产物合成,进而可影响植物的花色(Bennett & Cahill, 2018)。一些研究发现接种AMF可以提高植物体内酚类、黄酮类等化合物的合成而影响植物花朵的颜色(Chen et al., 2013; Zhang et al., 2013)。AMF接种可影响番茄、辣椒、草莓(Fragaria × ananassa)、莴苣(Lactuca sativa)花青素和类黄酮等色素类物质的含量,并在一定程度上影响植物的花色(Luciano et al., 2018)。另外,AMF的侵染也会促进宿主植物花内次生挥发性有机物的生成(Rasmann et al, 2017; Bennett & Meek, 2020),如贯叶连翘与AMF共生后花朵中的主要次生化合物假金丝桃素和金丝桃素含量有所增加,改变了植物花朵中挥发性化合物的含量(Lazzara et al., 2017)。花的气味在很大程度上是由其挥发性次生化合物决定,而花朵所散发的特殊气味是植物吸引传粉者的一个重要方式。因此,AMF与植物共生后促进花气味的微小变化将会影响访花者的行为,并在一定程度上改变花朵的传粉效率以及子代的结实率。AMF改变宿主植物次生代谢产物以及相关化合物的量从而调控植物花色的机制,以及影响访花者的行为途径目前还不清楚。
AMF为宿主植物所提供的营养条件在一定程度上也会促进植物花粉数量和花蜜量增加(表1)。在花粉的形成过程中需要磷的参与,由于AMF共生可以促进植物对土壤中磷元素的吸收,因此,AMF的共生作用可促进宿主植物雄性功能的提高。有研究发现,AMF侵染能够显著增加西葫芦(Cucurbita pepo)单花花粉产量和花粉粒径(Lau et al., 1995),同样千屈菜(Lythrum salicaria)在AMF侵染后其单花花药和花粉数量均有所增加(Philip et al., 2001),表明AMF侵染能够提高植物的雄性功能,从而有助于植物增强花粉传递和种群间的基因流动。AMF侵染某些植物后,也会增加植物雄蕊的数量从而增加花粉数量,来提高植物的雄性适合度(Varga & Kytviita, 2010; Pereyra et al., 2019)。
花蜜在植物与传粉者的关系中也扮演着重要角色(Gehrels et al., 2004),AMF可通过影响植物体内次生化合物(如茉莉酸和水杨酸)的产生从而影响花蜜的产生量(Jacobsen & Raguso, 2018)。如AMF侵染万寿菊形成菌根后增加了其花蜜产量(Gange & Smith, 2005),可能的原因是菌根增强了植物的固碳作用从而导致花蜜产量增加,进而有助于吸引传粉者的拜访。但也有不同的研究结论,如匐根花荵(Polemonium viscosum)生长在没有AMF存在的基质中,其花蜜量显著高于含有AMF的对照组(Becklin et al., 2011),说明在含有AMF的基质中植物分泌花蜜量较少,这在一定程度上可能会降低花对传粉者的吸引力。降低花蜜量也能减少蚂蚁、鸟类等盗蜜者的访问,也可能是植物有效降低非传粉者盗蜜风险的一种间接性防御策略。对于显花植物来说,增加花粉和花蜜的量可在一定程度上提高传粉者的访问频率而促进传粉。AMF的共生效应在改善宿主个体营养条件的同时也促进了植物在花粉和花蜜上的投资,最终使传粉者访问频率增加,从而提升植物的传粉效率。但是也要注意到AMF的侵染如果降低植物花蜜的产量或改变了花蜜的化学成分,则可能有着更为复杂的生态学意义,值得更深入的探究。
4 AMF对植物花期的影响
植物的开花在很大程度上是由其生存的微环境特征和遗传因素决定的(包宇, 2018)。微环境特征包括多种生物因子(个体间竞争、传粉者活动、土壤微生物等)和非生物因子(温度、海拔、光照和降水等),它们在一定程度上影响着植物的开花进程(Wright & Meagher, 2003; Elzinga et al., 2007; Craine et al., 2012; Davies et al., 2013; Wang et al., 2015; 李慢如和张玲, 2019; 陶泽兴等, 2020)。AMF作为土壤微生物类群中活跃且变化较大的生物因子之一,其对植物花期有直接或间接影响。因为AMF共生可以促进植物的生长发育,而许多植物的开花与其个体大小有一定程度的关联(Otarola et al., 2013),所以AMF可能通过促进植物生长而间接改变植物的开花进程,个体大小与花期的变化可以共同影响植物的繁殖适合度(图1)。
目前,关于AMF侵染后植物开花是否提前和花期是否延长并没有一致的结论,大部分研究认为植物接种AMF后花期会提前(表1),如利用AMF分别侵染番茄、苘麻(Abutilon theophrasti)和三色魔杖(Sparaxis tricolor)后,3种植物的开花时间均明显提前(Bryla & Koide, 1990; Lu & Koide, 1994; Scagel, 2004a)。最近在蒺藜苜蓿(Medicago truncatula)和勋章菊(Gazania rigens)等植物中也得到了类似的结论(Liu et al., 2018; Saini et al., 2019),这些研究结果认为AMF可能促进了植物对钾的吸收,导致生长素和赤霉素合成增加,从而诱导花芽的分化与形成(Asmelash et al., 2016)。也有研究发现接种AMF对宿主植物的开花时间没有影响或延后宿主植物開花时间,以海洲香薷(Elsholtzia splendens)为材料的研究中并没有发现AMF促进其开花提前的现象(Jin et al., 2015),在风信子(Hyacinth orientalis)的研究中也发现接种不同的AMF对开花没有影响,甚至开花还略有延迟(Xie & Wu, 2018)。AMF侵染的另外一个重要影响是改变植物的花期长短,较为一致的结论是AMF侵染促进了宿主植物花期的延长(表1),但是这种促进作用并不是绝对的(Liu et al., 2018; Xie & Wu, 2018; Saini et al., 2019)。如AMF可使Brodiaea laxa的开花时间提前,却不影响其花期长短(Scagel, 2004b)。AMF对宿主植物花期的影响可能是通过增加宿主植物本身的营养条件,从而使植物有更多的资源可以投入花生产上,促进植物开花提前以及改变花期长短。植物花期的延长在一定程度上能够增加植物有性繁殖的成功率,扩大基因交流的范围,有助于提高植物对环境的适应性。目前还没有对由于AMF的存在而造成植物花期变化的规律进行合理解释的理论,然而可以明确的是,AMF的存在的确会影响宿主植物的花期,但具体的影响途径和机理尚不清楚。
5 展望
植物与AMF的共生效应一直以来都是研究生物互作的热点领域之一。本文综述了AMF共生对植物繁殖分配、花部特征、虫媒传粉、花期等方面的影响及其机制。但从已有的研究报道来看,国内外学者对AMF共生条件下宿主植物繁殖响应规律的研究还相对不足,主要表现在以下几个方面。首先,大多数研究是对植物接种单一的AMF研究其繁殖响应,缺乏多种AMF混合接种的实证研究;其次,已有研究大多集中在一年生植物上,而多年生植物较少,多年生植物因其具有较长的生活史周期可能对AMF的响应有更复杂的机制;再次,在AMF对宿主植物繁殖影响的机制方面缺乏深入解析,特别是AMF如何影响宿主植物内在基因表达以及调控过程从而影响其繁殖,这方面研究还有所欠缺。因此,未来的研究应该从以下方面考虑:(1)研究多种AMF共同作用下植物繁殖的响应,实际上混合菌种的接种可以更好地模拟自然条件,研究结果将更有意义。(2)增加AMF对多年生植物繁殖影响的研究,并扩大研究的时间尺度,跟踪AMF作用下多年生植物长期的繁殖过程。同时增加一年生植物在AMF共生条件下代际之间的繁殖响应研究,特别要关注AMF对宿主植物种子特性(包括产量、大小、活力等)的影响,由于种子会直接决定后代的适合度和竞争能力,进而对种群的扩散和延续产生一定的影响。(3)关注AMF共生后宿主植物有性繁殖和无性繁殖的权衡机制,以及有性繁殖过程中其雌雄功能的响应变化规律。(4)对AMF共生下宿主植物繁殖响应机制进行深入挖掘研究,阐明其生理机制以及分子调控原理等,包括AMF如何与宿主植物进行信息交流从而提高宿主植物体内激素水平或者次生代谢物的含量,以及如何影响宿主植物的基因表达,进而使宿主植物与繁殖相关的性状发生改变等。未来可通过对以上方面的深入研究,揭示在AMF共生后宿主植物繁殖策略的变化,以及其提高繁殖适合度和增强环境适应能力的机制。
参考文献:
AFFOKPON A, COYNE DL, LAWOUIN L, 2011. Effectiveness of native West African arbuscular mycorrhizal fungi in protecting vegetable crops against root-knot nematodes [J]. Biol Fert Soils, 47(2): 207-217.
AGUILAR-CHAMA A, GUEVARA R, 2012. Mycorrhizal colonization does not affect tolerance to defoliation of an annual herb in different light availability and soil fertility treatments but increases flower size in light-rich environments [J]. Oecologia, 168: 131-139.
ASMELASH F, BEKELE T, BIRHANE E, 2016. The Potential Role ofarbuscular mycorrhizal fungi in the restoration of degraded lands \[J\]. Front Microbiol, 7: 1095.
BAO Y, 2018. Study on reproductive ecology of Lychnis cognate Maxim and Lychnis wilfordii(Regel) Maxim [D]. Changchun: Northeast Normal University. [包宇, 2018. 淺裂剪秋萝和丝瓣剪秋萝的繁殖生态学研究 [D]. 长春: 东北师范大学.]
BATTINI F, CRISTANI C, GIOVANNETTI M, et al., 2016. Multifunctionality and diversity of culturable bacterial communities strictly associated with spores of the plant beneficial symbiont Rhizophagus intraradices [J]. Microbiol Res, 183: 68-79.
BECKLIN KM, GAMEZ G, UELK B, et al., 2011. Soil fungal effects on floral signals, rewards, and above ground interactions in an alpine pollination web [J]. Am J Bot, 98(8): 1299-1308.
BENNETT AE, MEEK HC, 2020. The influence of arbuscular mycorrhizal fungi on plant reproduction [J]. J Chem Ecol, 46: 707-721.
BENNETT JA, CAHILL JFJ, 2018. Flowering and floral visitation predict changes in community structure provided that mycorrhizas remain intact [J]. Ecology, 99(6): 1480-1489.
BERRUTI A, LUMIN E, BALESTRINI R, et al., 2016. Arbuscular mycorrhizal fungi as natural biofertilizers: Let’s benefit from past successes [J]. Front Microbiol, 6(426): 1-13.
BHARADWAJ DP, ALSTROM S, LUNDQUIST PO, 2012. Interactions among Glomus irregulare, arbuscular mycorrhizal spore-associated bacteria, and plant pathogens under in vitro conditions [J]. Mycorrhiza, 22(6): 437-447.
BOLIN LG, BENNING JW, MOELLER DA, 2018. Mycorrhizal interactions do not influence plant-herbivore interactions in populations of Clarkia xantiana ssp. xantiana spanning from center to margin of the geographic range [J]. Ecol Evol, 8(22): 10743-10753.
BONA E, CANTAMESSA S, MASSA N, et al., 2017. Arbuscular mycorrhizal fungi and plant growth-promoting pseudomonads improve yield, quality and nutritional value of tomato: A field study [J]. Mycorrhiza, 27(1): 1-11.
BONA E, LINGUA G, MANASSERO P, et al., 2015. AM fungi and PGP pseudomonads increase flowering, fruit production, and vitamin content in strawberry grown at low nitrogen and phosphorus levels [J]. Mycorrhiza, 25(3): 181-193.
BRYLA DR, KOIDE RT, 1990. Regulation of reproduction in wild and cultivated Lycopersicon esculentum Mill. by vesicular-arbuscular mycorrhizal infection [J]. Oecologia, 84(1): 74-81.
CAHILL JF, ELLE E, SMITH GR, et al., 2008. Disruption of a belowground mutualism alters interactions between plants and their floral visitors [J]. Ecology, 89(7): 1791-1801.
CAMPO S, MARTN-CARDOSO H, OLIV M, et al., 2020. Effect of root colonization by arbuscular mycorrhizal fungi on growth, productivity and blast resistance in rice [J]. Rice, 13: 42-53.
CASER M, VICTORINO MM, DEMASI S, et al., 2020. Arbuscular mycorrhizal fungi association promotes corm multiplication in potted saffron(Crocus sativus L.) plants [J]. Acta Hortic, 1287: 441-446.
CHEN S, JIN W, LIU A, et al., 2013. Arbuscular mycorrhizal fungi (AMF) increase growth and secondary metabolism in cucumber subjected to low temperature stress [J]. Sci Hortic, 160(3): 222-229.
CRAINE JM, WOLKOVICH EM, TOWNE EG, et al., 2012. Flowering phenology as a functional trait in a tallgrass prairie [J]. New Phytol, 193(3): 673-682.
DAVIES TJ, WOLKOVICH EM, KRAFT NJB, et al., 2013. Phylogenetic conservatism in plant phenology [J]. J Ecol, 101(6): 1520-1530.
DUFAULT RJ, PHILLIPS TL, KELLY JW, 1990. Nitrogen and potassium fertility and plant populations influence field production of Gerbera [J]. HortScience, 25(12): 1599-1602.
ELZINGA JA, ATLAN A, BIERE A, et al., 2007. Time after time: Flowering phenology and biotic interactions [J]. Trends Ecol Evol, 22(8): 432-439.
GANADE G, BROWN VK, 1997. Effects of below-ground insects, mycorrhizal fungi and soil fertility on the establishment of Vicia in grassland communities [J]. Oecologia, 109: 374-381.
GANGE AC, SMITH AK, 2005. Arbuscular mycorrhizal fungi influence visitation rates of pollinating insects [J]. Ecol Entomol, 30: 600-606.
GAUR A, GAUR A, ADHOLEYA A, 2000. Growth and flowering in Petunia hybrida, Callistephus chinensis and Impatiens balsamina inoculated with mixed AM inocula or chemical fertilizers in a soil of low P fertility [J]. Sci Hortic, 84: 151-162.
GEHRELS N, CHINCARINI G, GIOMMI P, et al., 2004. Erratum: The swift gamma-ray burst mission [J]. Astrophys J, 611(2): 1005-1020.
GIULIA C, ANNA B, CORRADO L, et al., 2015. Influence of biochar, mycorrhizal inoculation, and fertilizer rate on growth and flowering of Pelargonium (Pelargonium zonale L.) plants [J]. Front plant sci, 6: 429.
GU L, ZHANG DX, 2009. A review on ornithophily in the chinese flora [J]. J Trop Subtrop Bot, 17(2): 194-204. [顧垒, 张奠湘, 2009. 中国植物区系的鸟类传粉现象 [J]. 热带亚热带植物学报, 17(2): 194-204.]
GUO W, DENG W, YAN XF, et al., 2010. Research advances on impact factors of plant reproductive allocation [J]. J NE Agric Univ, 41(9): 150-155. [郭伟, 邓巍, 燕雪飞, 等, 2010. 植物生殖分配影响因素的研究进展 [J]. 东北农业大学学报, 41(9): 150-155.]
HE JM, ZHANG C, DAI HL, et al., 2019. A LysM receptor heteromer mediates perception of arbuscular mycorrhizal symbiotic signal in rice [J]. Mol Plant, 12(12): 1561-1576.
HEITOR LC, FREITAS MS, BRITO VN, et al., 2016. Growth and yield of marigold flowers in response to mycorrhizal inoculation and phosphorus [J]. Hortic Bras, 34: 26-30.
HOEKSEMA JD, CHAUDHARY VB, GEHRING CA, et al., 2010. A meta-analysis of context-dependency in plant response to inoculation with mycorrhizal fungi [J]. Ecol Lett, 13(3): 394-407.
HU WW, 2015. Ecological effects of arbuscular mycorrhizal fungi (AMF) on common compositae invasive species in Guangzhou [D]. Guangzhou: Sun Yat-sen University. [胡文武, 2015. 叢枝菌根真菌(AMF)对广州常见菊科入侵植物的生态学效应研究 [D]. 广州: 中山大学.]
HUANG YF, WU QL, WAN Q, et al., 2019. Research progress of arbuscular mycorrhizal fungi [J]. Mod Agric, 2019, (12): 9-12. [黄艳飞, 吴庆丽, 万群, 等, 2019. 丛枝菌根真菌的研究进展 [J]. 现代农业, (12): 9-12.]
IGIEHON NO, BABALOLA OO, 2021. Effects of rhizobia and arbuscular mycorrhizal fungi on yield, size distribution and fatty acid of soybean seeds grown under drought stress [J]. Microbiol Res, 242: 126640.
JACOBSEN DJ, RAGUSO RA, 2018. Lingering effects of herbivory and plant defenses on pollinators [J]. Curr Biol, 28(19): 1164-1169.
JIANG YN, WANG WX, XIE QJ, et al., 2017. Plants transfer lipids to sustain colonization by mutualistic mycorrhizal and parasitic fungi [J]. Science, 356: 1172-1175.
JIMENEZ-LEYVA JA, GUTIERREZ A, OROZCO JA, et al., 2017. Phenological and ecophysiological responses of Capsicum annuum var. glabriusculum to native arbuscular mycorrhizal fungi and phosphorus availability [J]. Environ Exp Bot, 138: 193-202.
JIN ZX, LI JM, LI YL, 2015. Interactive effects of arbuscular mycorrhizal fungi and copper stress on flowering phenology and reproduction of Elsholtzia splendens [J]. PLoS ONE, 10: 0145793.
KENNY H, KAMAL PA, BENTE JG, et al., 2020. Earlier onset of flowering and increased reproductive allocation of an annual invasive plant in the north of its novel range [J]. Ann Bot-London, 126(6): 1005-1016.
KIERS ET, ADLER LS, GRMAN EL, et al., 2010. Manipulating the jasmonate response: How do methyl jasmonate additions mediate characteristics of aboveground and belowground mutualisms? [J]. Funct Ecol, 24: 434-443.
KOIDE RT, DICKIE IA, 2002.Effects of mycorrhizal fungi on plant populations [J]. Plant Soil, 244: 307-317.
LAU TC, LU X, KOIDE RT, et al., 1995. Effects of soil fertility and mycorrhizal infection on pollen production and pollen grain size of Cucurbita pepo (Cucurbitaceae) [J]. Plant Cell Environ, 18: 169-177.
LAZZARA S, MILITELLO M, CARRUBBA A, et al., 2017. Arbuscular mycorrhizal fungi altered the hypericin, pseudohypericin, and hyperforin content in flowers of Hypericum perforatum grown under contrasting P availability in a highly organic substrate [J]. Mycorrhiza, 27: 345-354.
LEI X, QU WL, SHA YC, et al., 2019. Primary study on floral syndrome and breeding system of Tamarindus indica L. [J]. Chin Trop Agric, 4: 64-68. [雷虓, 瞿文林, 沙毓滄, 等, 2019. 罗望子花部综合特征和繁育系统初步研究 [J]. 中国热带农业, 4: 64-68.]
LEWIS JD, KOIDE RT, 1990. Phosphorus supply, mycorrhizal infection and plant offspring vigor [J]. Funct Ecol, 4(5): 695-702.
LI GY, ZHANG ZQ, 2019. Development, lifespan and reproduction of spider mites exposed to predator-induced stress across generations [J]. Biogerontology, 20: 871-882.
LI MR, ZHANG L, 2019. Reproductive phenological characteristics and impact factors of Macrosolen cochinchinensis in Xishuangbanna [J]. Guihaia, 39(9): 1252-1260. [李慢如, 张玲, 2019. 西双版纳地区鞘花的繁殖物候特征及影响因素 [J]. 广西植物, 39(9): 1252-1260.]
LIU JJ, LIU JL, LIU JH, et al., 2019. The potassium transporter SlHAK10 is involved in mycorrhizal potassium uptake [J]. Plant physiol, 180(1): 465-479.
LIU SJ, GUO HL, XU J, et al., 2018. Arbuscular mycorrhizal fungi differ in affecting the flowering of a host plant under two soil phosphorus conditions [J]. J Plant Ecol, 11: 623-631.
LU XH, KOIDE RT, 1994. The effects of mycorrhizal infection on components of plant growth and reproduction [J]. New Phytol, 128(2): 211-218.
LUCIANO A, ALESSANDRA T, MANUELA G, et al., 2018. Designing the ideotype mycorrhizal symbionts for the production of healthy food [J]. Front Plant Sci, 9: 1089.
MERLIN E, MELATO E, LOURENO E, et al., 2020. Inoculation of arbuscular mycorrhizal fungi and phosphorus addition increase coarse mint (Plectranthus amboinicus Lour.) plant growth and essential oil content [J]. Rhizosphere, 15: 100217.
MICHENEAU C, 2006. Bird pollination in an angraecoid orchid on Reunion island (Mascarene Archipelago, Indian Ocean) [J]. Ann Bot-London, 97(6): 965-974.
MIRANDA H, EHRET DL, KRUMBEIN A, et al., 2015. Inoculation with arbuscular mycorrhizal fungi improves the nutritional value of tomatoes [J]. Mycorrhiza, 25: 359-376.
NICHOLAS B, NICOLE G, 2015. How do belowground organisms influence plant-pollinator interactions? [J]. J Plant Ecol, 8(1): 1-11.
NURBAITY A, HAMDANI JS, RAHAYU RP, 2019. Effect of arbuscular mycorrhizal fungi and different composition of growing medium on growth and production of potato seed cultivars Medians in Inceptisols Jatinangor [J]. Earth Environ Sci, 393: 012052.
OEHL F, DA SILVA GA, GOTO BT, et al., 2011. Glomeromycota: Three new genera and glomoid species reorganized [J]. Mycotaxon, 116(1): 75-120.
OTAROLA MF, SAZIMA M, SOLFERINI VN, 2013. Tree size and its relationship with flowering phenology and reproductive output in wild nutmeg trees [J]. Ecol Evol, 3(10): 3536-3544.
PENDLETON RL, 2000. Pre-inoculation by an arbuscular mycorrhizal fungus enhances male reproductive output of Cucurbita foetidissima [J]. Int J Plant Sci, 161: 683-689.
PEREYRA M, GRILLI G, GALETTO L, 2019. Root-associated fungi increase male fitness, while high simulated herbivory decreases indirect defenses in Croton lachnostachyus plants [J]. Plant Ecol, 220: 29-39.
PHILIP LJ, POSLUSZNY U, KLIRONOMOS JN, 2001. The influence of mycorrhizal colonization on the vegetative growth and sexual reproductive potential of Lythrum salicaria L. [J]. Can J Bot, 79: 381-388.
PSCHEL D, RYDLOV J, VOSTKA M, 2014. Can mycorrhizal inoculation stimulate the growth and flowering of peat-grown ornamental plants under standard or reduced watering? [J]. Appl Soil Ecol, 80: 93-99.
RASMANN S, BENNETT AE, BIERE A, et al, 2017. Root symbionts: Powerful drivers of plant above- and belowground indirect defences [J]. Insect Sci, 24: 947-960.
REN MX, ZHANG QG, ZHANG DY, 2004. Geographical variation in the breeding systems of an invasive plant, Eichhornia crassipes, within China [J]. Chin J Plant Ecol, 28(6): 753-760. [任明迅, 張全国, 张大勇, 2004. 入侵植物凤眼蓝繁育系统在中国境内的地理变异 [J]. 植物生态学报, 28(6): 753-760.]
SABIA E, CLAPS S, MORONE G, et al., 2015. Field inoculation of arbuscular mycorrhiza on maize (Zea mays L.) under low inputs: Preliminary study on quantitative and qualitative aspects [J]. Ital J Agron, 10: 30-33.
SAINI I, AGGARWAL A, KAUSHIK P, 2019. Inoculation with mycorrhizal fungi and other microbes to improve the morpho-physiological and floral traits of Gazania rigens (L.) Gaertn [J]. Agriculture, 9(3): 51.
SCAGEL CF, 2004a. Inoculation with vesicular-arbuscular mycorrhizal fungi and rhizobacteria alters nutrient allocation and flowering of harlequin flower [J]. HortTechnology, 14: 39-48.
SCAGEL CF, 2004b. Soil pasteurization and mycorrhizal inoculation alter flower production and corm composition of Brodiaea laxa “queen Fabiola” [J]. HortScience, 39: 1432-1437.
SCAGEL CF, SCHREINER RP, 2006. Phosphorus supply alters tuber composition, flower production, and mycorrhizal responsiveness of container-grown hybrid Zantedeschia [J]. Plant Soil, 283: 323-337.
SCHUBERT R, WERNER S, CIRKA H, et al., 2020. Effects of arbuscular mycorrhization on fruit quality in industrialized tomato production [J]. Int J Mol Sci, 21(19): 7029.
SMITH SE, SMITH FA, 2011. Roles of arbuscular mycorrhizas in plant nutrition and growth: New paradigms from cellular to ecosystem scales [J]. Ann Rev Plant Biol, 62(1): 227-250.
SONG JL, LI S, LIN Y, et al., 2019. Effects of arbuscular mycorrhizal fungi on the growth and reproduction of Ageratina adenophora [J]. J Biosafety, 28(1): 44-48. [宋潔蕾, 李胜, 林莹, 等, 2019. 丛枝菌根真菌对紫茎泽兰生长与繁殖的影响 [J]. 生物安全学报, 28(1): 44-48.]
SUN SG, LU B, LU XM, et al., 2018. On reproductive strategies of invasive plants and their impacts on native plants [J]. Biodivers Sci, 26(5): 457-467. [孙士国, 卢斌, 卢新民, 等, 2018. 入侵植物的繁殖策略以及对本土植物繁殖的影响 [J]. 生物多样性, 26(5): 457-467.]
TANG Y, 2020. Impact of grazing intensities on reproduction patterns of elm trees (Ulmus pumila) in degraded sandy lands in China [J]. PeerJ, 8(4): 9013.
TAO ZX, GE QS, XU YJ, et al., 2020. Comparison of changes in flowering phenology of woody plants and temperature sensitivity between Xi’an and Baoji [J]. Acta Ecol Sin, 40(11): 3666-3676. [陶泽兴, 葛全胜, 徐韵佳, 等, 2020. 西安和宝鸡木本植物花期物候变化及温度敏感度对比 [J]. 生态学报, 40(11): 3666-3676.]
TIAN DS, PAN QM, SIMMONS M, et al., 2012. Hierarchical reproductive allocation and allometry within a perennial bunchgrass after 11 years of nutrient addition [J]. PLoS ONE, 7(9): 42833.
VAINGANKAR JD, RODRIGUES BF, 2015. Effect of arbuscular mycorrhizal (AM) inoculation on growth and flowering in Crossandra infundibuliformis (L.) Nees [J]. J Plant Nutr, 38: 1478-1488.
VARGA S, KYTVIITA MM, 2010. Gender dimorphism and mycorrhizal symbiosis affect floral visitors and reproductive output in Geranium sylvaticum [J]. Funct Ecol, 24: 750-758.
WANG AZ, WANG QJ, LI CH, et al., 2020. Study on pollination biology of Sorbus pohuashanensis [J]. Nor Horticul, 2: 73-79. [王爱芝, 王清君, 李长海, 等, 2020. 花楸的传粉生物学研究 [J]. 北方园艺, 2: 73-79.]
WANG HJ, GE QS, DAI JH, et al., 2015. Geographical pattern in first bloom variability and its relation to temperature sensitivity in the USA and China [J]. Int J Biometeorol, 59(8): 961-969.
WASER NM, PRICE MV, 1981. Effects of grazing on diversity of annual plants in the Sonoran Desert [J]. Oecologia, 50: 407-411.
WESTON LA, MATHESIUS U, 2013. Flavonoids: Their structure, biosynthesis and role in the rhizosphere, including allelopathy [J]. J Chem Ecol, 39: 283-297.
WRIGHT JW, MEAGHER TR, 2003. Pollination and seed predation drive flowering phenology in Silene latifolia (Caryophyllaceae) [J]. Ecology, 84(8): 2062-2073.
XIE MM, WU QS, 2018. Arbuscular mycorrhizal fungi regulate flowering of Hyacinths orientalis I. Anna marie [J]. Emir J Food Agric, 30: 144-149.
XING JF, HU XY, ZHANG XB, 2015. Research on floral syndrome of Cassia surattensisin Haikou [J]. Chin Hortic Abstr, 5: 10-11. [邢剑锋, 胡翔宇, 张玄兵, 2015. 海口黄槐决明花部综合特征研究 [J]. 中国园艺文摘, 5: 10-11.]
YANG X, YU HQ, ZHANG T, et al., 2016.Arbuscular mycorrhizal fungi improve the antioxidative response and the seed production of suaedoideae species Suaeda physophora Pall under salt stress [J]. Not Bot Hortic Agrobo, 44(2): 533-540.
YU T, ELKE GN, BENARD N, et al., 2013. Effects of single and mixed inoculation with two arbuscular mycorrhizal fungi in two different levels of phosphorus supply on β-carotene concentrations in sweet potato(Ipomoea batatas L.) tubers [J]. Plant Soil, 372: 361-374.
ZHANG C, AN YM, JASCHKE Y, et al., 2020. Processes on reproductive ecology of plant species in the Qinghai-Xizang Plateau and adjacent highlands [J]. Chin J Plant Ecol, 44: 1-12. [張婵, 安宇梦, Jschke Y, 等, 2020. 青藏高原及周边高山地区的植物繁殖生态学研究进展 [J]. 植物生态学报, 44: 1-21.]
ZHANG C, CHA SQ, YANG YP, et al., 2012. Effects of the yellow barbs of the staminodes on reproductive success of Delphinium caeruleum (Ranunculaceae) [J]. Biodivers Sci, 20(3): 348-353. [张婵, 查绍琴, 杨永平, 等, 2012. 蓝翠雀花退化雄蕊上的黄色髯毛对其繁殖成功的影响 [J]. 生物多样性, 20(3): 348-353.]
ZHAO FG, DONG ZH, CHE YM, et al., 2019.AM fungi Glomous mosseae promote tobacco(Nicotiana tabacum) growth by regulating IAA metabolism [J]. J Agric Biotechnol, 27(1): 63-70. [赵方贵, 董志昊, 车永梅, 等, 2019. AMF摩西球囊霉调控IAA代谢促进烟草生长 [J]. 农业生物技术学报, 27(1): 63-70.]
ZHANG L, FAN JQ, DING XD, et al., 2014. Hyphosphere interactions between an arbuscular mycorrhizal fungus and a phosphate solubilizing bacterium promote phytate mineralization in soil [J]. Soil Biol Biochem, 74: 177-183.
ZHANG RQ, ZHU HH, ZHAO HQ, et al., 2013. Arbuscular mycorrhizal fungal inoculation increases phenolic synthesis in clover roots viahydrogen peroxide, salicylic acid and nitric oxide signaling pathways [J]. J Plant Physiol, 170(1): 74-79.
ZHANG T, YANG P, GUO YW, et al., 2018. Variation of floral scent composition of Amomum villosum before and after pollination and its ecological significance [J]. Chinese J Ecol, 37(12): 3640-3646. [张亭, 杨培, 郭岩伟, 等, 2018. 阳春砂仁(Amomum villosum)花气味传粉前后的变化及其生态意义 [J]. 生态学杂志, 37(12): 3640-3646.]
ZHANG YX, BI YL, GUO N, et al., 2019. Effects of arbuscular mycorrhizal fungi on the growth of Medicago falcate [J]. J Chin Coal Soc, 44(12): 3815-3822. [張延旭, 毕银丽, 郭楠, 等, 2019. 接种不同丛枝菌根真菌对黄花苜蓿生长影响 [J]. 煤炭学报, 44(12): 3815-3822.]
ZHU Y, LIU YX, GONG XF, et al., 2016. Symbiosis between three arbuscular mycorrhizal fungi and three host plants [J]. Microbiol Chin, 43(3): 527-533. [祝英, 刘英霞, 巩晓芳, 等, 2016. 3种丛枝菌根真菌与3种寄主植物的共生关系 [J]. 微生物学通报, 43(3): 527-533.]
(责任编辑 何永艳)
猜你喜欢
科学之友(2021年12期)2021-12-23
东坡赤壁诗词(2021年4期)2021-09-05
VOGUE服饰与美容(2020年1期)2020-04-17
东坡赤壁诗词(2018年6期)2018-12-22
科教导刊·电子版(2018年6期)2018-06-06
大科技·百科新说(2018年3期)2018-04-11
科学种养(2017年11期)2017-11-28
滇池(2017年6期)2017-06-14
中国绿色画报(2016年9期)2017-05-31
诗选刊(2016年4期)2016-10-21
