
不同遗传背景籼稻白叶枯病抗性基因Xa21、Xa23品系的抗性评价
2021-01-13 05:05于江辉刘廷昌翁绿水李锦江邓力华
热带作物学报 2021年12期
关键词:水稻
于江辉 刘廷昌 翁绿水 李锦江 邓力华


摘 要:以含有水稻白葉枯病抗性基因Xa21和Xa23的材料为供体,以课题组选育不含Xa21或Xa23基因的材料为受体,通过杂交、复交等方法,并采用分子标记辅助选择(molecular marker-assisted selection,MAS)世代选择,将含有Xa21和Xa23基因的材料、含与不含Xa21或Xa23基因的姊妹系接种7种广泛使用的白叶枯病菌种生理小种,分析了这些材料白叶枯病的抗性。研究表明含有抗性基因Xa21的水稻材料抗性累计(HR、R、MR)所占比例依次为菌株HNA1-4、PXO86(84.62%)>YN24(82.14%)>GD1358(73.08)>PXO99(67.86%)>GDA2(63.33%)>FuJ(6.67%),表明含Xa21基因材料对菌株FuJ所诱发的白叶枯病基本无抗病能力,但对于其他菌株,仍有36.67%~15.38%的材料易感病;含Xa23基因的材料在所有诱发菌株中抗性累计(HR、R、MR)均在76%以上,仍有23.02%~15.52%的材料对7种诱发菌株没有抗性。通过对菌斑长度变异系数大小的分析,表明含有Xa21或Xa23基因的材料对不同菌株抗病能力或对同一菌株的抗病能力不同材料有较大的变化。通过对诱发白叶枯病菌株间菌斑长度的相关性分析,表明含有Xa21的材料菌株GDA2或HNA1-4与对应的其他6菌株之间达到了显著或极显著正相关,含有Xa23基因的材料菌株YN24、GD1358、PXO86与对应的其他6种菌株间呈极显著正相关,含此2类基因材料抗性鉴定选取对应菌系可以提高选育效率。通过对姊妹系的分析表明,Xa21或Xa23基因的白叶枯病抗性与选育材料的背景有关。相关性分析表明,Xa21基因的材料抗性与亲本材料呈极显著正相关(R=0.5725**),Xa23基因材料抗性与亲本材料呈显著正相关(R=0.2212*)。
关键词:水稻;白叶枯病;抗性分析;抗性基因
中图分类号:S184 文献标识码:A
Abstract: The rice varieties of bacterial leaf blight resistance genes Xa21 or Xa23 were used as the donors, and the va-rieties without the genes as the receptor. By back crossing, composite crossing and other methods, combined with molecular marker-assisted selection (MAS), the varieties of Xa21 or Xa23, sister lines with or without Xa21 or Xa23 gene were inoculated with 7 widely used bacterial leaf blight pathogens, and the resistance of the varieties to the bac-terial leaf blight was analyzed. The cumulative proportion of resistance (HR, R, MR) of the Xa21varieties was HNA1-4/PXO86 (84.62%)>YN24 (82.14%)>GD1358 (73.08)>PXO99 (67.86%)>GDA2 (63.33%)>FuJ (6.67%), and the varieties of Xa21 gene had no resistance to the bacterial leaf blight induced by FuJ, but for other pathogens, there were still 36.67%~15.38% of the varieties susceptible to diseases. The cumulative resistance (HR, R, MR) of the varieties of the Xa23 was over 76%, but there were still 23.02%~15.52% of the varieties that had no resistance to the 7 pathogens. Lesion length analysis, showed that the varieties of the Xa21 or Xa23 were resistant to different pathogen or the same pathogen. Different varieties had much variance. Lesion length analysis of of the7 bacterial leaf blight pathogens among the varieties of Xa21and Xa23 showed significant or extremely significant difference to GDA2 or HNA1-4 and other pathogens, and there was a extremely significant difference of YN24, GD1358 or PXO86 and other pathogens. The sister lines showed that bacterial leaf blight resistance of Xa21 or Xa23 was related to the donor of the varieties. Correlation of the varieties of Xa21 and parents showed extremely significant difference (R=0.5725**), and there was a significant difference correlations of the varieties of Xa23 and parents (R=0.2212*).
Keywords: rice; bacterial leaf blight; resistance analysis; resistance gene
DOI: 10.3969/j.issn.1000-2561.2021.12.008
白叶枯病是一种全球性的细菌病害,与稻瘟病、纹枯病被称为水稻的“三大病害”,其发生范围广、流行速度快、危害大、突变性高,导致水稻生产造成了巨大的损失,受害田块一般减产10%~20%,重者减产50%以上,甚至绝收[1-4]。使用化学农药或种植抗病品种能有效控制白叶枯病的发生。研究表明水稻白叶枯病是由稻黄单胞菌致病变种引起的,是一种维管束疾病,即水稻维管感染此病菌产生灰白色的病变,糖转运蛋白将蔗糖转运到细胞外部空间,导致水稻白叶枯病菌在维管束中大量繁殖,从而堵塞维管的养分运输,同时,高水平的蔗糖释放会干扰到细胞膜的功能,扰乱糖代谢信号途径,使用农药并不能直接接触病灶,致使农药效果不佳,导致严重的环境污染,而且破坏生态平衡、增加种植成本。利用抗病基因培育抗病品种对防治水稻白叶枯病最为经济有效,同时可以减少污染环境[5-6]。迄今为止,已有44个抗白叶枯病基因(显性基因27个,隐性基因17个)被研究报道[7-8],其中Xa1、Xa3/Xa26、xa5、Xa10、xa13、Xa21、Xa23、xa25和Xa27等9个基因已被克隆,为培育抗病品种提供了极好的遗传资源[5, 9]。在雜交稻抗病改良上应用比较多的基因有Xa4、Xa21、Xa23[10]。研究发现来源于长药野生稻(Oryza longistaminata)的Xa21和来源于普通野生稻(O. rufipogon)的Xa23广谱高抗显性基因在实际育种中利用更广泛[11-12]。利用常规育种与分子标记辅助选择(molecular marker-assisted selection,MAS)的方法,将白叶枯病抗性基因Xa21和Xa23导入具有高产优质但抗病能力弱的恢复系或不育系中,配制出高抗高产优质组合,已成为近年来抗病育种的重要且有效途径[13-22]。
湖南省自2007年开始水稻的白叶枯病抗性鉴定以来,审定品种的抗性评价基本保持中抗水平,但2018年审定品种的平均白叶枯病抗性评价等级为中感(5.6级),白叶枯病抗性育种水平有所下降[23]。笔者根据湖南省近2年来水稻区试或审定情况统计,2019年参加湖南省区试的120个水稻组合(品种)中,白叶枯病抗病组合(3.0级)仅有1个,34个组合抗性为7.0级(感),3个组合抗性为9.0级(高感),感和高感组合占到30.8%,其余均为5.0级(中感);2020年参加湖南省区试的136个水稻品种中,白叶枯病抗性在7级以上的有75个,占半数之多,剩下的材料大多抗性为5.0级;2020年湖南省审定水稻品种103个中,仅有2个组合白叶枯病抗性为3.0级(抗病),其余为5.0级或以上。可见,水稻白叶枯病抗性品种的匮乏,白叶枯病的抗性育种迫在眉睫。同时,由于抗性基因与白叶枯病菌一直在不断地进化,任何一方发生变化均可能导致具有抗病的品种失效。为此,不断发掘或培育新的抗病材料对白叶枯病抗性育种具有重大意义,而且培育对多种白叶枯病生理小种抗性材料可以加宽抗谱和加长抗性时间。本研究将含有水稻白叶枯病抗性基因Xa21和Xa23的材料为供体,以课题组多年来选育不含Xa21或Xa23基因的材料为受体,通过杂交、回交、复交等方法利用MAS通过前景和背景选育,目的在于提高这些材料的白叶枯病抗性或培育抗谱较宽及抗性持久的水稻品系。将育成的高世代稳定的品系接种7种广泛应用的白叶枯病菌种生理小种,分析了含Xa21或Xa23基因的水稻材料白叶枯病的抗性,通过田间鉴定评价了不同水稻品系的白叶枯病抗性,筛选出抗性得到改良并且综合性状优良的株系,为白叶枯病抗性育种及防治水稻白叶枯病等工作提供种子资源,同时评价了Xa21或Xa23基因材料对各生理小种抗病能力的强弱,抗性基因与各生理小种之间关联性的大小,以及含Xa21或Xa23基因水稻材料与不含基因姊妹系抗白叶枯病的相关能力大小,旨在为白叶枯病分子抗性育种提供参考。
1 材料与方法
1.1 材料
供试材料为中国科学院亚热带农业生态研究所水稻耐逆境分子育种课题组选育的含有Xa21基因的材料112份和含有Xa23基因的材料117份,含有与不含Xa21基因的姊妹系12份,含有与不含Xa23基因的姊妹系26份。这些材料是多年来课题组通过杂交、复交和MAS在F6代以上已经稳定的材料。
试验于2019年在中国科学院亚热带农业生态研究所长沙水稻试验站(l13º9 E,28º9 N)进行,该区域年平均气温17.2 ℃,积温5457 ℃,属亚热带季风湿润气候,气候温和,降水充沛,四季分明。
1.2 方法
试验采用人工插秧栽培,5月25日播种,6月20日移栽,株行距为16.5 cm×26.4 cm,每穴栽植1粒谷苗,每材料50兜。按常规栽培方式进行田间管理。插秧后2周取供试材料的单株叶片,CTAB[24]法提取水稻基因组DNA。采用10 μL的PCR扩增体系对供试材料的Xa21和Xa23基因进行扩增,体系组成为2×PARMS master mix 5 μL,10 mmol/L正向标记引物0.15 μL,10 mmol/L反向标记引物0.15 μL,模板DNA 1.0 μL,ddH2O 3.7 μL。Xa21基因的选择标记是与其紧密连锁的PCR标记pTA248[25],引物序列为:5‒AGACGC GGAAGGGTGGTTCCCGGA‒3(pTA248-F),5‒ CGATCGCTATAACAGCAAAAC‒3(pTA248- R),退火温度60 ℃;Xa23基因的选择标记引物序列为:5‒TTGCTCAAGGCTAGGAAAATG‒3(M- Xa23-F),5‒CCCCATCAACGAACTACAGG‒3(M-Xa23-R),退火温度55 ℃;PCR扩增产物用1.5%琼脂糖凝胶电泳检测。
本研究所用诱发白叶枯病菌种7种:FuJ和YN24(华南稻区优势致病菌)、HNA1-4(长江中下游水稻区试白叶枯病抗性鉴定菌种)、GDA2(广东省白叶枯病致病代表菌株)、PXO86和PXO99(菲律宾白叶枯病菌生理小种)、GD1358(南方稻区白叶枯病菌优势小种),均由湖南省植保所稻瘟病鉴定中心提供[26]。试验采用剪叶接种的方法进行鉴定。首先将在‒20 ℃保存的菌种在NA培养基上活化和复壮,其次在28~30 ℃恒温再培养48 h,最后用无菌水洗脱菌株,使菌液均匀悬浮,并且将浓度调节为3×108 CFU/mL(OD600=0.5),用于做接种试验。接种时将手术剪灭菌,蘸取已制备好的白叶枯病菌悬浮液。所有材料在孕穗期接种,将供试材料叶片叶尖剪去2~3 cm,每株植株叶片接种5片,每个材料接种3株,3次重复,以感病水稻‘金刚30’作为对照,并做好标记。接种21 d左右,当参试材料的病情趋于稳定时,量取病斑长度,鉴定供试植株的抗病性。
1.3 数据处理
整理测量数据,根据白叶枯病的分级标准[23]:平均病斑长度小于1 cm为高抗(HR);1.1~3.0 cm为抗(R);3.1~5.0 cm为中抗(MR);5.1~12.0 cm为中感(MS);12.1~20.0 cm为感(S);大于21.0 cm高感(HS)。根据对照材料‘金刚30’的发病等级,分析水稻对7種白叶枯病菌的致病力,并按照抗性分级标准对供试植株进行抗感分级。利用Microsoft Excel 2018进行常规的数据统计,采用DPS 15.10软件计算均值和均值标准差,对Xa21和Xa23抗性基因在各菌株间的相关性、含有和不含抗性基因姊妹系的比较及不同材料对白叶枯病菌株抗性的相关分析进行分析。
2 结果与分析
2.1 Xa21和Xa23基因材料对诱发白叶枯病菌株的抗性分析
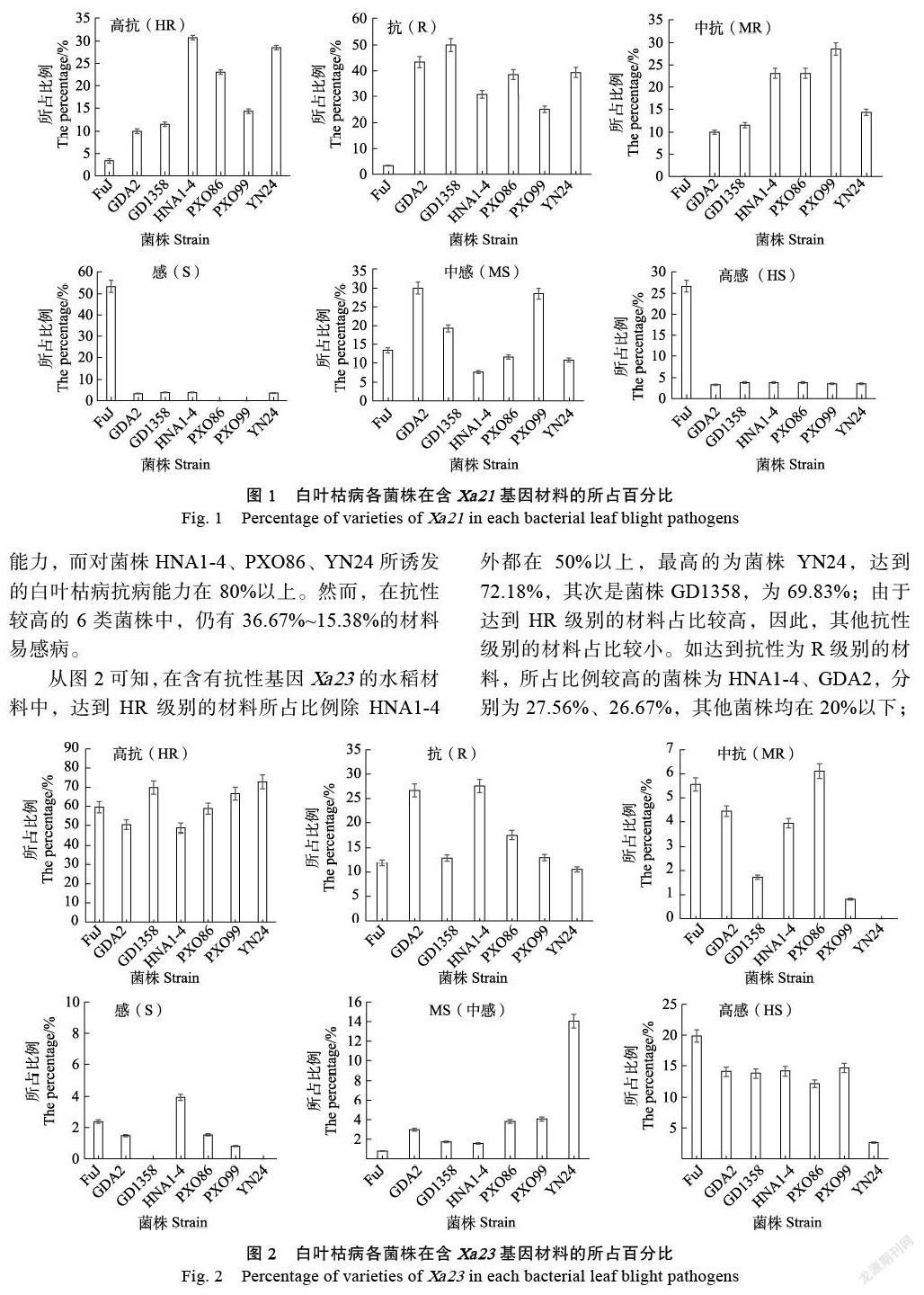
2.1.1 抗性频率分析 研究结果表明,含有抗性基因Xa21的水稻材料在诱发白叶枯病的菌株HNA1-4中达到高抗(HR)级别的材料所占比例最高,为30.77%,而菌株FuJ中高抗(HR)材料所占比例最小,仅为3.33%;菌株GD1358中抗性(R)材料所占比例最高,达到50%,占比最小的菌株是FuJ(3.33%);菌株PXO99中抗(MR)材料所占比例最高,但不到30%,而菌株FuJ中无中抗材料(图1)。在所有诱发菌株中水稻材料抗性累计(HR、R、MR)所占比例依次为菌株HNA1-4、PXO86(84.62%)>YN24(82.14%)>GD1358(73.08)>PXO99(67.86%)>GDA2(63.33%)>FuJ(6.67%),表明白叶枯病抗性基因Xa21对菌株FuJ所诱发的白叶枯病基本无抗病能力,而对菌株HNA1-4、PXO86、YN24所诱发的白叶枯病抗病能力在80%以上。然而,在抗性较高的6类菌株中,仍有36.67%~15.38%的材料易感病。
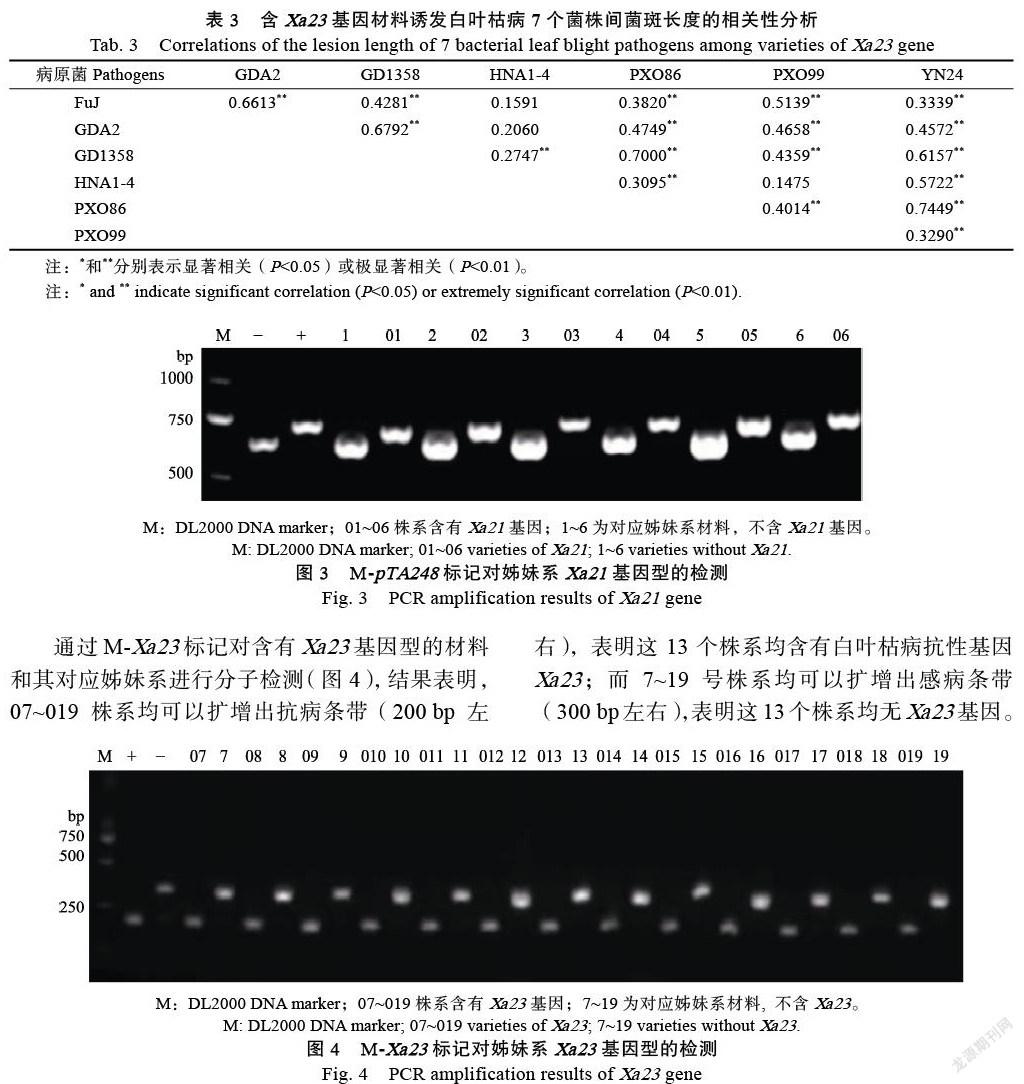
从图2可知,在含有抗性基因Xa23的水稻材料中,达到HR级别的材料所占比例除HNA1-4外都在50%以上,最高的为菌株YN24,达到72.18%,其次是菌株GD1358,为69.83%;由于达到HR级别的材料占比较高,因此,其他抗性级别的材料占比较小。如达到抗性为R级别的材料,所占比例较高的菌株为HNA1-4、GDA2,分别为27.56%、26.67%,其他菌株均在20%以下; 而达到MR级别的材料,在所有菌株中所占比例均在10%以下。但是在所有诱发菌株中水稻材料抗性累计(HR、R、MR)均在70%以上,最高的为菌株GD1358,最低的为菌株FuJ。因此,白叶枯病抗性基因Xa23的材料有76.98%~84.48%对7个诱发菌株有抗性,表明Xa23基因较Xa21基因有较好的抗白叶枯病能力,且比Xa21基因抗菌谱宽,但是有23.02%~15.52%的材料易感病。
2.1.2 病斑长度及变异系数分析 研究结果表明,含有白叶枯病抗性基因Xa21的材料对不同诱发菌株所引发的病斑长度变异大小不同,而且所有菌株变异系数较大(表1),表明材料间的抗病能力有极大不同。其中变异系数最大的为YN24菌株,其次为HNA1-4菌株,变异系数均在100%以上,变异系数最小的菌株为FuJ(41.3%)。而含有Xa23基因的材料在各诱发菌株中的变异系数较大,均在140%以上,并且PXO99、FuJ的变异系数超过了200%,相比之下,变异系数最小的菌株为PXO86。综上所述,含有Xa21或Xa23基因的材料,同一材料对不同菌株的抗病能力或不同材料对同一菌株的抗病能力有较大的变化。
2.1.3 诱发Xa21或Xa23基因材料白叶枯病7菌株间的相关性分析 由表2可知,含有Xa21基因的材料中,菌株GD1358与FuJ之间、PXO86与GD1358之间、FuJ与PXO99之间、GD1358与PXO99之间、YN24与FuJ之间菌斑长度无显著相关性,GDA2与其他6个菌株之间、HNA1-4与其他6个菌株之间均达到了显著(P<0.05)或极显著(P<0.01)正相关,表明在含有Xa21基因的材料中,只要抗GDA2、HNA1-4中任何一种菌株,就有可能抗其他菌株。
相比含有白叶枯病抗性基因Xa21的材料,含有抗性基因Xa23的材料抗病能力增强。从表3可看出,在含有Xa23基因的材料中,除菌株FuJ、GDA2与HNA1-4之间及PXO99与HNA1-4之间抗性菌斑长度无显著相关性外,其余菌株间均呈极显著正相关,而且YN24与其他6菌株间、GD1358与其他6菌株间、PXO86与其他6菌株间抗性菌斑长度呈极显著正相关,表明抗YN24、GD1358、PXO86中任何一种菌株,就有可能抗其他菌株,Xa23基因白叶枯病抗普较宽。
2.2 Xa21和Xa23基因材料与其不含基因的姊妹系抗性分析
2.2.1 分子检测结果 通过M-pTA248标记对含有Xa21基因的材料和其对应姊妹系进行分子检测,结果表明,01~06株系均含有Xa21基因抗性条带(图3),表明这6个材料均含有白叶枯病抗性基因Xa21,姊妹系1~6均含有感病条带,表明这6个材料都不含有Xa21基因。
通过M-Xa23标记对含有Xa23基因型的材料和其对应姊妹系进行分子检测(图4),结果表明,07~019株系均可以扩增出抗病条带(200 bp左右),表明这13个株系均含有白叶枯病抗性基因Xa23;而7~19号株系均可以扩增出感病条带(300 bp左右),表明这13个株系均无Xa23基因。
2.2.2 Xa21和Xa23基因材料与不含基因姊妹系的比较 从表4可看出,接种白叶枯病菌株FuJ后,含Xa21基因的材料与不含该基因的姊妹系抗性水平没有多大改变,仍为感病,再次证明Xa21基因对菌株FuJ没有抗病能力;而含Xa23基因的材料07、08、010、011、015、017、019号与不含该基因的姊妹系比较白叶枯病抗性有较大改变,从感病或高感转为了抗病或高抗,而09、012、013号抗性水平没有改变,抗性频率达到76.9%。接种白叶枯病菌株GDA2后,与对应姊妹系比较,含Xa21基因的株系抗性水平均有较大改变,其中04、05号株系从高感转变为中抗;含Xa23基因的07、08、010、012、013、015、017、019号株系白叶枯病抗性水平有较大提升,转为抗或高抗,抗性频率达到92.3%。接种白叶枯病菌株GD1358后,含Xa21基因的04、05、06号株系抗性有较大改变;含Xa23基因的08、012、015、017、019号株系抗性有较大的改变,而09、013号仍为感病,抗性频率达到84.6%。接种白叶枯病菌株HNA1-4后,含Xa21基因除02号株系外,其他株系抗性都增强了,抗性频率达到83.3%;含Xa23基因的07、08、011、014、015、019号株系抗性有极大改变,而09、010、012、013号抗性水平没有提升,但抗性频率为69.2%。接种白叶枯病菌株PXO86后,含Xa21基因除02号株系外,其他株系抗性均有增强,抗性频率达到83.3%;含Xa23基因除09號株系外,其他株系抗性水平有较大改变,抗性频率为84.6%。接种白叶枯病菌株PXO99后,含Xa21基因除02号株系外,其他抗性水平都有所增强,但抗性频率为66.7%;含Xa23基因除09、013号为感病株系外,其余均为抗性株系,抗性频率为84.6%。接种白叶枯病菌株YN24后,含Xa21基因除02号株系外,其他抗性水平为抗或高抗;含Xa23基因除09、013号为感病株系外,其他均为抗性株系,抗性频率为84.6%。
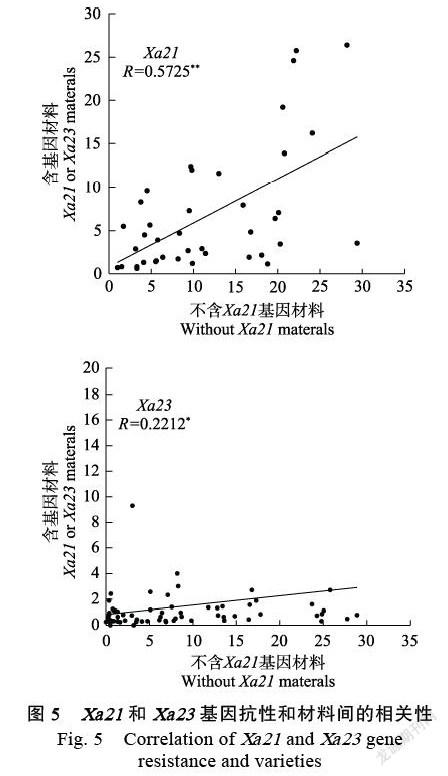
2.2.3 Xa21和Xa23基因抗性和材料间的相关性 通过对含有Xa21或Xa23基因的材料与其不含基因的姊妹系进行抗白叶枯病的相关性分析(图5),研究表明,含有Xa21基因的材料与其姊妹系之间抗性呈极显著正相关,相关系数达到0.5725;含有Xa23基因材料与其姊妹系之间抗性呈显著正相关,但相关系数不大,为0.2212。
3 讨论
本研究表明,含白叶枯病抗性基因Xa21的籼稻品系对7个白叶枯病菌种生理小种的抗性最高的为菌株HNA1-4、PXO86,最低的为菌株FuJ,而含抗性基因Xa23的籼稻品系抗性最高的为菌株GD1358,最低的为菌株FuJ,可见菌株FuJ对这2种抗性基因材料的致病力最强。吴云雨等[27]通过12个白叶枯病生理小种(包括国内的FuJ、GD414、HEN11、YN1、YN7、YN11、YN18、YN24和SYb共9个小种及菲律宾的PXO079、PXO145和PXO099 3个小种)对国内不同稻区推广应用的65个籼稻品种的致病力差异分析,表明致病力最强的是菌株FuJ小种,可以使测试的全部品种都表现出感病或中感反应,本研究与其相印证;但杨军等[18]通过对64个粳稻品种对10个白叶枯病菌种(YN18、YN1、GD414、HEN11、SYb、YN7、YN11、FuJ、YN24和菲律宾菌株PXO99)的抗性分析,结果表明致病力最强的菌株为YN24、PXO99和YN1,而菌株FuJ的致病力较弱,与本研究不同。究其原因可能是病原菌FuJ对籼稻品种致病能力更强,而病原菌YN24对粳稻品种致病能力更强。在进一步的研究中还发现含Xa21基因的水稻品系对菌株FuJ所诱发的白叶枯病基本无抗病能力,同时对其他6个生理小种仍有36.67%~15.38%的材料易感病;而含Xa23基因的材料对7个诱发菌株感病材料为23.02%~15.52%。表明Xa23基因对白叶枯病的抗病能力强于Xa21基因。Xa21基因虽然抗性强,但是对中国浙江、云南等一些白叶枯病小种没有抗性[13-14],对韩国96%的菌株没有抗性[15]。而Xa23基因由于具有抗谱广、抗性强、完全显性和全生育期抗病等特点,备受国内外育种家的关注,相继被用于开展分子标记的开发与育种利用研究[16-18]。本研究所选7种白叶枯病菌种生理小种,在南方稻区白叶枯病抗性中广泛应用,具有很高的代表性[26],通过对诱发Xa21或Xa23基因材料白叶枯病的7种菌株间的相关性分析表明,在含Xa21基因的材料中,GDA2、HNA1-4与对应的其他6个菌株之间均达到了显著或极显著正相关,只要抗这2种菌株中任何一种菌株,就有可能抗其他菌株;在含Xa23基因的材料中,YN24、GD1358、PXO86与对应其他6种菌株间呈差异极显著正相关,只要抗这3种菌株的任何一种菌株,就有可能抗其他菌株。可能是由于材料所含抗性基因不同对白叶枯病菌株的抗感性有较大差异,因此研究结果不一致,但前人对此类研究未见报道。
周元飞等[25]利用杂交、回交和自交选育的方法,通过MAS选育,将抗白叶枯病基因Xa21转到籼稻恢复系‘9311’中,利用菲律宾6号小种PXO99接种鉴定,表明后代的恢复系及相应杂交稻组合对白叶枯病抗性显著增强;已有研究表明,利用国内主要白叶枯病菌系Ⅲ、Ⅳ、Ⅴ、Ⅵ对选育的含Xa21基因的恢复系‘R8006’‘R1176’进行剪叶接种,均显示出良好的抗性[28]。而本研究对含有Xa21基因材料112份以及含有或不含Xa21基因的姊妹系白叶枯病抗性分析表明,导入Xa21后,并非对所有菌株抗性均增强,其中对菌株FuJ没有抗病能力。兰艳荣等[19]通过1次杂交、3次回交和3次以上自交得到的含Xa21基因株系‘YR7009’和‘YR7014’对白叶枯病4个菌株PXO61、PXO99、ZHE173和GD1358具有良好的抗性,而对FuJ和YN24不具有抗性;罗彦长等[20]将Xa21导入光敏核不育系‘3418S’中,在分蘖期用13(我国7个病原型和6个菲律宾国际鉴别小种)个代表菌系接种,结果表明‘3418S’(Xa21)对供试的菌系多数抗病,但也有少数表现中度感病。可见,对Xa21基因水稻材料的白叶枯病的抗感性,不同材料或不同生理小种的研究结果不尽相同,究其原因,和选育材料的背景来源有较大的关系,同时,其他基因对其表达可能有拮抗作用,就其机理还需进一步剖析。结果表明,对于含有Xa23基因的品系受遗传背景的影响更大,有7对姊妹系抗性均增强,1对姊妹系(09和9号)抗性没有改变,5对姊妹系抗性菌株不同抗性变化不一。可见,在本研究中水稻品系导入Xa23基因后,亦不能使所有品系抗性都增强。前人的研究采用单菌系接种结果也说明了含Xa23基因的水稻材料并不能完全抗病,杨德卫等[29]通过白叶枯病强毒菌系P6对8份含Xa23基因的姊妹系进行田间接种鉴定,结果表明7份表现抗病,1份表现感病;于洁等[30]对含Xa23基因的351份近等位基因系通过菌系P6进行白叶枯病的田间接种鉴定,筛选到抗性材料74份,其中高抗材料54份;范宏环等[31]通过杂交和MAS技术,在‘18113/H705F3’和‘18113/H706F3’株系中,分别获得71和52份携有Xa23纯合基因型恢复系,采用水稻白叶枯病Ⅳ型小种代表菌株浙173进行剪叶接种,但分别鉴定出61和44份抗病株系。李进波等[32]从160份背景来源相同且Xa23基因纯合株系中选取21份使用鉴别菌系P6进行了抗性鉴定,表明所选株系在苗期和孕穗期都表现抗病。虽然不同的研究结果有异,但利用与抗病基因紧密连锁的分子標记辅助选择选育材料,能够准确、快速地进行抗性基因鉴定或抗源筛选,在后代群体里可以筛选出农艺性状优良且抗性增强的材料,进而提高育种效率,而且多代回交可以加快材料稳定缩短育种周期。
4 结论
通过对含有白叶枯病抗性基因Xa21或Xa23的材料对7种白叶枯病菌株的抗性分析,结果表明含Xa21基因的材料在抗性较高的6种菌株中(除FuJ菌株),抗性频率在60%以上,含Xa23基因的材料对7种菌株的抗性频率在76%以上,表明Xa23基因比Xa21基因抗性效果强。相关分析表明,含有Xa21的材料中,菌株GDA2或HNA1-4与对应的其他6种菌株之间达到了显著或极显著正相关,含有Xa23基因的材料中,菌株YN24、GD1358、PXO86与对应的其他6种菌株间呈极显著正相关,选取此类菌系进行鉴定可以提高选育效率。通过对含有或不含基因的姊妹系分析,表明Xa21基因材料和Xa23基因材料抗性分别与亲本材料呈极显著或显著正相关,说明后代材料的抗性和亲本的遗传背景有极大的关系。
参考文献
[1] Mew T W. Current status and future prospects of research on bacterial blight of rice[J]. Annual Review of Phytopathology, 1987, 25(1): 359-382.
[2] 钱 君, 张世珖, 姬广海. 云南稻种对水稻白叶枯病抗性的评价[J]. 云南农业大学学报, 2003, 18(3): 239-245, 252.
[3] 章 琦. 中国杂交水稻白叶枯病抗性的遗传改良[J]. 中国水稻科学, 2009, 23(2): 111-119.
[4] Niño-Liu D O, Ronald P C, Bogdanove A J. Xanthomonas oryzae pathovars: model pathogens of a model crop[J]. Mo-lecular plant pathology, 2006, 7(5): 303-324.
[5] 陈析丰, 梅 乐, 冀占东, 等. 中国稻种资源中新抗白叶枯病基因的发掘[J]. 浙江师范大学学报(自然科学版), 2020, 43(1): 8-12.
[6] 向 贤, 陈露露, 张丹丹, 等. 水稻白叶枯病抗性基因物理图谱定位与功能标记[J]. 分子植物育种, 2019, 17(2): 509-516.
[7] Liang L Q, Wang C Y, Zeng L X, et al. The rice cultivar Baixiangzhan harbours a recessive gene xa42(t) determining resistance against Xanthomonas oryzae pv. oryzae[J]. Plant Breeding, 2017, 136(5): 603-609.
[8] Kim S M. Identification of novel recessive gene xa44(t) conferring resistance to bacterial blight races in rice by QTL linkage analysis using an SNP chip[J]. Theoretical and Ap-plied Genetics, 2018, 131(12): 2733-2743.
[9] Hutin M, Sabot F, Ghesquière A, et al. A knowledge-based molecular screen uncovers a broad-spectrum OsSWEET 14 resistance allele to bacterial blight from wild rice[J]. The Plant Journal, 2015, 84(4): 694-703.
[10] Balachiranjeevi C H, Bhaskar Naik S, Abhilash Kumar V, et al. Marker-assisted pyramiding of two major, broad-spectrum bacterial blight resistance genes, Xa21 and Xa33 into an elite maintainer line of rice, DRR17B[J]. PLoS One, 2018, 13(10): e0201271.
[11] Song W Y, Wang G L, Chen L L, et al. A receptor ki-nase-like protein encoded by the rice disease resistance gene, Xa21[J]. Science, 1995, 270(5243): 1804-1806.
[12] Wang C L, Fan Y L, Zheng C K, et al. High-resolution ge-netic mapping of rice bacterial blight resistance gene Xa23[J]. Molecular Genetics and Genomics, 2014, 289(5): 745-753.
[13] 鄭康乐, 庄杰云, 王汉荣. 基因聚合提高了水稻对白叶枯病的抗性[J]. 遗传, 1998, 20(4): 4-6.
[14] 姬广海, 许志刚, 张世光. 水稻近等基因系对白叶枯病、条斑病抗性的比较研究[J]. 云南农业大学学报, 2000, 15(3): 187-191.
[15] Lee S W, Choi S H, Han S S, et al. Distribution of Xantho-monas oryzae pv. oryzae strains virulent to Xa21 in Ko-rea[J]. Phytopathology, 1999, 89(10): 928-933.
[16] Wang C, Zhang X, Fan Y, et al. Xa23 is an executor R pro-tein and confers broad-spectrum disease resistance in rice[J]. 2015, 22(2): 290-302.
[17] 闫 影, 王春连, 刘丕庆, 等. 野生稻抗病虫基因的挖掘和利用[J]. 作物杂志, 2011(4): 1-6.
[18] 杨 军, 王海凤, 林 枫, 等. 不同水稻品种(系)对白叶枯病的抗性分析[J]. 西南农业学报, 2017, 30(9): 1991- 1996.
[19] 兰艳荣, 王俊义, 王 弋, 等. 分子标记辅助选择改良水稻光温敏核不育系华201S的白叶枯病抗性[J]. 中国水稻科学, 2011, 25(2): 169-174.
[20] 罗彦长, 王守海, 李成荃, 等. 应用分子标记辅助选择培育抗稻白叶枯病光敏核不育系3418S[J]. 作物学报, 2003, 29(3): 402-407.
[21] 覃宝祥, 张月雄, 杨 萌, 等. 抗白叶枯病水稻不育系先抗A和天抗A的选育[J]. 杂交水稻, 2015, 30(2): 6-9.
[22] 罗彦长, 吴 爽, 王守海, 等. 聚合抗稻白叶枯病双基因三系不育系R106A的选育研究[J]. 中国农业科学, 2005, 38(11): 2157-2164.
[23] 朱明东, 谢红军, 汤国华, 等. 湖南水稻品种审定回顾与展望[J]. 湖南农业科学, 2019(12): 96-102, 107.
[24] 王 亚, 臧之光, 陈献功, 等. 6个抗稻瘟病基因在河南省主要稻种资源中的分布情况[J]. 分子植物育种, 2017, 15(12): 4976-4984.
[25] 周元飞, 戚华雄, 万丙良, 等. 运用分子标记辅助选择技术改良9311白叶枯病抗性的研究[J]. 分子植物育种, 2003, 1(3): 343-349.
[26] Xiao Y L, Li J J, Yu J H, et al. Improvement of bacterial blight and brown planthopper resistance in an elite restorer line Huazhan of Oryza[J]. Field Crops Research, 2016, 186: 47-57.
[27] 吴云雨, 王建飞, 鲍永美, 等. 65个水稻主栽品种(系)对白叶枯病菌的抗性鉴定[J]. 江苏农业学报, 2012, 28(2): 448-450.
[28] 曹立勇, 庄杰云, 占小登, 等. 抗白叶枯病杂交水稻的分子标记辅助育种[J]. 中国水稻科学, 2003, 17(2): 184-186.
[29] 杨德卫, 叶 宁, 叶新福, 等. 分子标记辅助选择Xa23基因改良早稻恢复系白叶枯病抗性研究[J]. 福建农业学报, 2015, 30(4): 351-356.
[30] 于 洁, 王耀雯, 马文清, 等. 水稻抗白叶枯病基因Xa23群体的MAS育种研究[J]. 华南农业大学学报, 2010, 31(4): 1-5.
[31] 范宏环, 王林友, 张礼霞, 等. 通过分子标记辅助选择技术选育携有水稻白叶枯病抗性基因Xa23的水稻株系[J]. 中国水稻科学, 2011, 25(3): 331-334.
[32] 李进波, 王春连, 夏明元, 等. 分子标记辅助选择Xa23基因培育杂交稻抗白叶枯病恢复系[J]. 作物学报, 2006, 32(10): 1423-1429.
责任编辑:黄东杰
猜你喜欢
世界热带农业信息(2021年7期)2021-08-11
格言·校园版(2021年30期)2021-07-06
好孩子画报(2021年3期)2021-04-02
农民致富之友(2020年8期)2020-05-11
农民致富之友(2020年1期)2020-02-11
农民致富之友(2019年30期)2019-10-21
农民致富之友(2019年23期)2019-08-16
农民致富之友(2019年14期)2019-05-24
新农村(2018年29期)2018-04-02
农产品市场周刊(2017年35期)2017-10-25
